
Extracellular Matrix-Based Biomaterials for Cardiovascular Tissue Engineering



Abstract
:1. Introduction
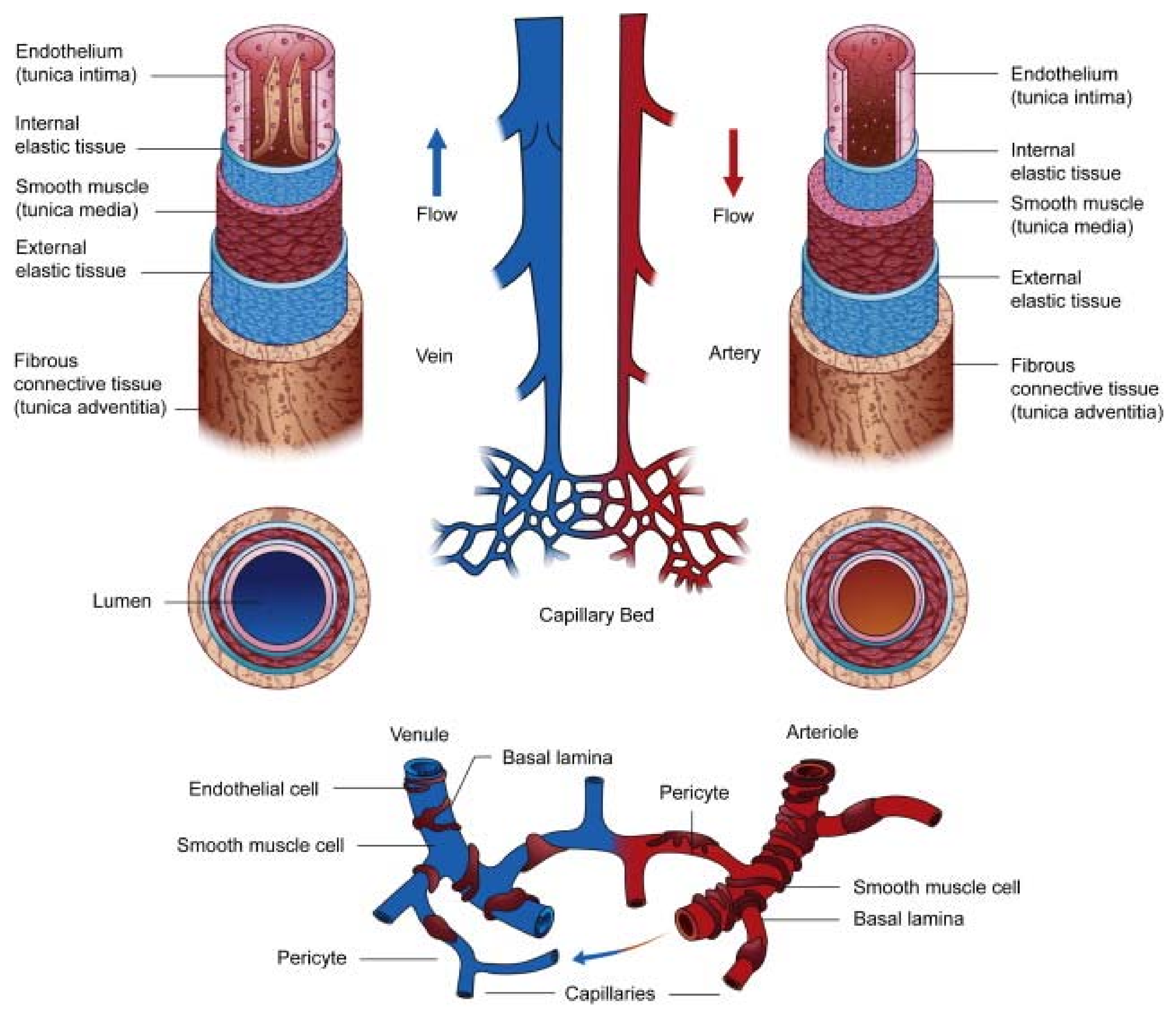
2. Overview of the Vasculature
3. ECM-Based Biomaterials
4. ECM Regulation of Vascular Function and Cell Fate
5. Considerations of Hydrogel Assembly, Biofabrication, and Spatial Patterning
6. Translational Applications of ECM-Based Biomaterials
6.1. Engineered Vascular Grafts
6.2. Cardiac Patches
6.3. Organ-on-a-Chip
7. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Reyna, M.A.; Sadr, N.; Perez Alday, E.A.; Gu, A.; Shah, A.; Robichaux, C.; Rab, A.B.; Elola, A.; Seyedi, S.; Ansari, S.; et al. Heart disease and stroke statistics—2021 update: A report from the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar]
- Yannas, I.V.; Burke, J.F.; Orgill, D.P.; Skrabut, E.M. Wound tissue can utilize a polymeric template to synthesize a functional extension of skin. Science 1982, 215, 174–176. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, J.A.; Callanan, A. Hybrid cardiovascular sourced extracellular matrix scaffolds as possible platforms for vascular tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 910–924. [Google Scholar] [CrossRef] [Green Version]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma 2014, 23, S20–S23. [Google Scholar] [CrossRef]
- Bissell, M.J.; Hall, H.G.; Parry, G. How does the extracellular matrix direct gene expression? J. Theor. Biol. 1982, 99, 31–68. [Google Scholar] [CrossRef]
- Bornstein, P. Synthesis and secretion of structural macromolecules by endothelial cells in culture. Pathobiol. Endothel. Cell 1982, 215–228. [Google Scholar]
- Schultz, G.S.; Davidson, J.M.; Kirsner, R.S.; Bornstein, P.; Herman, I.M. Dynamic reciprocity in the wound microenvironment. Wound Repair Regen. 2011, 19, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Banuth, A.M.M.; Nader, H.B.; Dreyfuss, J.L. Altered shear stress on endothelial cells leads to remodeling of extracellular matrix and induction of angiogenesis. PLoS ONE 2020, 15, e0241040. [Google Scholar] [CrossRef]
- Neve, A.; Cantatore, F.P.; Maruotti, N.; Corrado, A.; Ribatti, D. Extracellular Matrix Modulates Angiogenesis in Physiological and Pathological Conditions. BioMed Res. Int. 2014, 756078. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M. Extracellular Matrix Regulation of Stem Cell Behavior. Curr. Stem Cell Rep. 2016, 2, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 2506–2519. [Google Scholar] [CrossRef]
- Sottile, J. Regulation of angiogenesis by extracellular matrix. Biochim. Biophys. Acta (BBA) Rev. Cancer 2004, 1654, 13–22. [Google Scholar] [CrossRef]
- Schultz, G.S.; Wysocki, A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef]
- Agren, M.S.; Werthen, M. The Extracellular Matrix in Wound Healing: A Closer Look at Therapeutics for Chronic Wounds. Int. J. Low. Extrem. Wounds 2007, 6, 82–97. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, L. Human Physiology: From Cells to Systems; Cengage Learning: Belmont, CA, USA, 2015. [Google Scholar]
- Fox, S.I. Fundamentals of Human Physiology; McGraw-Hill: New York, NY, USA, 2009. [Google Scholar]
- Maton, A. Human Biology and Health; Prentice Hall: Hoboken, NJ, USA, 1997. [Google Scholar]
- Stratman, A.N.; Yu, J.A.; Mulligan, T.S.; Butler, M.G.; Sause, E.T.; Weinstein, B.M. Chapter 24—Blood Vessel Formation. Principles of Developmental Genetics, 2nd ed.; Academic Press: Oxford, UK, 2015; pp. 421–449. [Google Scholar]
- McMillan, D.B.; Harris, R.J. An Atlas of Comparative Vertebrate Histology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Tucker, W.D.; Arora, Y.; Mahajan, K. Anatomy, Blood Vessels; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Lacolley, P.; Regnault, V.; Nicoletti, A.; Li, Z.; Michel, J.-B. The vascular smooth muscle cell in arterial pathology: A cell that can take on multiple roles. Cardiovasc. Res. 2012, 95, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Cronenwett, J.L.; Johnston, K.W. Rutherford’s Vascular Surgery e-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Lüscher, T.; Predel, H.-G.; Yang, Z.; Bühler, F.; von Segesser, L.; Turina, M. Implications of pulsatile stretch on growth of saphenous vein and mammary artery smooth muscle. Lancet 1992, 340, 878–879. [Google Scholar] [CrossRef]
- Wong, A.P.; Nili, N.; Strauss, B.H. In vitro differences between venous and arterial-derived smooth muscle cells: Potential modulatory role of decorin. Cardiovasc. Res. 2005, 65, 702–710. [Google Scholar] [CrossRef]
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial function and dysfunction: Testing and clinical relevance. Circulation 2007, 115, 1285–1295. [Google Scholar] [CrossRef]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Senger, D.R.; Davis, G.E. Angiogenesis. Cold Spring Harb. Perspect. Biol. 2011, 3, a005090. [Google Scholar] [CrossRef] [Green Version]
- Hussey, G.S.; Dziki, J.L.; Badylak, S.F. Extracellular matrix-based materials for regenerative medicine. Nat. Rev. Mater. 2018, 3, 159–173. [Google Scholar] [CrossRef]
- Peppas, N.A.; Sahlin, J.J. Hydrogels as mucoadhesive and bioadhesive materials: A review. Biomaterials 1996, 17, 1553–1561. [Google Scholar] [CrossRef]
- Hu, W.; Wang, Z.; Xiao, Y.; Zhang, S.; Wang, J. Advances in crosslinking strategies of biomedical hydrogels. Biomater. Sci. 2019, 7, 843–855. [Google Scholar] [CrossRef]
- Nguyen, K.T.; West, J.L. Photopolymerizable hydrogels for tissue engineering applications. Biomaterials 2002, 23, 4307–4314. [Google Scholar] [CrossRef]
- Wu, S.; Peng, S.; Wang, C.H. Multifunctional Polymer Nanocomposites Reinforced by Aligned Carbon Nanomaterials. Polymers 2018, 10, 542. [Google Scholar] [CrossRef] [Green Version]
- Hennink, W.E.; van Nostrum, C.F. Novel crosslinking methods to design hydrogels. Adv. Drug Deliv. Rev. 2012, 64, 223–236. [Google Scholar] [CrossRef]
- Yokoyama, F.; Masada, I.; Shimamura, K.; Ikawa, T.; Monobe, K. Morphology and structure of highly elastic poly (vinyl alcohol) hydrogel prepared by repeated freezing-and-melting. Colloid Polym. Sci. 1986, 264, 595–601. [Google Scholar] [CrossRef]
- Eagland, D.; Crowther, N.J.; Butler, C.J. Complexation between polyoxyethylene and polymethacrylic acid—The importance of the molar mass of polyoxyethylene. Eur. Polym. J. 1994, 30, 767–773. [Google Scholar] [CrossRef]
- Wichterle, O.; LÍM, D. Hydrophilic Gels for Biological Use. Nature 1960, 185, 117–118. [Google Scholar] [CrossRef]
- Sperinde, J.J.; Griffith, L.G. Control and prediction of gelation kinetics in enzymatically cross-linked poly (ethylene glycol) hydrogels. Macromolecules 2000, 33, 5476–5480. [Google Scholar] [CrossRef]
- Pomeroy, J.E.; Helfer, A.; Bursac, N. Biomaterializing the promise of cardiac tissue engineering. Biotechnol. Adv. 2020, 42, 107353. [Google Scholar] [CrossRef]
- Patel, B.; Xu, Z.; Pinnock, C.B.; Kabbani, L.S.; Lam, M.T. Self-assembled Collagen-Fibrin Hydrogel Reinforces Tissue Engineered Adventitia Vessels Seeded with Human Fibroblasts. Sci. Rep. 2018, 8, 3294. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.; Kurisawa, M. Formation and stability of interpenetrating polymer network hydrogels consisting of fibrin and hyaluronic acid for tissue engineering. Acta Biomater. 2013, 9, 5143–5152. [Google Scholar] [CrossRef]
- Xu, Q.; Torres, J.E.; Hakim, M.; Babiak, P.M.; Pal, P.; Battistoni, C.M.; Nguyen, M.; Panitch, A.; Solorio, L.; Liu, J.C. Collagen-and hyaluronic acid-based hydrogels and their biomedical applications. Mater. Sci. Eng. R Rep. 2021, 146, 100641. [Google Scholar] [CrossRef]
- Kim, H.D.; Lee, E.A.; An, Y.H.; Kim, S.L.; Lee, S.S.; Yu, S.J.; Jang, H.L.; Nam, K.T.; Im, S.G.; Hwang, N.S. Chondroitin Sulfate-Based Biomineralizing Surface Hydrogels for Bone Tissue Engineering. ACS Appl. Mater. Interfaces 2017, 9, 21639–21650. [Google Scholar] [CrossRef]
- Pacak, C.A.; MacKay, A.A.; Cowan, D.B. Au–Cowan. An Improved Method for the Preparation of Type I Collagen from Skin. JoVE 2014, 83, e51011. [Google Scholar]
- Xing, H.; Lee, H.; Luo, L.; Kyriakides, T.R. Extracellular matrix-derived biomaterials in engineering cell function. Biotechnol. Adv. 2020, 42, 107421. [Google Scholar] [CrossRef]
- Tharmalingam, T.; Sunley, K.; Spearman, M.; Butler, M. Enhanced Production of Human Recombinant Proteins from CHO cells Grown to High Densities in Macroporous Microcarriers. Mol. Biotechnol. 2011, 49, 263–276. [Google Scholar] [CrossRef]
- Nagaoka, M.; Jiang, H.L.; Hoshiba, T.; Akaike, T.; Cho, C.S. Application of Recombinant Fusion Proteins for Tissue Engineering. Ann. Biomed. Eng. 2010, 38, 683–693. [Google Scholar] [CrossRef]
- Kutty, J.K.; Cho, E.; Lee, J.S.; Vyavahare, N.R.; Webb, K. The effect of hyaluronic acid incorporation on fibroblast spreading and proliferation within PEG-diacrylate based semi-interpenetrating networks. Biomaterials 2007, 28, 4928–4938. [Google Scholar] [CrossRef]
- Nakayama, K.H.; Surya, V.N.; Gole, M.; Walker, T.W.; Yang, W.; Lai, E.S.; Ostrowski, M.A.; Fuller, G.G.; Dunn, A.R.; Huang, N.F. Nanoscale patterning of extracellular matrix alters endothelial function under shear stress. Nano Lett. 2016, 16, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.H.; Joshi, P.A.; Lai, E.S.; Gujar, P.; Joubert, L.-M.; Chen, B.; Huang, N.F. Bilayered vascular graft derived from human induced pluripotent stem cells with biomimetic structure and function. Regen. Med. 2015, 10, 745–755. [Google Scholar] [CrossRef]
- Ozguldez, H.O.; Cha, J.; Hong, Y.; Koh, I.; Kim, P. Nanoengineered, cell-derived extracellular matrix influences ECM-related gene expression of mesenchymal stem cells. Biomater. Res. 2018, 22, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.; Cho, S.; Discher, D.E. Stem cell differentiation is regulated by extracellular matrix mechanics. Physiology 2018, 33, 16–25. [Google Scholar] [CrossRef]
- Pati, F.; Jang, J.; Ha, D.H.; Kim, S.W.; Rhie, J.W.; Shim, J.H.; Kim, D.H.; Cho, D.W. Printing three-dimensional tissue analogues with decellularized extracellular matrix bioink. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Coller, J.; Natu, V.; Hastie, T.J.; Huang, N.F. Combinatorial extracellular matrix microenvironments promote survival and phenotype of human induced pluripotent stem cell-derived endothelial cells in hypoxia. Acta Biomater. 2016, 44, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Okogbaa, J.; Lee, J.; Jha, A.; Zaitseva, T.S.; Paukshto, M.V.; Sun, J.S.; Punjya, N.; Fuller, G.G.; Cooke, J.P. The modulation of endothelial cell morphology, function, and survival using anisotropic nanofibrillar collagen scaffolds. Biomaterials 2013, 34, 4038–4047. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.Y.; Chen, Y.-C.; Kao, Y.-H.; Wu, T.-J.; Chen, S.-A.; Chen, Y.-J. Extracellular matrix of collagen modulates intracellular calcium handling and electrophysiological characteristics of HL-1 cardiomyocytes with activation of angiotensin II type 1 receptor. J. Card. Fail. 2011, 17, 82–90. [Google Scholar] [CrossRef]
- Schönberger, T.; Ziegler, M.; Borst, O.; Konrad, I.; Nieswandt, B.; Massberg, S.; Ochmann, C.; Jürgens, T.; Seizer, P.; Langer, H.; et al. The dimeric platelet collagen receptor GPVI-Fc reduces platelet adhesion to activated endothelium and preserves myocardial function after transient ischemia in mice. Am. J. Physiol. Physiol. 2012, 303, C757–C766. [Google Scholar] [CrossRef]
- Zhang, W.; Kong, C.W.; Tong, M.H.; Chooi, W.H.; Huang, N.; Li, R.A.; Chan, B.P. Maturation of human embryonic stem cell-derived cardiomyocytes (hESC-CMs) in 3D collagen matrix: Effects of niche cell supplementation and mechanical stimulation. Acta Biomater. 2017, 49, 204–217. [Google Scholar] [CrossRef]
- Bassat, E.; Mutlak, Y.E.; Genzelinakh, A.; Shadrin, I.; Umansky, K.B.; Yifa, O.; Kain, D.; Rajchman, D.; Leach, J.; Bassat, D.R.; et al. The extracellular matrix protein agrin promotes heart regeneration in mice. Nature 2017, 547, 179–184. [Google Scholar] [CrossRef]
- Schenke-Layland, K.; Rhodes, K.E.; Angelis, E.; Butylkova, Y.; Heydarkhan-Hagvall, S.; Gekas, C.; Zhang, R.; Goldhaber, J.I.; Mikkola, H.K.; Plath, K.; et al. Reprogrammed Mouse Fibroblasts Differentiate into Cells of the Cardiovascular and Hematopoietic Lineages. Stem Cells 2008, 26, 1537–1546. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.J.; Kant, R.J.; Minor, A.J.; Coulombe, K.L. Optimizing Blended Collagen-Fibrin Hydrogels for Cardiac Tissue Engineering with Human iPSC-derived Cardiomyocytes. ACS Biomater. Sci. Eng. 2019, 5, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Fathi, A.; Mithieux, S.M.; Wei, H.; Chrzanowski, W.; Valtchev, P.; Weiss, A.S.; Dehghani, F. Elastin based cell-laden injectable hy-drogels with tunable gelation, mechanical and biodegradation properties. Biomaterials 2014, 35, 5425–5435. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ali, M.S.; Lacerda, C.M.R. A Three-Dimensional Collagen-Elastin Scaffold for Heart Valve Tissue Engineering. Bioengineering 2018, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Crapo, P.M.; Gilbert, T.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [Green Version]
- Keane, T.; Swinehart, I.T.; Badylak, S.F. Methods of tissue decellularization used for preparation of biologic scaffolds and in vivo relevance. Methods 2015, 84, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.S.; Kim, H.; Gao, G.; Jang, J.; Cho, D.-W. Decellularized extracellular matrix: A step towards the next generation source for bioink manufacturing. Biofabrication 2017, 9, 034104. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, T.; Sellaro, T.L.; Badylak, S.F. Decellularization of tissues and organs. Biomaterials 2006, 27, 3675–3683. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Jang, J. 3D bioprinting and decellularized ECM-based biomaterials for in vitro CV tissue engineering. J. 3D Print. Med. 2018, 2, 69–87. [Google Scholar] [CrossRef]
- McFetridge, P.S.; Daniel, J.W.; Bodamyali, T.; Horrocks, M.; Chaudhuri, J.B. Preparation of porcine carotid arteries for vascular tissue engineering applications. J. Biomed. Mater. Res. 2004, 70A, 224–234. [Google Scholar] [CrossRef]
- Sellaro, T.L.; Ravindra, A.K.; Stolz, D.B.; Badylak, S.F. Maintenance of Hepatic Sinusoidal Endothelial Cell PhenotypeIn VitroUsing Organ-Specific Extracellular Matrix Scaffolds. Tissue Eng. 2007, 13, 2301–2310. [Google Scholar] [CrossRef]
- Bejleri, D.; Davis, M.E. Decellularized Extracellular Matrix Materials for Cardiac Repair and Regeneration. Adv. Healthc. Mater. 2019, 8, e1801217. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Extracellular matrix as an inductive scaffold for functional tissue reconstruction. Transl. Res. 2013, 163, 268–285. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Hwang, D.G.; Jang, J. 3D Pancreatic Tissue Modeling in vitro: Advances and Prospects. BioChip J. 2020, 14, 84–99. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.-G.; Lee, H.; Cho, D.-W. 3D Printing of Organs-On-Chips. Bioengineering 2017, 4, 10. [Google Scholar] [CrossRef]
- Taylor, D.A.; Sampaio, L.C.; Ferdous, Z.; Gobin, A.S.; Taite, L.J. Decellularized matrices in regenerative medicine. Acta Biomater. 2018, 74, 74–89. [Google Scholar] [CrossRef]
- Serpooshan, V.; Zhao, M.; Metzler, S.A.; Wei, K.; Shah, P.B.; Wang, A.; Mahmoudi, M.; Malkovskiy, A.V.; Rajadas, J.; Butte, M.; et al. The effect of bioengineered acellular collagen patch on cardiac remodeling and ventricular function post myocardial infarction. Biomaterials 2013, 34, 9048–9055. [Google Scholar] [CrossRef] [Green Version]
- Basara, G.; Ozcebe, S.; Ellis, B.; Zorlutuna, P. Tunable Human Myocardium Derived Decellularized Extracellular Matrix for 3D Bioprinting and Cardiac Tissue Engineering. Gels 2021, 7, 70. [Google Scholar] [CrossRef]
- Simsa, R.; Padma, A.M.; Heher, P.; Hellström, M.; Teuschl, A.; Jenndahl, L.; Bergh, N.; Fogelstrand, P. Systematic in vitro comparison of decellu-larization protocols for blood vessels. PLoS ONE 2018, 13, e0209269. [Google Scholar] [CrossRef]
- Kurokawa, S.; Hashimoto, Y.; Funamoto, S.; Murata, K.; Yamashita, A.; Yamazaki, K.; Ikeda, T.; Minatoya, K.; Kishida, A.; Masumoto, H. In vivo recellularization of xenogeneic vascular grafts decellularized with high hydrostatic pressure method in a porcine carotid arterial interpose model. PLoS ONE 2021, 16, e0254160. [Google Scholar] [CrossRef]
- Karantalis, V.; Suncion-Loescher, V.Y.; Bagno, L.; Golpanian, S.; Wolf, A.; Sanina, C.; Premer, C.; Kanelidis, A.J.; McCall, F.; Wang, B.; et al. Synergistic Effects of Combined Cell Therapy for Chronic Ischemic Cardiomyopathy. J. Am. Coll. Cardiol. 2015, 66, 1990–1999. [Google Scholar] [CrossRef]
- Spitzer, G.; Adkins, D.; Mathews, M.; Velasquez, W.; Bowers, C.; Dunphy, F.; Kronmueller, N.; Niemeyer, R.; McIntyre, W.; Petruska, P. Randomized comparison of G-CSF + GM-CSF vs G-CSF alone for mobilization of peripheral blood stem cells: Effects on hematopoietic recovery after high-dose chemotherapy. Bone Marrow Transplant. 1997, 20, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Ballotta, V.; Smits, A.I.; Driessen-Mol, A.; Bouten, C.V.; Baaijens, F.P. Synergistic protein secretion by mesenchymal stromal cells seeded in 3D scaffolds and circulating leukocytes in physiological flow. Biomaterials 2014, 35, 9100–9113. [Google Scholar] [CrossRef]
- Iwase, T.; Nagaya, N.; Fujii, T.; Itoh, T.; Murakami, S.; Matsumoto, T.; Kangawa, K.; Kitamura, S. Comparison of angiogenic potency between mesenchymal stem cells and mononuclear cells in a rat model of hindlimb ischemia. Cardiovasc. Res. 2005, 66, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Van Velthoven, C.T.; Kavelaars, A.; Heijnen, C.J. Mesenchymal stem cells as a treatment for neonatal ischemic brain damage. Pediatr. Res. 2012, 71, 474–481. [Google Scholar] [CrossRef]
- Carvalho, M.S.; Silva, J.C.; Hoff, C.M.; Cabral, J.M.S.; Linhardt, R.J.; Da Silva, C.L.; Vashishth, D. Loss and rescue of osteocalcin and osteopontin modulate osteogenic and angiogenic features of mesenchymal stem/stromal cells. J. Cell. Physiol. 2020, 235, 7496–7515. [Google Scholar] [CrossRef]
- Jehn, P.; Winterboer, J.; Kampmann, A.; Zimmerer, R.; Spalthoff, S.; Dittmann, J.; Gellrich, N.-C.; Tavassol, F. Angiogenic effects of mesenchymal stem cells in combination with different scaffold materials. Microvasc. Res. 2019, 127, 103925. [Google Scholar] [CrossRef]
- Efraim, Y.; Sarig, H.; Anavy, N.C.; Sarig, U.; de Berardinis, E.; Chaw, S.-Y.; Krishnamoorthi, M.; Kalifa, J.; Bogireddi, H.; Duc, T.V.; et al. Biohybrid cardiac ECM-based hydrogels improve long term cardiac function post myocardial infarction. Acta Biomater. 2016, 50, 220–233. [Google Scholar] [CrossRef]
- Jung, J.P.; Hu, D.; Domian, I.J.; Ogle, B.M. An integrated statistical model for enhanced murine cardiomyocyte differentiation via optimized engagement of 3D extracellular matrices. Sci. Rep. 2015, 5, 18705. [Google Scholar] [CrossRef] [Green Version]
- Hirata, M.; Yamaoka, T. Effect of stem cell niche elasticity/ECM protein on the self-beating cardiomyocyte differentiation of induced pluripotent stem (iPS) cells at different stages. Acta Biomater. 2018, 65, 44–52. [Google Scholar] [CrossRef]
- Tao, Z.-W.; Mohamed, M.; Hogan, M.; Gutierrez, L.; Birla, R.K. Optimizing a spontaneously contracting heart tissue patch with rat neonatal cardiac cells on fibrin gel. J. Tissue Eng. Regen. Med. 2014, 11, 153–163. [Google Scholar] [CrossRef]
- Edalat, S.G.; Jang, Y.; Kim, J.; Park, Y. Collagen Type I Containing Hybrid Hydrogel Enhances Cardiomyocyte Maturation in a 3D Cardiac Model. Polymers 2019, 11, 687. [Google Scholar] [CrossRef] [Green Version]
- Le, L.; Mohindra, P.; Fang, Q.; Sievers, R.E.; Mkrtschjan, M.A.; Solis-Ocampo, C.; Safranek, C.W.; Russell, B.; Lee, R.J.; Desai, T.A. Injectable hyaluronic acid based microrods provide local micromechanical and biochemical cues to attenuate cardiac fibrosis after myocardial infarction. Biomaterials 2018, 169, 11–21. [Google Scholar] [CrossRef]
- French, K.M.; Maxwell, J.T.; Bhutani, S.; Ghosh-Choudhary, S.; Fierro, M.J.; Johnson, T.D.; Christman, K.L.; Taylor, W.R.; Davis, M.E. Fibronectin and Cyclic Strain Improve Cardiac Progenitor Cell Regenerative Potential In Vitro. Stem Cells Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, J.M.; Czajka, C.A.; Patel, U.B.; Freytes, D.O.; Tobita, K.; Gilbert, T.; Badylak, S.F. Preparation of Cardiac Extracellular Matrix from an Intact Porcine Heart. Tissue Eng. Part C Methods 2010, 16, 525–532. [Google Scholar] [CrossRef]
- Weymann, A.; Loganathan, S.; Takahashi, H.; Schies, C.; Claus, B.; Hirschberg, K.; Soós, P.; Korkmaz, S.; Schmack, B.; Karck, M.; et al. Development and Evaluation of a Perfusion Decellularization Porcine Heart Model-Generation of 3-Dimensional Myocardial Neoscaffolds. Circ. J. 2011, 75, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Akhyari, P.; Aubin, H.; Gwanmesia, P.; Barth, M.; Hoffmann, S.; Huelsmann, J.; Preuss, K.; Lichtenberg, A. The quest for an optimized protocol for whole-heart decellularization: A comparison of three popular and a novel decellularization technique and their diverse ef-fects on crucial extracellular matrix qualities. Tissue Eng. Part C Methods 2011, 17, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Grauss, R.W.; Hazekamp, M.G.; Oppenhuizen, F.; Van Munsterena, C.J.; Groot, A.C.G.-D.; DeRuiter, M.C. Histological evaluation of decellularised porcine aortic valves: Matrix changes due to different decellularisation methods. Eur. J. Cardio-Thoracic Surg. 2005, 27, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Muto, A.; Chan, S.A.; Breuer, C.; Niklason, L.E. Development of Decellularized Human Umbilical Arteries as Small-Diameter Vascular Grafts. Tissue Eng. Part A 2009, 15, 2665–2676. [Google Scholar] [CrossRef] [PubMed]
- Seifu, D.G.; Purnama, A.; Mequanint, K.; Mantovani, D. Small-diameter vascular tissue engineering. Nat. Rev. Cardiol. 2013, 10, 410–421. [Google Scholar] [CrossRef]
- Shin, Y.J.; Shafranek, R.T.; Tsui, J.H.; Walcott, J.; Nelson, A.; Kim, D.H. 3D bioprinting of mechanically tuned bioinks derived from cardiac decellularized extracellular matrix. Acta Biomater. 2021, 119, 75–88. [Google Scholar] [CrossRef]
- Tsui, J.H.; Leonard, A.; Camp, N.D.; Long, J.T.; Nawas, Z.Y.; Chavanachat, R.; Smith, A.S.; Choi, J.S.; Dong, Z.; Ahn, E.H.; et al. Tunable electroconductive decellularized extracellular matrix hydrogels for engineering human cardiac microphysiological systems. Biomaterials 2021, 272, 120764. [Google Scholar] [CrossRef]
- Das, S.; Kim, S.-W.; Choi, Y.-J.; Lee, S.; Lee, S.-H.; Kong, J.-S.; Park, H.-J.; Cho, D.-W.; Jang, J. Decellularized extracellular matrix bioinks and the external stimuli to enhance cardiac tissue development in vitro. Acta Biomater. 2019, 95, 188–200. [Google Scholar] [CrossRef]
- Akbay, E.; Onur, M.A. Investigation of survival and migration potential of differentiated cardiomyocytes transplanted with decellularized heart scaffold. J. Biomed. Mater. Res. Part A 2018, 107, 561–570. [Google Scholar] [CrossRef]
- Cheng, H.-W.; Tsui, Y.-K.; Cheung, K.M.; Chan, D.; Chan, B.P. Decellularization of Chondrocyte-Encapsulated Collagen Microspheres: A Three-Dimensional Model to Study the Effects of Acellular Matrix on Stem Cell Fate. Tissue Eng. Part C Methods 2009, 15, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.-H.; Choi, B.H.; Park, S.R.; Kim, B.J.; Min, B.-H. The chondrogenic differentiation of mesenchymal stem cells on an extracellular matrix scaffold derived from porcine chondrocytes. Biomaterials 2010, 31, 5355–5365. [Google Scholar] [CrossRef]
- Liao, J.; Guo, X.; Grande-Allen, K.J.; Kasper, F.; Mikos, A.G. Bioactive polymer/extracellular matrix scaffolds fabricated with a flow perfusion bioreactor for cartilage tissue engineering. Biomaterials 2010, 31, 8911–8920. [Google Scholar] [CrossRef] [Green Version]
- Wolchok, J.C.; Tresco, P.A. The isolation of cell derived extracellular matrix constructs using sacrificial open-cell foams. Biomaterials 2010, 31, 9595–9603. [Google Scholar] [CrossRef]
- Lu, H.; Hoshiba, T.; Kawazoe, N.; Chen, G. Autologous extracellular matrix scaffolds for tissue engineering. Biomaterials 2011, 32, 2489–2499. [Google Scholar] [CrossRef]
- Alberts, B. Cell Biology: The Endless Frontier. Mol. Biol. Cell 2010, 21, 3785. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Shi, G.-P. Vascular wall extracellular matrix proteins and vascular diseases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 2106–2119. [Google Scholar] [CrossRef] [Green Version]
- Yap, L.; Tay, H.G.; Nguyen, M.T.; Tjin, M.S.; Tryggvason, K. Laminins in Cellular Differentiation. Trends Cell Biol. 2019, 29, 987–1000. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.; Okina, E.; Chai, X.; Tan, K.H.; Hovatta, O.; Ghosh, S.; Tryggvason, K. Differentiation of Human Embryonic Stem Cells to Endothelial Progenitor Cells on Laminins in Defined and Xeno-free Systems. Stem Cell Rep. 2016, 7, 802–816. [Google Scholar] [CrossRef] [Green Version]
- Stamati, K.; Priestley, J.V.; Mudera, V.; Cheema, U. Laminin promotes vascular network formation in 3D in vitro collagen scaffolds by regulating VEGF uptake. Exp. Cell Res. 2014, 327, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.E.; Senger, D.R. Endothelial Extracellular Matrix. Circ. Res. 2005, 97, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- Watt, F.M.; Huck, W.T.S. Role of the extracellular matrix in regulating stem cell fate. Nat. Rev. Mol. Cell Biol. 2013, 14, 467–473. [Google Scholar] [CrossRef]
- Hou, L.; Kim, J.J.; Wanjare, M.; Patlolla, B.; Coller, J.; Natu, V.; Hastie, T.J.; Huang, N.F. Combinatorial Extracellular Matrix Microenvironments for Probing Endothelial Differentiation of Human Pluripotent Stem Cells. Sci. Rep. 2017, 7, 6551. [Google Scholar] [CrossRef] [Green Version]
- Stratman, A.N.; Saunders, W.B.; Sacharidou, A.; Koh, W.; Fisher, K.E.; Zawieja, D.C.; Davis, M.J.; Davis, G.E. Endothelial cell lumen and vascular guidance tunnel formation requires MT1-MMP–dependent proteolysis in 3-dimensional collagen matrices. Blood 2009, 114, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Monchaux, E.; Vermette, P. Effects of surface properties and bioactivation of biomaterials on endothelial cells. Front. Biosci. 2009, 2, 239–255. [Google Scholar]
- Daum, R.; Visser, D.; Wild, C.; Kutuzova, L.; Schneider, M.; Lorenz, G.; Weiss, M.; Hinderer, S.; Stock, U.A.; Seifert, M.; et al. Fibronectin Adsorption on Electrospun Synthetic Vascular Grafts Attracts Endothelial Progenitor Cells and Promotes Endothelialization in Dynamic In Vitro Culture. Cells 2020, 9, 778. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-S.; Lee, F.; Lim, J.; Du, C.; Wan, A.C.; Lee, S.S.; Kurisawa, M. Enzymatic conjugation of a bioactive peptide into an injectable hyaluronic acid–tyramine hydrogel system to promote the formation of functional vasculature. Acta Biomater. 2014, 10, 2539–2550. [Google Scholar] [CrossRef]
- Ning, L.-J.; Zhang, Y.-J.; Zhang, Y.; Qing, Q.; Jiang, Y.-L.; Yang, J.-L.; Luo, J.-C.; Qin, T.-W. The utilization of decellularized tendon slices to provide an inductive microenvironment for the proliferation and tenogenic differentiation of stem cells. Biomaterials 2015, 52, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Jin, L.; Wu, B.; He, H.; Yang, F.; Xu, L.; Lei, Y.; Wang, Y. A method for simultaneously crosslinking and functionalizing extracellular matrix-based biomaterials as bioprosthetic heart valves with enhanced endothelialization and reduced inflammation. Acta Biomater. 2020, 119, 89–100. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, M.; Yang, Z.; Luo, R.; Lu, X.; Huang, N.; Huang, P.; Leng, Y. Cooperative control of blood compatibility and re-endothelialization by immobilized heparin and substrate topography. Acta Biomater. 2015, 15, 150–163. [Google Scholar] [CrossRef]
- Filová, E.; Brynda, E.; Riedel, T.; Chlupáč, J.; Vandrovcová, M.; Švindrych, Z.; Lisá, V.; Houska, M.; Pirk, J.; Bačáková, L. Improved adhesion and differentiation of endothelial cells on surface-attached fibrin structures containing extracellular matrix proteins. J. Biomed. Mater. Res. Part A 2013, 102, 698–712. [Google Scholar] [CrossRef]
- Caiado, F.; Carvalho, T.; Silva, F.; Castro, C.; Clode, N.; Dye, J.F.; Dias, S. The role of fibrin E on the modulation of endothelial progenitors adhesion, differentiation and angiogenic growth factor production and the promotion of wound healing. Biomaterials 2011, 32, 7096–7105. [Google Scholar] [CrossRef]
- Kang, D.; Kim, J.H.; Jeong, Y.H.; Kwak, J.Y.; Yoon, S.; Jin, S. Endothelial monolayers on collagen-coated nanofibrous membranes: Cell–cell and cell–ECM interactions. Biofabrication 2016, 8, 025008. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lin, J.; Roy, K. Effect of 3D scaffold and dynamic culture condition on the global gene expression profile of mouse embryonic stem cells. Biomaterials 2006, 27, 5978–5989. [Google Scholar] [CrossRef] [PubMed]
- VanWinkle, W.B.; Snuggs, M.B.; Buja, L.M. Cardiogel: A biosynthetic extracellular matrix for cardiomyocyte culture In Vitro. Cell. Dev. Biol.-Anim. 1996, 32, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Uchida, N.; Sivaraman, S.; Amoroso, N.J.; Wagner, W.R.; Nishiguchi, A.; Matsusaki, M.; Akashi, M.; Nagatomi, J. Nanometer-sized extracellular matrix coating on polymer-based scaffold for tissue engineering applications. J. Biomed. Mater. Res. Part A 2015, 104, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Maia, F.R.; Fonseca, K.B.; Rodrigues, G.; Granja, P.L.; Barrias, C.C. Matrix-driven formation of mesenchymal stem cell–extracellular matrix microtissues on soft alginate hydrogels. Acta Biomater. 2014, 10, 3197–3208. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Foster, A.A.; Dewi, R.E.; Cai, L.; Hou, L.; Strassberg, Z.; Alcazar, C.A.; Heilshorn, S.C.; Huang, N.F. Protein-engineered hydrogels enhance the survival of induced pluripotent stem cell-derived endothelial cells for treatment of peripheral arterial disease. Biomater. Sci. 2018, 6, 614–622. [Google Scholar] [CrossRef]
- Sun, T.; Shi, Q.; Huang, Q.; Wang, H.; Xiong, X.; Hu, C.; Fukuda, T. Magnetic alginate microfibers as scaffolding elements for the fabrication of microvascular-like structures. Acta Biomater. 2017, 66, 272–281. [Google Scholar] [CrossRef]
- Benning, L.; Gutzweiler, L.; Tröndle, K.; Riba, J.; Zengerle, R.; Koltay, P.; Zimmermann, S.; Stark, G.B.; Finkenzeller, G. Cytocompatibility testing of hydrogels toward bioprinting of mesenchymal stem cells. J. Biomed. Mater. Res. Part A 2017, 105, 3231–3241. [Google Scholar] [CrossRef]
- Suri, S.; Schmidt, C.E. Photopatterned collagen–hyaluronic acid interpenetrating polymer network hydrogels. Acta Biomater. 2009, 5, 2385–2397. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, Z.; O’Neill, J.; Wan, L.Q.; Freytes, D.O.; Vunjak-Novakovic, G. Hybrid gel composed of native heart matrix and collagen induces cardiac differentiation of human embryonic stem cells without supplemental growth factors. J. Cardiovasc. Transl. Res. 2011, 4, 605. [Google Scholar] [CrossRef] [Green Version]
- Shakouri-Motlagh, A.; O’Connor, A.J.; Brennecke, S.P.; Kalionis, B.; Heath, D.E. Native and solubilized decellularized extracellular matrix: A critical assessment of their potential for improving the expansion of mesenchymal stem cells. Acta Biomater. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Candiello, J.; Grandhi, T.S.P.; Goh, S.K.; Vaidya, V.; Lemmon-Kishi, M.; Eliato, K.R.; Ros, R.; Kumta, P.N.; Rege, K.; Banerjee, I. 3D heterogeneous islet organoid generation from human embryonic stem cells using a novel engineered hydrogel platform. Biomaterials 2018, 177, 27–39. [Google Scholar] [CrossRef]
- Giobbe, G.G.; Crowley, C.; Luni, C.; Campinoti, S.; Khedr, M.; Kretzschmar, K.; De Santis, M.M.; Zambaiti, E.; Michielin, F.; Meran, L.; et al. Extracellular matrix hydrogel derived from decellularized tissues enables endodermal organoid culture. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, K.; Pan, Z.; Guan, E.; Ge, S.; Liu, Y.; Nakamura, T.; Ren, X.-D.; Rafailovich, M.; Clark, R.A. Cell adaptation to a physiologically relevant ECM mimic with different viscoelastic properties. Biomaterials 2007, 28, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Dziki, J.L.; Badylak, S.F. Extracellular Matrix for Myocardial Repair. Adv. Exp. Med. Biol. 2018, 151–171. [Google Scholar] [CrossRef]
- Drury, J.L.; Mooney, D. Hydrogels for tissue engineering: Scaffold design variables and applications. Biomaterials 2003, 24, 4337–4351. [Google Scholar] [CrossRef]
- Francis, R.; Kumar, D.S. Biomedical Applications of Polymeric Materials and Composites; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Brigham, M.D.; Bick, A.; Lo, E.; Bendali, A.; Burdick, J.A.; Khademhosseini, A. Mechanically Robust and Bioadhesive Collagen and Photocrosslinkable Hyaluronic Acid Semi-Interpenetrating Networks. Tissue Eng. Part A 2009, 5, 1645–1653. [Google Scholar] [CrossRef]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef]
- Malda, J.; Visser, J.; Melchels, F.P.; Jüngst, T.; Hennink, W.E.; Dhert, W.; Groll, J.; Hutmacher, D.W. 25th Anniversary Article: Engineering Hydrogels for Biofabrication. Adv. Mater. 2013, 25, 5011–5028. [Google Scholar] [CrossRef]
- Murphy, S.V.; Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef]
- Colly, A.; Marquette, C.; Courtial, E.-J. Poloxamer/Poly(ethylene glycol) Self-Healing Hydrogel for High-Precision Freeform Reversible Embedding of Suspended Hydrogel. Langmuir 2021, 37, 4154–4162. [Google Scholar] [CrossRef]
- Xavier, J.R.; Thakur, T.; Desai, P.; Jaiswal, M.K.; Sears, N.; Cosgriff-Hernandez, E.; Kaunas, R.; Gaharwar, A. Bioactive Nanoengineered Hydrogels for Bone Tissue Engineering: A Growth-Factor-Free Approach. ACS Nano 2015, 9, 3109–3118. [Google Scholar] [CrossRef]
- Gaharwar, A.; Rivera, C.P.; Wu, C.-J.; Schmidt, G. Transparent, elastomeric and tough hydrogels from poly(ethylene glycol) and silicate nanoparticles. Acta Biomater. 2011, 7, 4139–4148. [Google Scholar] [CrossRef]
- Peak, C.W.; Carrow, J.K.; Thakur, A.; Singh, A.; Gaharwar, A.K. Elastomeric Cell-Laden Nanocomposite Microfibers for Engineering Complex Tissues. Cell. Mol. Bioeng. 2015, 8, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.W.; Lee, S.J.; Ko, I.K.; Kengla, C.; Yoo, J.J.; Atala, A. A 3D bioprinting system to produce human-scale tissue constructs with structural integrity. Nat. Biotechnol. 2016, 34, 312–319. [Google Scholar] [CrossRef]
- Gasperini, L.; Mano, J.F.; Reis, R.L. Natural polymers for the microencapsulation of cells. J. R. Soc. Interface 2014, 11, 20140817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimene, D.; Lennox, K.K.; Kaunas, R.R.; Gaharwar, A.K. Advanced Bioinks for 3D Printing: A Materials Science Perspective. Ann. Biomed. Eng. 2016, 44, 2090–2102. [Google Scholar] [CrossRef]
- Grover, G.N.; Rao, N.; Christman, K.L. Myocardial matrix–polyethylene glycol hybrid hydrogels for tissue engineering. Nanotechnology 2013, 25, 014011. [Google Scholar] [CrossRef]
- Mintz, B.R.; Cooper, J.A., Jr. Hybrid hyaluronic acid hydrogel/poly (ε-caprolactone) scaffold provides mechanically favorable platform for cartilage tissue engineering studies. J. Biomed. Mater. Res. Part A 2014, 102, 2918–2926. [Google Scholar] [CrossRef] [PubMed]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, S.; Kwon, M.; Burdick, J. Recent advances in hydrogels for cartilage tissue engineering. Eur. Cells Mater. 2017, 33, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yuan, T.; Xiao, Z.; Tang, P.; Xiao, Y.; Fan, Y.; Zhang, X. Hydrogels of collagen/chondroitin sulfate/hyaluronan interpenetrating polymer network for cartilage tissue engineering. J. Mater. Sci. Mater. Med. 2012, 23, 2267–2279. [Google Scholar] [CrossRef] [PubMed]
- Kérourédan, O.; Bourget, J.-M.; Rémy, M.; Crauste-Manciet, S.; Kalisky, J.; Catros, S.; Thébaud, N.B.; Devillard, R. Micropatterning of endothelial cells to create a capillary-like network with defined architecture by laser-assisted bioprinting. J. Mater. Sci. Mater. Med. 2019, 30, 1–12. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension—How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.F.; Lee, R.J.; Li, S. Chemical and physical regulation of stem cells and progenitor cells: Potential for cardio-vascular tissue engineering. Tissue Eng. 2007, 13, 1809–1823. [Google Scholar] [CrossRef]
- O’Neill, C.; Jordan, P.; Ireland, G. Evidence for two distinct mechanisms of anchorage stimulation in freshly explanted and 3T3 Swiss mouse fibroblasts. Cell 1986, 44, 489–496. [Google Scholar] [CrossRef]
- Kane, R.S.; Takayama, S.; Ostuni, E.; Ingber, D.E.; Whitesides, G.M. Patterning Proteins and Cells Using Soft Lithography, The Bio-Materials: Silver Jubilee Compendium; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Whitesides, G.M.; Ostuni, E.; Takayama, S.; Jiang, X.; Ingber, D.E. Soft Lithography in Biology and Biochemistry. Annu. Rev. Biomed. Eng. 2001, 3, 335–373. [Google Scholar] [CrossRef] [Green Version]
- Wanjare, M.; Hou, L.; Nakayama, K.H.; Kim, J.J.; Mezak, N.P.; Abilez, O.J.; Tzatzalos, E.; Wu, J.C.; Huang, N.F. Anisotropic microfibrous scaffolds enhance the organization and function of cardiomyocytes derived from induced pluripotent stem cells. Biomater. Sci. 2017, 5, 1567–1578. [Google Scholar] [CrossRef]
- Murugan, R.; Ramakrishna, S. Design Strategies of Tissue Engineering Scaffolds with Controlled Fiber Orientation. Tissue Eng. 2007, 13, 1845–1866. [Google Scholar] [CrossRef]
- Di Cio, S.; Bøggild, T.M.; Connelly, J.; Sutherland, D.S.; Gautrot, J.E. Differential integrin expression regulates cell sensing of the matrix nanoscale geometry. Acta Biomater. 2017, 50, 280–292. [Google Scholar] [CrossRef]
- Lai, E.S.; Anderson, C.M.; Fuller, G.G. Designing a tubular matrix of oriented collagen fibrils for tissue engineering. Acta Biomater. 2011, 7, 2448–2456. [Google Scholar] [CrossRef]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kawazoe, N.; Chen, G. Biomimetic Assembly of Vascular Endothelial Cells and Muscle Cells in Microgrooved Collagen Po-rous Scaffolds. Tissue Eng. Part C Methods 2017, 23, 367–376. [Google Scholar] [CrossRef]
- Hudlicka, O. What makes blood vessels grow? J. Physiol. 1991, 444, 1–24. [Google Scholar] [CrossRef]
- Wells, R.G. The role of matrix stiffness in regulating cell behavior. Hepatology 2008, 47, 1394–1400. [Google Scholar] [CrossRef]
- Xie, S.-A.; Zhang, T.; Wang, J.; Zhao, F.; Zhang, Y.-P.; Yao, W.-J.; Hur, S.S.; Yeh, Y.-T.; Pang, W.; Zheng, L.-S.; et al. Matrix stiffness determines the phenotype of vascular smooth muscle cell in vitro and in vivo: Role of DNA methyltransferase 1. Biomaterials 2018, 155, 203–216. [Google Scholar] [CrossRef]
- Qiu, Y.; Bayomy, A.F.; Gomez, M.V.; Bauer, M.; Du, P.; Yang, Y.; Zhang, X.; Liao, R. A role for matrix stiffness in the regulation of cardiac side population cell function. Am. J. Physiol. Circ. Physiol. 2015, 308, H990–H997. [Google Scholar] [CrossRef] [Green Version]
- Kural, M.H.; Billiar, K.L. Regulating tension in three-dimensional culture environments. Exp. Cell Res. 2013, 319, 2447–2459. [Google Scholar] [CrossRef] [Green Version]
- Emerman, J.T.; Burwen, S.J.; Pitelka, D.R. Substrate properties influencing ultrastructural differentiation of mammary epithelial cells in culture. Tissue Cell 1979, 11, 109–119. [Google Scholar] [CrossRef]
- Boudreau, N.; Werb, Z.; Bissell, M.J. Suppression of apoptosis by basement membrane requires three-dimensional tissue organization and withdrawal from the cell cycle. Proc. Natl. Acad. Sci. USA 1996, 93, 3509–3513. [Google Scholar] [CrossRef] [Green Version]
- Roskelley, C.D.; Desprez, P.Y.; Bissell, M.J. Extracellular matrix-dependent tissue-specific gene expression in mammary epithelial cells requires both physical and biochemical signal transduction. Proc. Natl. Acad. Sci. USA 1994, 91, 12378–12382. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R.; Ben-Ze’ev, A.; Benecke, B.J.; Penman, S. Altered translatability of messenger RNA from suspended anchorage-dependent fibroblasts: Reversal upon cell attachment to a surface. Cell 1978, 15, 627–637. [Google Scholar] [CrossRef]
- Edwards, W.S.; Mohtashemi, M.; Holdefer, W.F. The importance of proper caliber of lumen in femoral popliteal arterial reconstruction. J. Cardiovasc. Surg. 1967, 8, 195–197. [Google Scholar]
- Hess, C.N.; Lopes, R.D.; Gibson, C.M.; Hager, R.; Wojdyla, D.M.; Englum, B.; Mack, M.J.; Califf, R.M.; Kouchoukos, N.T.; Peterson, E.D.; et al. Saphenous Vein Graft Failure After Coronary Artery Bypass Surgery. Circulation 2014, 130, 1445–1451. [Google Scholar] [CrossRef]
- Angelini, G.D.; Breckenridge, I.M.; Butchart, E.G.; Armistead, S.H.; Middleton, K.M.; Henderson, A.H.; Newby, A.C. Metabolic damage to human saphenous vein during preparation for coronary artery bypass grafting. Cardiovasc. Res. 1985, 19, 326–334. [Google Scholar] [CrossRef] [Green Version]
- McKavanagh, P.; Yanagawa, B.; Zawadowski, G.; Cheema, A. Management and Prevention of Saphenous Vein Graft Failure: A Review. Cardiol. Ther. 2017, 6, 203–223. [Google Scholar] [CrossRef] [Green Version]
- Roll, S.; Müller-Nordhorn, J.; Keil, T.; Scholz, H.; Eidt, D.; Greiner, W.; Willich, S.N. Dacron® vs. PTFE as bypass materials in peripheral vascular surgery—Systematic review and meta-analysis. BMC Surg. 2008, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Toursarkissian, B.; Eisenberg, P.R.; Abendschein, D.R.; Rubin, B.G. Thrombogenicity of small-diameter prosthetic grafts: Relative contributions of graft-associated thrombin and factor Xa. J. Vasc. Surg. 1997, 25, 730–735. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.H.; Chen, C.; Bush, R.L.; Yao, Q.; Lumsden, A.B.; Hanson, S.R. Small-caliber heparin-coated ePTFE grafts reduce platelet deposition and neointimal hyperplasia in a baboon model. J. Vasc. Surg. 2004, 39, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Kidson, I.G.; Abbott, W.M. Low compliance and arterial graft occlusion. Circulation 1978, 58, I1–I4. [Google Scholar] [PubMed]
- Aussel, A.; Thébaud, N.B.; Bérard, X.; Brizzi, V.; Delmond, S.; Bareille, R.; Siadous, R.; James, C.; Ripoche, J.; Durand, M.; et al. Chitosan-based hydrogels for developing a small-diameter vascular graft: In vitro and in vivo evaluation. Biomed. Mater. 2017, 12, 065003. [Google Scholar] [CrossRef] [PubMed]
- Bracaglia, L.G.; Messina, M.; Winston, S.; Kuo, C.-Y.; Lerman, M.; Fisher, J.P. 3D Printed Pericardium Hydrogels to Promote Wound Healing in Vascular Applications. Biomacromolecules 2017, 18, 3802–3811. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.; Thottappillil, N. Scaffolds in vascular regeneration: Current status. Vasc. Health Risk Manag. 2015, 11, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Niklason, L.E.; Gao, J.; Abbott, W.M.; Hirschi, K.K.; Houser, S.; Marini, R.; Langer, R. Functional Arteries Grown in Vitro. Science 1999, 284, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Niklason, L.E.; Abbott, W.; Gao, J.; Klagges, B.; Hirschi, K.K.; Ulubayram, K.; Conroy, N.; Jones, R.; Vasanawala, A.; Sanzgiri, S.; et al. Morphologic and mechanical characteristics of engineered bovine arteries. J. Vasc. Surg. 2001, 33, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Quint, C.; Kondo, Y.; Manson, R.J.; Lawson, J.H.; Dardik, A.; Niklason, L.E. Decellularized tissue-engineered blood vessel as an arterial conduit. Proc. Natl. Acad. Sci. USA 2011, 108, 9214–9219. [Google Scholar] [CrossRef] [Green Version]
- Dahl, S.L.M.; Kypson, A.P.; Lawson, J.H.; Blum, J.L.; Strader, J.T.; Li, Y.; Manson, R.J.; Tente, W.E.; DiBernardo, L.; Hensley, M.T.; et al. Readily Available Tissue-Engineered Vascular Grafts. Sci. Transl. Med. 2011, 3, 68ra9. [Google Scholar] [CrossRef]
- Lawson, J.H.; Glickman, M.H.; Ilzecki, M.; Jakimowicz, T.; Jaroszynski, A.; Peden, E.K.; Pilgrim, A.J.; Prichard, H.L.; Guziewicz, M.; Przywara, S.; et al. Bioengineered human acellular vessels for dialysis access in patients with end-stage renal disease: Two phase 2 single-arm trials. Lancet 2016, 387, 2026–2034. [Google Scholar] [CrossRef] [Green Version]
- Dahl, S.L.M.; Rhim, C.; Song, Y.C.; Niklason, L.E. Mechanical Properties and Compositions of Tissue Engineered and Native Arteries. Ann. Biomed. Eng. 2007, 35, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Beamish, J.A.; He, P.; Kottke-Marchant, K.; Marchant, R.E. Molecular Regulation of Contractile Smooth Muscle Cell Phenotype: Implications for Vascular Tissue Engineering. Tissue Eng. Part B Rev. 2010, 16, 467–491. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Yamamoto, K.; Noumura, T. Type I Collagen Promotes Modulation of Cultured Rabbit Arterial Smooth Muscle Cells from a Contractile to a Synthetic Phenotype. Exp. Cell Res. 1993, 204, 121–129. [Google Scholar] [CrossRef]
- Khanna, A. Fabrication of Human Serum Albumin Film for Enhanced Hemocompatibility and Mitigation of Intimal Hyperplasia under Physiologically Relevant Flow Shear Conditions. Ph.D. Thesis, Clemson University, Clemson, SC, USA, 2017. [Google Scholar]
- Khanna, A. Fabrication of Human Serum Albumin film on expanded polytetrafluoroethylene (e-PTFE) for Enhanced Hemocompatibility and Adhesion Strength. In Proceedings of the Transactions of the 41st Annual Meeting of Society for Biomaterials (SFB), Minneapolis, MN, USA, 5–8 April 2017. [Google Scholar]
- Lu, X.; Khanna, A.; Luzinov, I.; Nagatomi, J.; Harman, M. Surface modification of polypropylene surgical meshes for improving adhesion with poloxamine hydrogel adhesive. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1047–1055. [Google Scholar] [CrossRef]
- Koobatian, M.T.; Row, S.; Smith, R.J., Jr.; Koenigsknecht, C.; Andreadis, S.T.; Swartz, D.D. Successful endothelialization and remodeling of a cell-free small-diameter arterial graft in a large animal model. Biomaterials 2015, 76, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Roeder, R.; Wolfe, J.; Lianakis, N.; Hinson, T.; Geddes, L.A.; Obermiller, J. Compliance, elastic modulus, and burst pressure of small-intestine submucosa (SIS), small-diameter vascular grafts. J. Biomed. Mater. Res. 1999, 47, 65–70. [Google Scholar] [CrossRef]
- Coakley, D.N.M.; Shaikh, F.; O’Sullivan, K.; Kavanagh, E.G.; Grace, P.; Walsh, S.; McGloughlin, T. Comparing the endothelialisation of extracellular matrix bioscaffolds with coated synthetic vascular graft materials. Int. J. Surg. 2015, 25, 31–37. [Google Scholar] [CrossRef]
- Peng, H.; Schlaich, E.M.; Row, S.; Andreadis, S.T.; Swartz, D.D. A Novel Ovine ex vivo Arteriovenous Shunt Model to Test Vascular Implantability. Cells Tissues Organs 2012, 195, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Nasiri, B.; Row, S.; Smith, R.J., Jr.; Swartz, D.D.; Andreadis, S.T. Cell-Free Vascular Grafts That Grow with the Host. Adv. Funct. Mater. 2020, 30, 2005769. [Google Scholar] [CrossRef]
- Row, S.; Peng, H.; Schlaich, E.M.; Koenigsknecht, C.; Andreadis, S.T.; Swartz, D.D. Arterial grafts exhibiting unprecedented cellular infiltration and remodeling in vivo: The role of cells in the vascular wall. Biomaterials 2015, 50, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Xue, F.; Fan, C.; Mo, X. Surface heparinization and blood compatibility modification of small intestinal submucosa (SIS) for small-caliber vascular regeneration. Bio-Med. Mater. Eng. 2017, 28, 213–222. [Google Scholar] [CrossRef]
- Ma, X.; He, Z.; Li, L.; Liu, G.; Li, Q.; Yang, D.; Zhang, Y.; Li, N. Development and in vivo validation of tissue-engineered, small-diameter vascular grafts from decellularized aortae of fetal pigs and canine vascular endothelial cells. J. Cardiothorac. Surg. 2017, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Sanyour, H.; Remund, T.; Kelly, P.; Hong, Z. Vascular extracellular matrix and fibroblasts-coculture directed differentiation of human mesenchymal stem cells toward smooth muscle-like cells for vascular tissue engineering. Mater. Sci. Eng. C 2018, 93, 61–69. [Google Scholar] [CrossRef]
- Kong, X.; Kong, C.; Wen, S.; Shi, J. The use of heparin, bFGF, and VEGF 145 grafted acellular vascular scaffold in small diameter vascular graft. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 107, 672–679. [Google Scholar] [CrossRef]
- Filova, E.; Steinerova, M.; Travnickova, M.; Knitlova, J.; Musilkova, J.; Eckhardt, A.; Hadraba, D.; Matejka, R.; Prazak, S.; Stepanovska, J.; et al. Accelerated in vitro recellularization of decellularized porcine pericardium for cardiovascular grafts. Biomed. Mater. 2021, 16, 025024. [Google Scholar] [CrossRef]
- Curtis, M.W.; Russell, B. Cardiac tissue engineering. J. Cardiovasc. Nurs. 2009, 24, 87. [Google Scholar] [CrossRef] [Green Version]
- Melhem, M.R.; Park, J.; Knapp, L.; Reinkensmeyer, L.; Cvetkovic, C.; Flewellyn, J.; Lee, M.K.; Jensen, T.W.; Bashir, R.; Kong, H.; et al. 3D printed stem-cell-laden, micro-channeled hydrogel patch for the enhanced release of cell-secreting factors and treatment of myocardial infarctions. ACS Biomater. Sci. Eng. 2017, 3, 1980–1987. [Google Scholar] [CrossRef]
- Sun, Z.; Lee, S.-Y. A systematic review of 3-D printing in cardiovascular and cerebrovascular diseases. Anatol. J. Cardiol. 2017, 17, 423. [Google Scholar] [CrossRef]
- Marieb, E.N.; Hoehn, K. Human Anatomy & Physiology; Pearson Education: San Francisco, CA, USA, 2007. [Google Scholar]
- Zhang, Y.S.; Arneri, A.; Bersini, S.; Shin, S.-R.; Zhu, K.; Goli-Malekabadi, Z.; Aleman, J.; Colosi, C.; Busignani, F.; Dell’Erba, V.; et al. Bioprinting 3D microfibrous scaffolds for engineering endothelialized myocardium and heart-on-a-chip. Biomaterials 2016, 110, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Heng, B.C.; Haider, H.K.; Sim, E.K.W.; Cao, T.; Ng, S.C. Strategies for directing the differentiation of stem cells into the cardiomyogenic line age in vitro. Cardiovasc. Res. 2004, 62, 34–42. [Google Scholar] [CrossRef]
- Jang, J.; Park, H.-J.; Kim, S.-W.; Kim, H.; Park, J.Y.; Na, S.J.; Kim, H.J.; Park, M.N.; Choi, S.H.; Park, S.H.; et al. 3D printed complex tissue construct using stem cell-laden decellularized extracellular matrix bioinks for cardiac repair. Biomaterials 2016, 112, 264–274. [Google Scholar] [CrossRef]
- Gao, G.; Lee, J.H.; Jang, J.; Lee, D.H.; Kong, J.S.; Kim, B.S.; Choi, Y.J.; Jang, W.B.; Hong, Y.J.; Kwon, S.M.; et al. Tissue engineered bio-blood-vessels constructed using a tissue-specific bioink and 3D coaxial cell printing technique: A novel therapy for ischemic disease. Adv. Funct. Mater. 2017, 27, 1700798. [Google Scholar] [CrossRef]
- Shin, Y.; Han, S.; Jeon, J.; Yamamoto, K.; Zervantonakis, I.; Sudo, R.; Kamm, R.D.; Chung, S. Microfluidic assay for simultaneous culture of multiple cell types on surfaces or within hydrogels. Nat. Protoc. 2012, 7, 1247–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, T.J.; Jallerat, Q.; Palchesko, R.N.; Park, J.H.; Grodzicki, M.S.; Shue, H.-J.; Ramadan, M.H.; Hudson, A.R.; Feinberg, A.W. Three-dimensional printing of complex biological structures by freeform reversible embedding of suspended hydrogels. Sci. Adv. 2015, 1, e1500758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastikhina, O.; Moon, B.-U.; Williams, K.; Hatkar, R.; Gustafson, D.; Mourad, O.; Sun, X.; Koo, M.; Lam, A.Y.; Sun, Y.; et al. Human cardiac fibrosis-on-a-chip model recapitulates disease hallmarks and can serve as a platform for drug testing. Biomaterials 2020, 233, 119741. [Google Scholar] [CrossRef] [PubMed]
- Ott, H.C.; Matthiesen, T.S.; Goh, S.K.; Black, L.D.; Kren, S.M.; Netoff, T.I.; Taylor, D.A. Perfusion-decellularized matrix: Using na-ture’s platform to engineer a bioartificial heart. Nat. Med. 2008, 14, 213–221. [Google Scholar] [CrossRef]
- Li, Q.; Williams, C.G.; Sun, D.D.N.; Wang, J.; Leong, K.; Elisseeff, J.H. Photocrosslinkable polysaccharides based on chondroitin sulfate. J. Biomed. Mater. Res. 2003, 68A, 28–33. [Google Scholar] [CrossRef]
- Nettles, D.L.; Vail, T.P.; Morgan, M.T.; Grinstaff, M.; Setton, L.A. Photocrosslinkable Hyaluronan as a Scaffold for Articular Cartilage Repair. Ann. Biomed. Eng. 2004, 32, 391–397. [Google Scholar] [CrossRef]
- Vedadghavami, A.; Minooei, F.; Mohammadi, M.H.; Khetani, S.; Kolahchi, A.R.; Mashayekhan, S.; Sanati-Nezhad, A. Manufacturing of hydrogel biomaterials with controlled mechanical properties for tissue engineering applications. Acta Biomater. 2017, 62, 42–63. [Google Scholar] [CrossRef]
- Cummings, C.L.; Gawlitta, D.; Nerem, R.M.; Stegemann, J.P. Properties of engineered vascular constructs made from collagen, fibrin, and collagen–fibrin mixtures. Biomaterials 2004, 25, 3699–3706. [Google Scholar] [CrossRef]
- Hinderer, S.; Layland, S.L.; Schenke-Layland, K. ECM and ECM-like materials—Biomaterials for applications in regenerative medicine and cancer therapy. Adv. Drug Deliv. Rev. 2016, 97, 260–269. [Google Scholar] [CrossRef]
- Johnson, K.A.; Rogers, G.J.; Roe, S.C.; Howlett, C.R.; Clayton, M.K.; Milthorpe, B.K.; Schindhelm, K. Nitrous acid pre-treatment of tendon xenografts cross-linked with glutaraldehyde and sterilized with gamma irradiation. Biomaterials 1999, 20, 1003–1015. [Google Scholar] [CrossRef]
- Roe, S.; Milthorpe, B.; True, K.; Rogers, G.; Schindhelm, K. The effect of gamma irradiation on a xenograft tendon bioprosthesis. Clin. Mater. 1992, 9, 149–154. [Google Scholar] [CrossRef]
- Cheung, D.T.; Perelman, N.; Tong, D.; Nimni, M.E. The effect of gamma-irradiation on collagen molecules, isolated alpha-chains, and crosslinked native fibers. J. Biomed. Mater. Res. 1990, 24, 581–589. [Google Scholar] [CrossRef]
- Möllers, S.; Heschel, I.; Damink, L.H.O.; Schügner, F.; Deumens, R.; Müller, B.; Bozkurt, A.; Nava, J.G.; Noth, J.; Brook, G.A. Cytocompatibility of a Novel, Longitudinally Microstructured Collagen Scaffold Intended for Nerve Tissue Repair. Tissue Eng. Part A 2009, 15, 461–472. [Google Scholar] [CrossRef]
- Huang, N.; Yu, J.; Sievers, R.; Li, S.; Lee, R.J. Injectable Biopolymers Enhance Angiogenesis after Myocardial Infarction. Tissue Eng. 2005, 11, 1860–1866. [Google Scholar] [CrossRef]
- Zaitseva, T.; Yang, G.; Dionyssiou, D.; Zamani, M.; Sawamura, S.; Yakubov, E.; Ferguson, J.; Hallett, R.L.; Fleischmann, D.; Paukshto, M.V.; et al. Delivery of hepatocyte growth factor mrna from nanofibrillar scaffolds in a pig model of peripheral arterial disease. Regen. Med. 2020, 15, 1761–1773. [Google Scholar] [CrossRef]
- Hume, P.; He, J.; Haskins, K.; Anseth, K.S. Strategies to reduce dendritic cell activation through functional biomaterial design. Biomaterials 2012, 33, 3615–3625. [Google Scholar] [CrossRef] [Green Version]
- Hume, P.S.; Anseth, K.S. Inducing local T cell apoptosis with anti-Fas-functionalized polymeric coatings fabricated via sur-face-initiated photopolymerizations. Biomaterials 2010, 31, 3166–3174. [Google Scholar] [CrossRef] [Green Version]
- Hume, P.S.; Bowman, C.N.; Anseth, K.S. Functionalized PEG hydrogels through reactive dip-coating for the formation of immunoactive barriers. Biomaterials 2011, 32, 6204–6212. [Google Scholar] [CrossRef] [Green Version]
- Kristofik, N.J.; Qin, L.; Calabro, N.E.; Dimitrievska, S.; Li, G.; Tellides, G.; Niklason, L.E.; Kyriakides, T.R. Improving in vivo outcomes of decellularized vascular grafts via incorporation of a novel extracellular matrix. Biomaterials 2017, 141, 63–73. [Google Scholar] [CrossRef]


{kind=link}
| ECM | ECM-Based Biomaterials | Model | Cardiovascular Tissue Engineering Advantages | Ref. |
|---|---|---|---|---|
| Collagen | COL I | Murine | Cardiomyocyte differentiation, maturation and contractile function | [90] |
| Collagen | ESC and iPSC/COL IV | In vitro | Differentiation of induced pluripotent stem cells (iPSs) into cardiomyocytes of contractile function. | [91] |
| Collagen and Fibrin | COL 1/Fibrin | In vitro | Improved physical property, cardiac tissue compaction | [63] |
| Collagen and Elastin | COL 1/Elastin | In vitro | Enhanced elasticity, maturation of valve interstitial cells and valve ECs. | [65] |
| Fibrin | Fibrin | In vitro | Cardiomyocyte proliferation and cardiac regeneration | [92] |
| Collagen | COL 1/Growth factors/MatrigelTM | In vitro | Cardiomyocyte differentiation and maturation | [93] |
| HA | HA | In vitro | Attenuates cardiac fibrosis and promote cardiac muscle tissue regeneration | [94,95] |
| Tissue/Organ | Decellularization Method | Model | Vascular Response | Ref. |
|---|---|---|---|---|
| Adult Porcine Heart | Pulsatile retrograde aortic perfusion | Porcine | Chicken cardiomyocytes were cultured. ECM retained collagen, elastin, glycosaminoglycans, and mechanical integrity. | [96] |
| Porcine Whole Heart | Perfusion of Trypsin/EDTA and Triton-X100/deoxycholic acid (DCA) | Porcine | ECM retained elastin, collagen, and proteoglycan | [97] |
| Rat Heart Adult | Three decellularization solutions were used: (1) SDS/TritonX100- v/s (2) Trypsin plus Triton/DCA v/s (3) SDS/DCA/saponin | In vitro | C2C12 myoblasts were seeded in vitro. ECM retained laminin in all groups, elastin in groups 1 and 2, collagen in groups 1 and 3. | [98] |
| Porcine Aortic Valves | Triton X-100 | In vitro | ECs were seeded. EC mediated ECM deposition was observed. | [99] |
| Human Umbilical Artery | SDS treatment | In vitro/In vivo | In vitro EC cell seeding and implantation in rats. EC culture growth was sustained for 8 weeks, ECM preserved. | [100] |
| Tissue Engineered Porcine Vessels | SDS | In vitro/Porcine | Pre-seeding of EC progenitor cells and implantation in porcine carotid artery. No clotting observed. No signs of intimal hyperplasia. | [101] |
| Bioartificial Tissue Engineered Heart | Coronary perfusion-based whole organ decellularization | In vitro/rat model | Recellularization with neonatal cardiac cells and rat aortic ECs. Constructs exhibited contractile function and EC culture growth. | [77,78] |
| Neonatal Mouse Heart | Liquid nitrogen, erythrocyte lysis, and DNA/RNA removal | In vivo model of acute MI | nmECM showed improved heart function compared to adult heart derived ECM, cardiac repair after MI | [79] |
| Cardiac Porcine Hearts | SDS | In vitro | cdECM promotes human cardiac fibroblast culture and human iPSC-derived cardiomyocytes. | [102] |
| Porcine Cardiac Tissue | SDS | In vivo | Decellularized porcine myocardial extracellular matrix (dECM)-reduced graphene oxide hydrogel promoted increased expression of genes that regulated contractile function. | [103] |
| Porcine Heart | SDS | In vitro | Enhanced maturation of cardiomyocytes (isolated from neonatal rats) in hdECM | [104] |
| Rat Heart Tissue | Ionic and non-ionic detergents | In vivo | (AdMSCs) promoted increased cardiomyocyte-specific gene expression for 2–4 weeks. | [105] |
| ECM Component | Model | Endothelial Cell Response | Ref. |
|---|---|---|---|
| Laminin | In vitro | Differentiation of embryonic stem cells into functional endothelial progenitor cells. | [114] |
| Laminin | In vitro | Laminin facilitates in vitro 3D vascular network formation by promoting uptake of VEGF by ECs. | [115] |
| COL IV- multi-component ECM | In vitro | Improved endothelial differentiation of human induced pluripotent stem cells. | [118] |
| Fibronectin | In vitro | EC growth and proliferation was supported by fibronectin coating on vascular grafts. | [121] |
| RGD-modified HA | In vivo | EC migration and proliferation, formation of functional vasculature | [122] |
| dECM | In vitro | endothelial progenitor cells encapsulated in blood vessel-derived dECM exhibited higher proliferation capability and enhanced vascular network formation. | [123] |
| dECM | In vitro | In vitro EC cell seeding and implantation in rats. EC culture growth was sustained for 8 weeks, ECM preserved. | [100] |
| dECM | In vivo | Hybrid ECM promoted proliferation and migration of HUVECs, significantly inhibited immune response and calcification, exhibited stability and biocompatibility compared to non-hybrid leaflet. | [124] |
| Heparin | In vitro | Surfaces covalently immobilized with heparin promoted endothelial cell growth and inhibited SMCs. | [125] |
| Fb/COL/LA/FN | In vitro | High EC cell densities were achieved in 7 days of culture | [126] |
| Fibrin Fragment E (FbnE) | In vitro | Increased adhesion and endothelial differentiation. | [127] |
| COL Coated PCL Membrane | In vitro | Continuous EC monolayer was observed on collagen coated membrane. ECs exhibited filopodia protruding from lamellipodia in the junctional areas on the collagen-coated membranes. | [128] |
| Matrigel Matrix Comprising of LA, COL IV, Heparen Sulfate Proteoglycans | Ischemic mouse model | Improved neovasculature formation, promote cell growth, proliferation and differentiation of ECs. | [129] |
| Cardiogel Composed of LA, FN and Interstitial COL I and IV | In vivo | ECM components promote growth of ECs and CMs, spontaneous contractile activity and phenotypic morphological differentiation. | [130] |
| 3D Bioprinting Technique | Advantages | Limitations | Ref. |
|---|---|---|---|
| Inkjet Bioprinting | Uses thermal, electromagnetic or piezoelectric technology to deposit droplets of “ink” (materials) Rapid printing speeds and high resolution. Capable of printing low-viscosity biomaterials. Availability and ease of replacement of bio-inks. High-cell viability and relatively low cost | Low material viscosity (<10 Pa·s) and low droplet directionality. Lack of precision with respect to droplet size. Requirement for low viscosity bio-ink. Nozzle clogging and cellular distortion due to high-cell density. Low mechanical strength. Inability to provide continuous stream of material. | [159] |
| Micro-Extrusion | Ability to print biomaterials with high cell densities (higher than 1 × 106 cells mL−1) comparable to physiological cell densities. Can produce continuous stream of material. Can successfully print high viscosity bio-inks such as polymers, clay-based substrates. | Low printing resolution (>100 µm) and slow printing speeds. Loss of cellular viability and distortion of cellular structure due to the pressure to expel the bio-ink. | [160] |
| Laser-Assisted Bioprinting: SLA and LIFT | Rapid printing speeds and ability to print biomaterials with wide range of viscosities (1–300 mPa/s). High degree of precision and resolution (1 cell/droplet). Can successfully print high density of cells 108 mL−1 | Time consuming: need to prepare reservoirs/ribbons. Lower cellular viability compared to other methods. Loss of cells due to thermal damage. SLA requires intense UV radiation for crosslinking process. Requires large amount of material. High cost. Long post processing time and fewer materials compatible with SLA. | [161,162] |
| Material | Treatment | Model | Vascular Graft Response | Ref. |
|---|---|---|---|---|
| PGA Scaffold | SMC | Bovine | Grafts exhibited goof vessel wall thickness, burst pressure, and collagen content. | [194,195] |
| PGA Scaffold | SMC | Canine | Grafts exhibited good long-term patency for 8–10 weeks. | [197] |
| PGA with Fibrinogen and Thrombin | SMC s and ECs derived from hiPSC | Porcine | Grafts exhibited endothelial differentiation. | [194,196] |
| PGA Scaffold | MSCs | In vitro | Grafts exhibited superior mechanical properties and cellular growth. | [198] |
| Fibrin Gel | Human fibroblast (hDFs) | Baboon | Grafts exhibited higher patency rates of >80%. | [196] |
| Material | Treatment | Model | Vascular Graft Response | Ref. |
|---|---|---|---|---|
| SIS | EC culture | In vitro | Grafts exhibited higher EC proliferation and cobblestone morphology. | [207] |
| SIS | Pre-seeded with SMC and fibrinogen/thrombin | Porcine | Grafts exhibited endothelial cell attachment and graft patency. | [208] |
| SIS | Heparin | Sheep | Grafts exhibited lumens with no sign of clotting or intimal hyperplasia. | [194,196] |
| Pericardium | MSCs | Bovine | ECM and growth factors facilitated differentiation into ECs. | [210] |
| Porcine SIS Tubes | Heparin/VEGF | Sheep | Grafts exhibited long term patency rates for 3 months with a confluent endothelium and no signs of thrombosis. | [211] |
| Material | Treatment | Model | Vascular Graft Response | Ref. |
|---|---|---|---|---|
| Carotid Artery | Autologous EC | Porcine | Grafts exhibited patency rates of >90% for 6 months. | [212] |
| Carotid Artery | MSCs | Porcine | ECM and growth factors cause differentiation of MSCs into ECs. | [213] |
| Carotid Artery | Heparin | Porcine | Grafts exhibited lumens with no sign of clotting/thrombus. | [214] |
| Carotid Artery | MSCs | Porcine | ECM and growth factors cause differentiation of MSCs into SMCs. | [213] |
| Porcine Aorta | Autologous ECs | Canine | Grafts exhibited long term patency rates for 3 months. | [210] |
| Aorta | Heparin/VEGF | Canine | Grafts exhibited patency of >90% post 2 years. | [214] |
| Porcine Pericardium Scaffold | Fibrin mesh/Heparin/VEGF | Porcine | Grafts exhibited potential to accelerate in situ endothelialization. | [215] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanna, A.; Zamani, M.; Huang, N.F. Extracellular Matrix-Based Biomaterials for Cardiovascular Tissue Engineering. J. Cardiovasc. Dev. Dis. 2021, 8, 137. https://doi.org/10.3390/jcdd8110137
Khanna A, Zamani M, Huang NF. Extracellular Matrix-Based Biomaterials for Cardiovascular Tissue Engineering. Journal of Cardiovascular Development and Disease. 2021; 8(11):137. https://doi.org/10.3390/jcdd8110137
Chicago/Turabian StyleKhanna, Astha, Maedeh Zamani, and Ngan F. Huang. 2021. "Extracellular Matrix-Based Biomaterials for Cardiovascular Tissue Engineering" Journal of Cardiovascular Development and Disease 8, no. 11: 137. https://doi.org/10.3390/jcdd8110137
APA StyleKhanna, A., Zamani, M., & Huang, N. F. (2021). Extracellular Matrix-Based Biomaterials for Cardiovascular Tissue Engineering. Journal of Cardiovascular Development and Disease, 8(11), 137. https://doi.org/10.3390/jcdd8110137