
Molecular Characterization of ASFV and Differential Diagnosis of Erysipelothrix in ASFV-Infected Pigs in Pig Production Regions in Cameroon



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Area
2.3. Study Design
2.4. Sample Collection
2.5. DNA Extraction and ASFV Detection
2.6. ASFV Molecular Characterization, Nucleotide Sequencing and Analysis
2.7. DNA Extraction and Erysipelothrix Detection
3. Results
3.1. Diagnosis of ASFV by PCR
3.2. B646L (P72) Gene Phylogeny
3.3. E183L (P54) Gene Phylogeny
3.4. CP204L (P30) Gene Phylogeny
3.5. Intergenic Region of I73R and I329L
3.6. Analysis of the B602L Gene
3.7. Analysis of the EP402R Gene
3.8. Differential Diagnosis of Erysipelothrix
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallardo, M.C.; de la Torre Reoyo, A.; Fernández-Pinero, J.; Iglesias, I.; Muñoz, M.J.; Arias, M.L. African swine fever: A global view of the current challenge. Porc. Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef] [PubMed]
- Compans, R.W.; Cooper, M.D. Lesser-Known Large dsDNA Viruses; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 9783540686170. Available online: https://link.springer.com/chapter/10.1007/978-3-540-68618-7_2#citeas (accessed on 22 January 2022).
- Berthe, F. The global economic impact of ASF. Bull. l’OIE 2020, 2020, 1–2. [Google Scholar] [CrossRef]
- You, S.; Liu, T.; Zhang, M.; Zhao, X.; Dong, Y.; Wu, B.; Wang, Y.; Li, J.; Wei, X.; Shi, B. African swine fever outbreaks in China led to gross domestic product and economic losses. Nat. Food 2021, 2, 802–808. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Penrith, M.; Cruci, C. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Mwaengo, D.M.; Macharia, J.M.; Arias, M.; Taracha, E.A.; Soler, A.; Okoth, E.; Okoth, E.; Okoth, E.; Bishop, R.P. Enhanced discrimination of African swine fever virus isolates through nucleotide sequencing of the p54, p72, and pB602L (CVR) genes. Virus Genes 2009, 38, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Michaud, V.; Randriamparany, T.; Albina, E. Comprehensive Phylogenetic Reconstructions of African Swine Fever Virus: Proposal for a New Classification and Molecular Dating of the Virus. PLoS ONE 2013, 8, e69662. [Google Scholar] [CrossRef]
- Sanna, G.; Giudici, S.D.; Bacciu, D.; Angioi, P.P.; Giammarioli, M.; De Mia, G.M.; Oggiano, A. Improved Strategy for Molecular Characterization of African Swine Fever Viruses from Sardinia, Based on Analysis of p30, CD2V and I73R/I329L Variable Regions. Transbound. Emerg. Dis. 2017, 64, 1280–1286. [Google Scholar] [CrossRef]
- Hernaez, B.; Escribano, J.M.; Alonso, C. African swine fever virus protein p30 interaction with heterogeneous nuclear ribonucleoprotein K (hnRNP-K) during infection. FEBS Lett. 2008, 582, 3275–3280. [Google Scholar] [CrossRef]
- Brun, A.; Sussman, M.D.; Onisk, D.V.; Escribano, M.; Rock, D.L. Characterization of P30, a Highly Antigenic Membrane and Secreted Protein of African Swine Fever Virus. Virology 1992, 373, 368–373. [Google Scholar]
- Rodrõ, F.; Oviedo, Â.M.; Brun, A.; Alonso, C.; Escribano, Â.M. The African Swine Fever Virus Proteins p54 and p30 Are Involved in Two Distinct Steps of Virus Attachment and Both Contribute to the Antibody-Mediated Protective Immune Response. Virology 1998, 471, 461–471. [Google Scholar]
- Opriessnig, T.; Forde, T.; Shimoji, Y. Erysipelothrix Spp.: Past, Present, and Future Directions in Vaccine Research. Front. Vet. Sci. 2020, 7, 174. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Fujisawa, T.; Benno, Y.; Tamura, Y.; Sawada, T.; Suzuki, S.; Muramatsu, M.; Mitsuoka, T. Erysipelothrix tonsillarum sp. nov. Isolated from Tonsils of Apparently Healthy Pigs. Int. J. Syst. Bacteriol. 1987, 37, 166–168. [Google Scholar] [CrossRef]
- Brooke, C.J.; Riley, S. V Erysipelothrix rhusiopathiae: Bacteriology, epidemiology and clinical manifestations of an occupational pathogen. J. Med. Microbiol. 1999, 48, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Okada, Y.; Maruyama, T.; Ishikawa, K. Direct and Rapid Detection of Erysipelothrix rhusiopathiae DNA in Animals by PCR. J. Clin. Microbiol. 1994, 32, 1526–1531. [Google Scholar] [CrossRef]
- Yamazaki, Y. A multiplex polymerase chain reaction for discriminating Erysipelothrix rhusiopathiae from Erysipelothrix tonsillarum. J. Vet. Diagn. Investig. 2006, 387, 384–387. [Google Scholar] [CrossRef]
- Ebwanga, E.J.; Ghogomu, S.M.; Paeshuyse, J. African Swine Fever: Prevalence, Farm Characteristics, Farmer’sInsight and Attitude toward Reporting of African Swine FeverCases in the Northwest, West, Littoral and Southwest Regionsof Cameroon. Agriculture 2021, 12, 44. [Google Scholar] [CrossRef]
- MINEPIA. Plan National Opérationnel De Contrôle De la Peste Porcine Africaine (PPA) Au Cameroun. 2016. Available online: https://www.prodel.cm/wp-content/uploads/2019/04/Plan.-opérationnel-de-contrôle-de-la-peste-porcine-africaine-au-Cameroun.pdf (accessed on 22 January 2022).
- MINEPIA. Plan Strategique De Prevention Et De Lutte Contre La PESTE Des Petits Ruminants (PPR) Au Cameroun Fevrier 2015; MINEPIA: Yaoundé, Cameroon, 2015. [Google Scholar]
- Adda, R. Contribution a L’Etude De La Peste Porcine Au Cameroun. Ph.D. Thesis, Universite de Dakar, Dakar, Senegal, 1986. Available online: http://www.beep.ird.fr/collect/eismv/index/assoc/TD86-12.dir/TD86-12.pdf (accessed on 22 January 2022).
- Wade, A.; Achenbach, J.E.; Gallardo, C.; Settypalli, T.B.K.; Souley, A.; Djonwe, G.; Loitsch, A.; Dauphin, G.; Justin, J.; Ngang, E.; et al. Genetic characterization of African swine fever virus in Cameroon, 2010–2018. J. Microbiol. 2019, 57, 316–324. [Google Scholar] [CrossRef]
- Nix, R.J.; Gallardo, C.; Hutchings, G.; Blanco, E.; Dixon, L.K. Molecular epidemiology of African swine fever virus studied by analysis of four variable genome regions. Arch. Virol. 2006, 151, 2475–2494. [Google Scholar] [CrossRef]
- Silatsa, B.A.; Simo, G.; Githaka, N.; Mwaura, S.; Kamga, R.M.; Oumarou, F.; Keambou, C.; Bishop, R.P.; Djikeng, A.; Kuiate, J.R.; et al. A comprehensive survey of the prevalence and spatial distribution of ticks infesting cattle in different agro-ecological zones of Cameroon. Parasites Vectors 2019, 12, 1–14. [Google Scholar] [CrossRef]
- Malogolovkin, A.; Burmakina, G.; Tulman, E.R.; Delhon, G.; Diel, D.G.; Salnikov, N.; Kutish, G.F.; Kolbasov, D.; Rock, D.L. African swine fever virus CD2v and C-type lectin gene loci mediate serological specificity. J. Gen. Virol. 2015, 96, 866–873. [Google Scholar] [CrossRef]
- Gallardo, C.; Fernández-pinero, J.; Pelayo, V.; Gazaev, I.; Markowska-daniel, I.; Pridotkas, G.; Nieto, R.; Fernández-pacheco, P.; Bokhan, S.; Nevolko, O.; et al. Genetic Variation among African Swine Fever Genotype II Viruses, Eastern and Central Europe. Emerg. Infect. Dis. 2014, 20, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- Njayou, A. Eco-Épidémiologie de la Peste Porcine Africaine au Cameroun; Ecole Inter-Etats de Sciences et de Médecine Vétérinaire-Université Cheikh Anta Diop: Dakar, Senegal, 2016; Available online: https://scholar.google.com/scholar_lookup?title=Eco-%C3%89pid%C3%A9miologie+de+la+P (accessed on 22 January 2022).
- Ngwa, V.N.; Abouna, A.; Zoli, P.; Attili, A. Veterinary sciences epidemiology of African swine fever in piggeries in the Center, South and South-West of Cameroon. Vet. Sci. 2020, 7, 123. [Google Scholar] [CrossRef] [PubMed]
- EKue, N.; Abouna, A.; Zoli, A.P.; Attili, A.R. Epidemiology of African Swine Fever in Cameroon. Ph.D. Thesis, University of Surrey, Guildford, UK, 1989. [Google Scholar]
- Luka, P.D.; Achenbach, J.E.; Mwiine, F.N.; Lamien, C.E.; Shamaki, D.; Unger, H.; Erume, J. Genetic Characterization of Circulating African Swine Fever Viruses in Nigeria (2007–2015). Transbound. Emerg. Dis. 2016, 64, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Simulundu, E.; Sinkala, Y.; Chambaro, H.M.; Chinyemba, A.; Banda, F.; Mooya, L.E.; Ndebe, J.; Chitanga, S.; Makungu, C.; Munthali, G.; et al. Genetic characterisation of african swine fever virus from 2017 outbreaks in Zambia: Identification of p72 genotype ii variants in domestic pigs. Onderstepoort J. Vet. Res. 2018, 85, 1562. [Google Scholar] [CrossRef]
- Irusta, P.M.; Borca, M.V.; Kutish, G.F.; Lu, Z.; Caler, E.; Carrillo, C.; Rock, D.L. Amino Acid Tandem Repeats within a Late Viral Gene Define the Central Variable Region of African Swine Fever Virus. Virology 1996, 27, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Giammarioli, M.; Gallardo, C.; Oggiano, A.; Iscaro, C.; Nieto, R.; Pellegrini, C.; Dei, S.; Arias, M.; Mario, G. Genetic characterisation of African swine fever viruses from recent and historical outbreaks in Sardinia (1978–2009 ). Virus Genes 2011, 42, 377–387. [Google Scholar] [CrossRef]
- Bisimwa, P.N.; Ishara, L.K.; Wasso, S.; Bantuzeko, F.; Tonui, R.; Bwihangane, A.B. Detection and genetic characterization of African swine fever virus (ASFV) in clinically infected pigs in two districts in South Kivu province, Democratic Republic Congo. Heliyon 2021, 7, e06419. [Google Scholar] [CrossRef]
- Rowlands, R.J.; Duarte, M.M.; Boinas, F.; Hutchings, G.; Dixon, L.K. The CD2v protein enhances African swine fever virus replication in the tick vector, Ornithodoros erraticus. Virology 2009, 393, 319–328. [Google Scholar] [CrossRef]
- Goatley, L.C.; Cox, L.; Miskin, J.E.; Parkhouse, R.M.E.; Wienands, J.; Dixon, L.K.; Dixon, L. The CD2v protein of African swine fever virus interacts with the actin-binding adaptor protein SH3P7 Printed in Great Britain. J. Gen. Virol. 2004, 85, 119–130. [Google Scholar] [CrossRef]
- Malogolovkin, A.; Kolbasov, D. Genetic and antigenic diversity of African swine fever virus. Virus Res. 2019, 271, 197673. [Google Scholar] [CrossRef]
- Progress, P. Cameroon subsidises erysipelas vaccination for its pigs. Pig Health 2018, 1–4. [Google Scholar] [CrossRef]
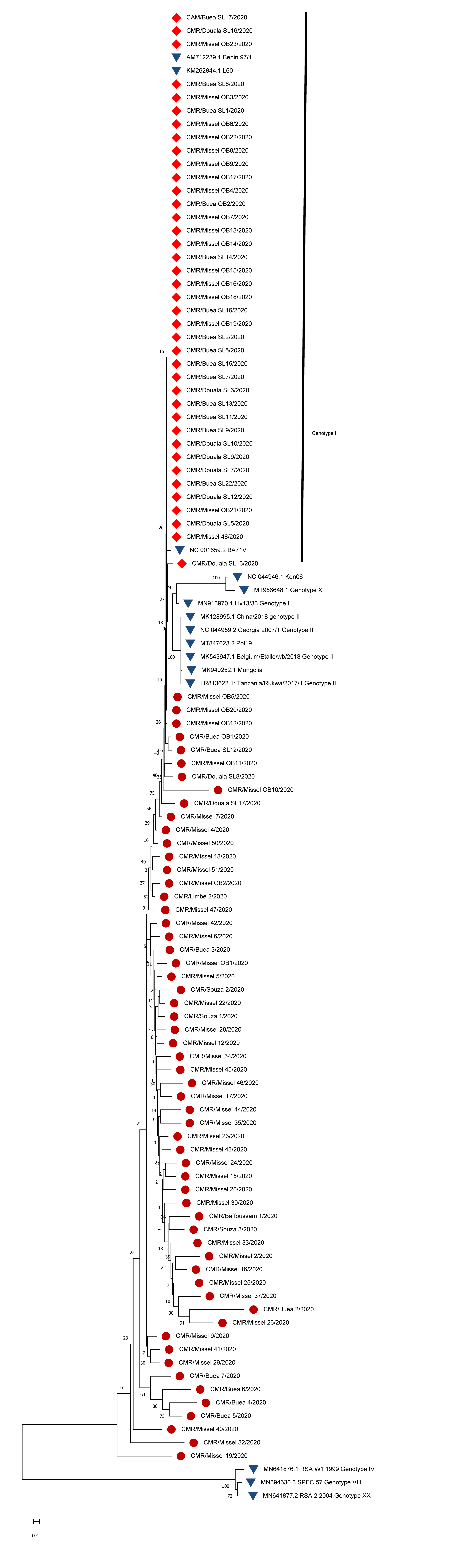
 ), isolates from previous studies in Cameroon (
), isolates from previous studies in Cameroon (  ) and other isolates representative of genotypes from GenBank (
) and other isolates representative of genotypes from GenBank (  ). The evolutionary tree was inferred by the neighbour-joining algorithm method with 1000 bootstraps, evolutionary distance computed using the p-distance method and 85% cut-off.
), isolates from previous studies in Cameroon ( ) and other isolates representative of genotypes from GenBank ( ). The evolutionary tree was inferred by the neighbour-joining algorithm method with 1000 bootstraps, evolutionary distance computed using the p-distance method and 85% cut-off.
). The evolutionary tree was inferred by the neighbour-joining algorithm method with 1000 bootstraps, evolutionary distance computed using the p-distance method and 85% cut-off.
), isolates from previous studies in Cameroon ( ) and other isolates representative of genotypes from GenBank ( ). The evolutionary tree was inferred by the neighbour-joining algorithm method with 1000 bootstraps, evolutionary distance computed using the p-distance method and 85% cut-off. ), those from previous studies in Cameroon in (
), those from previous studies in Cameroon in (  ) and other genotypes from GenBank in (
) and other genotypes from GenBank in (  ).
), those from previous studies in Cameroon in ( ) and other genotypes from GenBank in ( ).
).
), those from previous studies in Cameroon in ( ) and other genotypes from GenBank in ( ). ), isolates from this study in no cluster in (
), isolates from this study in no cluster in (  ) and other genotypes from GenBank in ( ).
), isolates from this study in no cluster in ( ) and other genotypes from GenBank in ( ).
) and other genotypes from GenBank in ( ).
), isolates from this study in no cluster in ( ) and other genotypes from GenBank in ( ).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Site | Region | Division | Farm Location or Slab | Number of Samples Collected | Total |
|---|---|---|---|---|---|
| Slaughter slabs (blood and tissue samples) | Southwest | Fako | Mile 16 Buea | 24 | 100 |
| Liottoral | Moungo | Grand Hanger market (Bonaberi) | 22 | ||
| Outbreak collection | Southwest | Fako | `Buea Misselleleh | 30 24 | |
| From pig farms (Blood only) | Littoral | Wouri | Bonaberi | 15 | 37 |
| Moungo | Souza | 22 | |||
| Southwest | Fako | Buea | 69 | 143 | |
| Limbe | 26 | ||||
| Misselleleh Area | 48 | ||||
| West | Mifi | Baffousam | 8 | 20 | |
| Koung Khi | Baham | 4 | |||
| Bandjoun | 8 | ||||
| Northwest | Mezam | Bafut | 22 | 77 | |
| Santa | 55 |
| Number of CVR Repeats | CVR Tandem Repeat Sequence (TRS) Signature | Accession Numbers of Sequences Involved |
|---|---|---|
| 19 | ABNAAAACBNABTDBNAFA | ON191096, ON191099, ON191100, ON191102, ON191103, ON191104, ON191106, ON191107, ON191110, ON191111, ON191112, ON191113, ON191115, ON191119, ON191121, ON191123, ON191124, ON191127, ON191128, ON191133, ON191134, ON191139, ON191146, ON191155, ON191157, ON191158, ON191160, ON191161, |
| 6 | AB--------------------NAFA | ON191097, ON191098, ON191101, ON191105, ON191108, ON191109, ON191114, ON191116, ON191117, ON191118, ON191120, ON191122, ON191125, ON191126, ON191129, ON191130, ON191131, ON191132, ON191135, ON191136, ON191137, ON191138, ON191140, ON191141, ON191142, ON191143, ON191144, ON191145, ON191147, ON191148, ON191149, ON191150, ON191151, ON191152, ON191153, ON191154, ON191156, ON191159, ON191162, ON191163, ON191164, ON191165, ON191166, ON191167, ON191168, ON191169, ON191170, ON191171, ON191172, ON191173, ON191174, ON191175, ON191176, ON191177, ON191178, ON191179, ON191180, ON191181, ON191182, ON191183, ON191184, ON191185, ON191186, ON191187, ON191188, ON191189, ON191190, ON191191, ON191192, ON191193, ON191194, ON191195, ON191196, ON191197, ON191198, ON191199, ON191200, ON191201, ON191202. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebwanga, E.J.; Ghogomu, S.M.; Paeshuyse, J. Molecular Characterization of ASFV and Differential Diagnosis of Erysipelothrix in ASFV-Infected Pigs in Pig Production Regions in Cameroon. Vet. Sci. 2022, 9, 440. https://doi.org/10.3390/vetsci9080440
Ebwanga EJ, Ghogomu SM, Paeshuyse J. Molecular Characterization of ASFV and Differential Diagnosis of Erysipelothrix in ASFV-Infected Pigs in Pig Production Regions in Cameroon. Veterinary Sciences. 2022; 9(8):440. https://doi.org/10.3390/vetsci9080440
Chicago/Turabian StyleEbwanga, Ebanja Joseph, Stephen Mbigha Ghogomu, and Jan Paeshuyse. 2022. "Molecular Characterization of ASFV and Differential Diagnosis of Erysipelothrix in ASFV-Infected Pigs in Pig Production Regions in Cameroon" Veterinary Sciences 9, no. 8: 440. https://doi.org/10.3390/vetsci9080440
APA StyleEbwanga, E. J., Ghogomu, S. M., & Paeshuyse, J. (2022). Molecular Characterization of ASFV and Differential Diagnosis of Erysipelothrix in ASFV-Infected Pigs in Pig Production Regions in Cameroon. Veterinary Sciences, 9(8), 440. https://doi.org/10.3390/vetsci9080440