Identification and Morphogenesis of Vestibular Atrial Septal Defects

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Methods
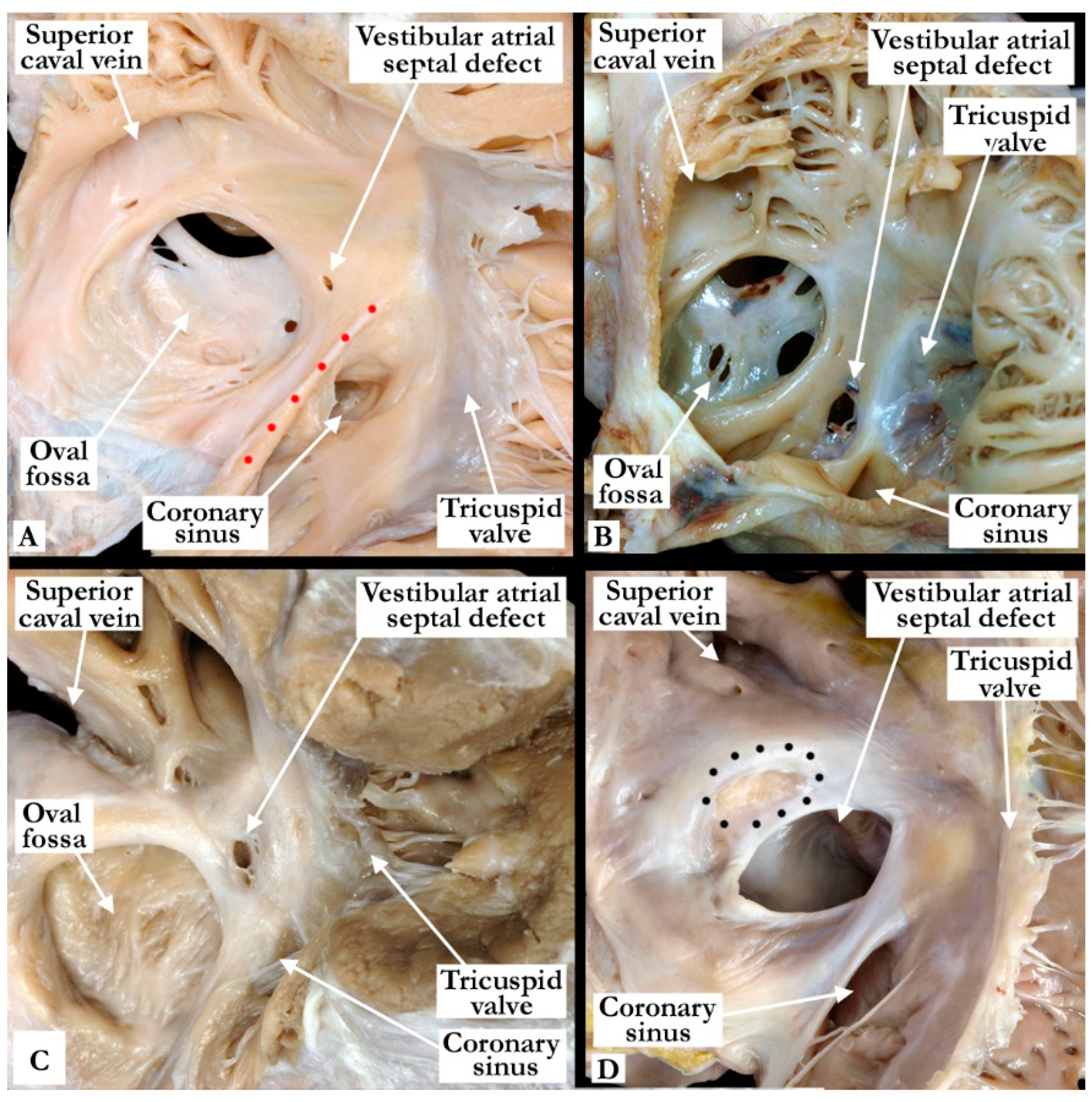
3. Results
3.1. Human Necroscopy Specimens
3.2. Developmental Considerations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jensen, B.; Spicer, D.E.; Sheppard, M.N.; Anderson, R.H. Development of the atrial septum in relation to postnatal anatomy and interatrial communications. Heart 2017, 103, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Kashima, I.; Yoshiba, S. Surgical treatment of an unusual atrial septal defect: The vestibular defect. Cardiol. Young 2004, 14, 212–214. [Google Scholar] [CrossRef]
- Sharratt, G.P.; Webb, S.; Anderson, R.H. The vestibular defect: An interatrial communication due to a deficiency in the atrial septal component derived from the vestibular spine. Cardiol. Young 2003, 13, 184–190. [Google Scholar] [CrossRef]
- Mohun, T.J.; Weninger, W.J. Imaging heart development using high-resolution episcopic microscopy. Curr. Opin. Genet. Dev. 2011, 21, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Ezon, D.; Goldberg, J.; Kyle, W. Atlas of Congenital Heart Disease Nomenclature; Ezon Educational Services: Kentucky, KY, USA, 2015. [Google Scholar]
- Webb, S.; Brown, N.A.; Anderson, R.H. Formation of the atrioventricular septal structures in the normal mouse. Circ. Res. 1998, 82, 645–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, N.A.; Ottenkamp, J.; Wenink, A.G.; Gittenberger-de Groot, A.C. Deficiency of the vestibular spine in atrioventricular septal defects in human fetuses with down syndrome. Am. J. Cardiol. 2002, 91, 180–184. [Google Scholar] [CrossRef]
- Snarr, B.S.; Wirrig, E.E.; Phelps, A.L.; Trusk, T.C.; Wessels, A. A spatiotemporal evaluation of the contribution of the dorsal mesenchymal protrusion to cardiac development. Dev. Dyn. 2007, 236, 1287–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.D.F.; Anderson, R.H. The anatomy of interatrial communications—What does the interventionist need to know? Cardiol. Young 2000, 10, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, A.; Windecker, S.; Meier, B. Evaluation and treatment of abnormalities of the interatrial septum. Catheter. Cardiovasc. Interv. 2004, 63, 94–103. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Right-Sided Superior Caval Vein | 68 (100) |
| Drainage of right-sided superior caval vein | |
| Right-sided atrium | 68 (100) |
| Left-sided superior caval vein | 4 (6) |
| Drainage of left-sided superior caval vein | |
| Coronary sinus | 4 (100) |
| Pulmonary vein drainage | |
| Left-sided atrium | 68 (100) |
| Right-sided atrium | |
| Normal | 26 (38) |
| Dilated | 42 (62) |
| Left-sided atrium | |
| Normal | 46 (68) |
| Dilated | 20 (29) |
| Hypoplastic | 2 (3) |
| Oval fossa-type atrial septal defect | 53 (78) |
| Coronary sinus defect | 3 (4) |
| Common atrioventricular junction | 10 (15) |
| Ventricular topology | |
| Right | 67 (98) |
| Left | 1 (2) |
| Atrioventricular connections | |
| Concordant | 65 (94) |
| Discordant | 1 (2) |
| Absent left-sided atrioventricular valve | 1 (2) |
| Absent right-sided atrioventricular valve | 1 (2) |
| Right-sided atrioventricular valve abnormality | 40 (59) |
| Left-sided atrioventricular valve abnormality | 19 (28) |
| Pulmonary atresia | 17 (25) |
| Aortic atresia | 1 (2) |
| Ventriculoarterial connections | |
| Concordant | 60 (89) |
| Discordant | 5 (7) |
| Double outlet right ventricle | 3 (4) |
| Right-sided ventricle morphology | |
| Tripartite | 51 (75) |
| Bipartite | 12 (18) |
| Unipartite | 5 (7) |
| Left-sided ventricle morphology | |
| Tripartite | 66 (96) |
| Bipartite | 1 (2) |
| Unipartite | 1 (2) |
| Right outflow tract obstruction | |
| None | 31 (46) |
| Infundibular | 3 (5) |
| Valvar | 6 (9) |
| Infundibular and valvar | 11 (15) |
| Valvar atresia | 17 (25) |
| Position of aorta relative to the pulmonary trunk | |
| Anterior | 1 (2) |
| Anterior, rightward | 7 (10) |
| Posterior, rightward | 58 (84) |
| Side by side, rightward | 1 (2) |
| Common arterial trunk | 1 (2) |
| Left ventricular outflow tract obstruction | |
| None | 54 (79) |
| Subvalvar | 4 (6) |
| Valvar | 7 (10) |
| Subvalvar and valvar | 2 (3) |
| Valvar atresia | 1 (2) |
| Interventricular communication | 31 (46) |
| Aortic arch sidedness | |
| Left | 66 (98) |
| Right | 1 (2) |
| Coarctation of the aorta | 4 (6) |
| Interruption of the aortic arch | 2 (3) |
| Abnormal coronary artery origins or course | 3 (6) |
| Arterial duct | |
| Absent | 27 (40) |
| Present | 33 (49) |
| Ligated | 8 (11) |
| Cardiac apex | |
| Leftward | 67 (98) |
| Pointing to middle | 1 (2) |
| Trisomy 21 | 5 (7) |
| 22 q deletion | 1 (2) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loomba, R.S.; Tretter, J.T.; Mohun, T.J.; Anderson, R.H.; Kramer, S.; Spicer, D.E. Identification and Morphogenesis of Vestibular Atrial Septal Defects. J. Cardiovasc. Dev. Dis. 2020, 7, 35. https://doi.org/10.3390/jcdd7030035
Loomba RS, Tretter JT, Mohun TJ, Anderson RH, Kramer S, Spicer DE. Identification and Morphogenesis of Vestibular Atrial Septal Defects. Journal of Cardiovascular Development and Disease. 2020; 7(3):35. https://doi.org/10.3390/jcdd7030035
Chicago/Turabian StyleLoomba, Rohit S., Justin T. Tretter, Timothy J. Mohun, Robert H. Anderson, Scott Kramer, and Diane E. Spicer. 2020. "Identification and Morphogenesis of Vestibular Atrial Septal Defects" Journal of Cardiovascular Development and Disease 7, no. 3: 35. https://doi.org/10.3390/jcdd7030035
APA StyleLoomba, R. S., Tretter, J. T., Mohun, T. J., Anderson, R. H., Kramer, S., & Spicer, D. E. (2020). Identification and Morphogenesis of Vestibular Atrial Septal Defects. Journal of Cardiovascular Development and Disease, 7(3), 35. https://doi.org/10.3390/jcdd7030035