
The Left Ventricular Myocardium in Hypoplastic Left Heart Syndrome

 ,
, 
{kind=link}
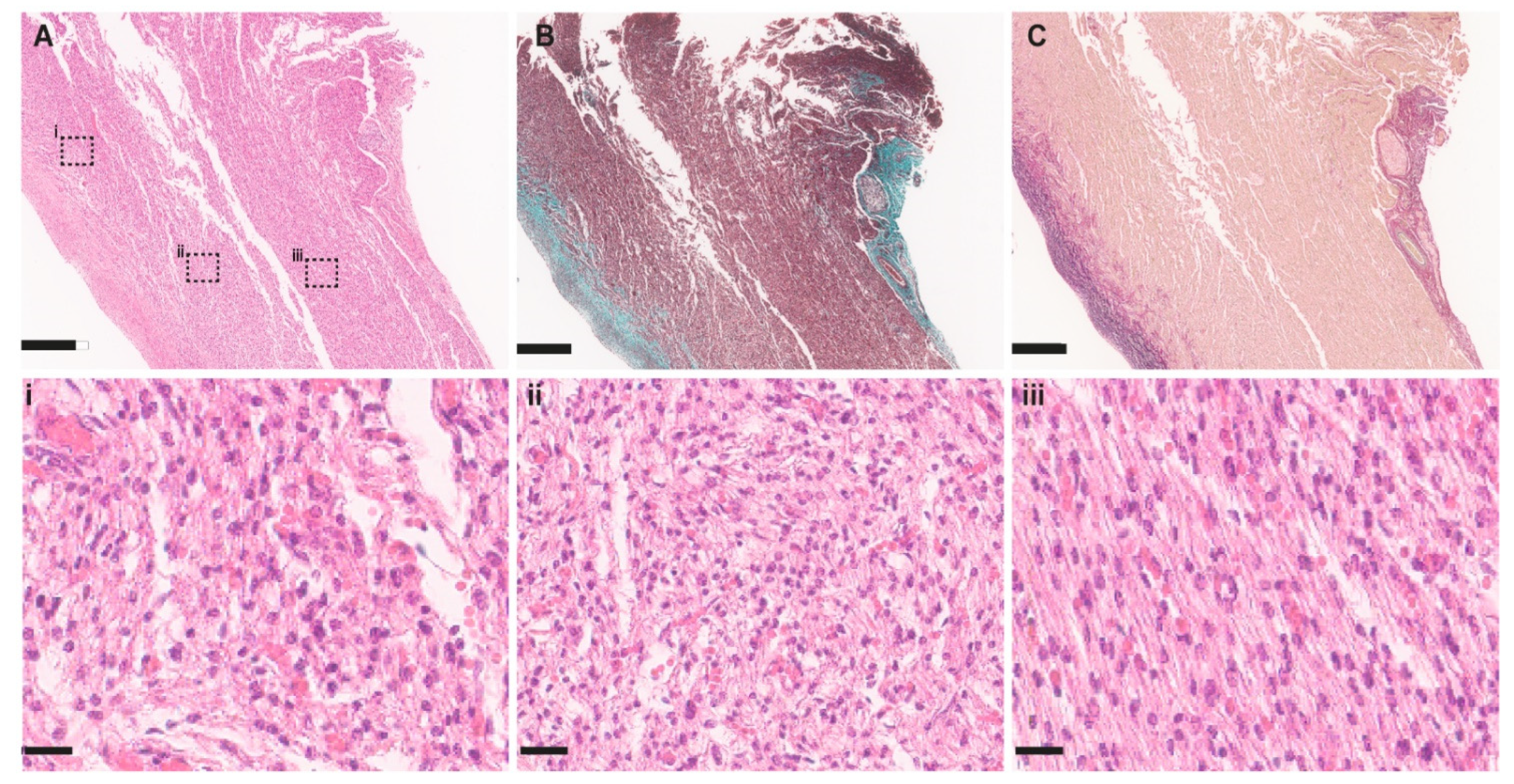{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Morphological Sub-Types of HLHS
3. The Developmental Origins of the Normal Left Ventricular Myocardium
4. The Myocardial Architecture in HLHS
5. Cardiomyocyte Organisation
6. Cardiomyocyte Ultrastructure in the Slit-like and Peach-like Ventricles
7. Endocardial Fibroelastosis in Peach-like Ventricles
8. Histological Evidence of Myocardial Conditions Leading to HLHS
9. Vascular Abnormalities in HLHS
10. Conclusions and Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kearney, D.L. The pathological spectrum of left-ventricular hypoplasia. Semin. Cardiothorac. Vasc. Anesth. 2013, 17, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Crucean, A.; Alqahtani, A.; Barron, D.J.; Brawn, W.J.; Richardson, R.V.; O’Sullivan, J.; Anderson, R.H.; Henderson, D.J.; Chaudhry, B. Re-evaluation of hypoplastic left heart syndrome from a developmental and morphological perspective. Orphanet. J. Rare Dis. 2017, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.C.G.; Béland, M.J.; Colan, S.D.; Walters, H.L.; Aiello, V.D.; Anderson, R.H.; Bailliard, F.; Boris, J.R.; Cohen, M.S.; Gaynor, J.W.; et al. Nomenclature for congenital and paediatric cardiac disease: The International Paediatric and Congenital Cardiac Code (IPCCC) and the Eleventh Iteration of the International Classification of Diseases (ICD-11). Cardiol. Young 2017, 27, 1872–1938. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.H.; Gupta, D.; Bleiweis, M.; Backer, C.L.; Anderson, R.H.; Spicer, D.E. Pathologic Characteristics of 119 Archived Specimens Showing the Phenotypic Features of Hypoplastic Left Heart Syndrome. Semin. Thorac. Cardiovasc. Surg. 2020, 32, 895–903. [Google Scholar] [CrossRef]
- Samánek, M.; Slavík, Z.; Zborilová, B.; Hrobonová, V.; Vorísková, M.; Skovránek, J. Prevalence, treatment, and outcome of heart disease in live-born children: A prospective analysis of 91,823 live-born children. Pediatr. Cardiol. 1989, 10, 205–211. [Google Scholar] [CrossRef]
- Cole, C.R.; Eghtesady, P. The myocardial and coronary histopathology and pathogenesis of hypoplastic left heart syndrome. Cardiol. Young 2016, 26, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Grossfeld, P.; Nie, S.; Lin, L.; Wang, L.; Anderson, R.H. Hypoplastic Left Heart Syndrome: A New Paradigm for an Old Disease? J. Cardiovasc. Dev. Dis. 2019, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Lev, M. Pathologic anatomy and interrelationship of hypoplasia of the aortic tract complexes. Lab. Investig. 1952, 1, 61–70. [Google Scholar]
- Noonan, J.A.; Nadas, A.S. The hypoplastic left heart syndrome; an analysis of 101 cases. Pediatr. Clin. N. Am. 1958, 5, 1029–1056. [Google Scholar] [CrossRef]
- Tchervenkov, C.I.; Jacobs, J.P.; Weinberg, P.M.; Aiello, V.D.; Béland, M.J.; Colan, S.D.; Elliott, M.J.; Franklin, R.C.; Gaynor, J.W.; Krogmann, O.N.; et al. The nomenclature, definition and classification of hypoplastic left heart syndrome. Cardiol. Young 2006, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, B.; Henderson, D.; Anderson, R. Double-outlet right ventricle is not hypoplastic left heart syndrome. Nat. Genet. 2019, 51, 198. [Google Scholar] [CrossRef] [PubMed]
- Tchervenkov, C.I.; Jacobs, M.L.; Tahta, S.A. Congenital Heart Surgery Nomenclature and Database Project: Hypoplastic left heart syndrome. Ann. Thorac. Surg. 2000, 69 (Suppl. 4), S170–S179. [Google Scholar] [CrossRef]
- Sharland, G.K.; Chita, S.K.; Fagg, N.L.; Anderson, R.H.; Tynan, M.; Cook, A.C.; Allan, L.D. Left ventricular dysfunction in the fetus: Relation to aortic valve anomalies and endocardial fibroelastosis. Br. Heart J. 1991, 66, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Feinstein, J.A.; Benson, D.W.; Dubin, A.M.; Cohen, M.S.; Maxey, D.M.; Mahle, W.T.; Pahl, E.; Villafañe, J.; Bhatt, A.B.; Peng, L.F.; et al. Hypoplastic left heart syndrome: Current considerations and expectations. J. Am. Coll. Cardiol. 2012, 59 (Suppl. 1), S1–S42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhry, B.; Ramsbottom, S.; Henderson, D.J. Genetics of cardiovascular development. Prog. Mol. Biol. Transl. Sci. 2014, 124, 19–41. [Google Scholar] [CrossRef]
- Henderson, D.J.; Anderson, R.H. The development and structure of the ventricles in the human heart. Pediatr. Cardiol. 2009, 30, 588–596. [Google Scholar] [CrossRef]
- Tian, X.; Li, Y.; He, L.; Zhang, H.; Huang, X.; Liu, Q.; Pu, W.; Zhang, L.; Li, Y.; Zhao, H.; et al. Identification of a hybrid myocardial zone in the mammalian heart after birth. Nat. Commun. 2017, 8, 87. [Google Scholar] [CrossRef]
- Bulkley, B.H.; Weisfeldt, M.L.; Hutchins, G.M. Asymmetric septal hypertrophy and myocardial fiber disarray. Features of normal, developing, and malformed hearts. Circulation 1977, 56, 292–298. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, W.N.; Cash, J.B.; Cottrill, C.M.; Johnson, G.L.; Noonan, J.A. Ventriculocoronary connections in hypoplastic left hearts: An autopsy microscopic study. Circulation 1982, 66, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Bulkley, B.H.; D’Amico, B.; Taylor, A.L. Extensive myocardial fiber disarray in aortic and pulmonary atresia. Relevance to hypertrophic cardiomyopathy. Circulation 1983, 67, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Bohlmeyer, T.J.; Helmke, S.; Ge, S.; Lynch, J.; Brodsky, G.; Sederberg, J.H.; Robertson, A.D.; Minobe, W.; Bristow, M.R.; Perryman, M.B. Hypoplastic left heart syndrome myocytes are differentiated but possess a unique phenotype. Cardiovasc. Pathol. 2003, 12, 23–31. [Google Scholar] [CrossRef]
- Gaber, N.; Gagliardi, M.; Patel, P.; Kinnear, C.; Zhang, C.; Chitayat, D.; Shannon, P.; Jaeggi, E.; Tabori, U.; Keller, G.; et al. Fetal reprogramming and senescence in hypoplastic left heart syndrome and in human pluripotent stem cells during cardiac differentiation. Am. J. Pathol. 2013, 183, 720–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, C.; Martinez, A.M.; Zuppan, C.W.; Shah, M.M.; Bailey, L.L.; Fletcher, W.H. Identification of connexin43 (alpha1) gap junction gene mutations in patients with hypoplastic left heart syndrome by denaturing gradient gel electrophoresis (DGGE). Mutat. Res. 2001, 479, 173–186. [Google Scholar] [CrossRef]
- Mahtab, E.A.; Gittenberger-de Groot, A.C.; Vicente-Steijn, R.; Lie-Venema, H.; Rijlaarsdam, M.E.; Hazekamp, M.G.; Bartelings, M.M. Disturbed myocardial connexin 43 and N-cadherin expressions in hypoplastic left heart syndrome and borderline left ventricle. J. Thorac. Cardiovasc. Surg. 2012, 144, 1315–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehbens, W.E.; Delahunt, B.; Zuccollo, J.M. The histopathology of endocardial sclerosis. Cardiovasc. Pathol. 2000, 9, 161–173. [Google Scholar] [CrossRef]
- Lurie, P.R. Changing concepts of endocardial fibroelastosis. Cardiol. Young 2010, 20, 115–123. [Google Scholar] [CrossRef]
- Griffin, L.D.; Kearney, D.; Ni, J.; Jaffe, R.; Fricker, F.J.; Webber, S.; Demmler, G.; Gelb, B.D.; Towbin, J.A. Analysis of formalin-fixed and frozen myocardial autopsy samples for viral genome in childhood myocarditis and dilated cardiomyopathy with endocardial fibroelastosis using polymerase chain reaction (PCR). Cardiovasc. Pathol. 1995, 4, 3–11. [Google Scholar] [CrossRef]
- Ni, J.; Bowles, N.E.; Kim, Y.H.; Demmler, G.; Kearney, D.; Bricker, J.T.; Towbin, J.A. Viral infection of the myocardium in endocardial fibroelastosis. Molecular evidence for the role of mumps virus as an etiologic agent. Circulation 1997, 95, 133–139. [Google Scholar] [CrossRef]
- Aiello, V.; De Lourdes Higuchi, M. Frequency and severity of endocardial fibroelastosis in dilated hearts from children—Endocardial thickness is inversely correlated to age. Cardiol. Young 1994, 4, 117–121. [Google Scholar] [CrossRef]
- Carceller, A.M.; Maroto, E.; Fouron, J.C. Dilated and contracted forms of primary endocardial fibroelastosis: A single fetal disease with two stages of development. Br. Heart J. 1990, 63, 311–313. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Takemura, G.; Kanamori, H.; Tsujimoto, A.; Goto, K.; Kawamura, I.; Watanabe, T.; Morishita, K.; Miyazaki, N.; Tanaka, T.; et al. Phenotype and physiological significance of the endocardial smooth muscle cells in human failing hearts. Circ. Heart Fail. 2015, 8, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weixler, V.; Marx, G.R.; Hammer, P.E.; Emani, S.M.; Del Nido, P.J.; Friehs, I. Flow disturbances and the development of endocardial fibroelastosis. J. Thorac. Cardiovasc. Surg. 2020, 159, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Friehs, I.; Zhong Hu, T.; Melnychenko, I.; Tampe, B.; Alnour, F.; Iascone, M.; Kalluri, R.; Zeisberg, M.; Del Nido, P.J.; et al. Endocardial fibroelastosis is caused by aberrant endothelial to mesenchymal transition. Circ. Res. 2015, 116, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, E.S.; Pepper, V.K.; Best, C.A.; Onwuka, E.A.; Yi, T.; Tara, S.; Cianciolo, R.; Baker, P.; Shinoka, T.; Breuer, C.K. A mouse model of endocardial fibroelastosis. Cardiovasc. Pathol. 2015, 24, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, S.; Robles, C.; Illigens, B.M.; Casar Berazaluce, A.M.; del Nido, P.J.; Friehs, I. Distention of the Immature Left Ventricle Triggers Development of Endocardial Fibroelastosis: An Animal Model of Endocardial Fibroelastosis Introducing Morphopathological Features of Evolving Fetal Hypoplastic Left Heart Syndrome. Biomed. Res. Int. 2015, 2015, 462469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Huang, X.; Liu, K.; Tang, J.; He, L.; Pu, W.; Liu, Q.; Li, Y.; Tian, X.; Wang, Y.; et al. Fibroblasts in an endocardial fibroelastosis disease model mainly originate from mesenchymal derivatives of epicardium. Cell Res. 2017, 27, 1157–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, P.; Christia, P.; Saxena, A.; Su, Y.; Frangogiannis, N.G. Lack of specificity of fibroblast-specific protein 1 in cardiac remodeling and fibrosis. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1363–H1372. [Google Scholar] [CrossRef]
- Anfinson, M.; Fitts, R.H.; Lough, J.W.; James, J.M.; Simpson, P.M.; Handler, S.S.; Mitchell, M.E.; Tomita-Mitchell, A. Significance of α-Myosin Heavy Chain (MYH6) Variants in Hypoplastic Left Heart Syndrome and Related Cardiovascular Diseases. J. Cardiovasc. Dev. Dis. 2022, 9, 144. [Google Scholar] [CrossRef]
- Theis, J.L.; Hu, J.J.; Sundsbak, R.S.; Evans, J.M.; Bamlet, W.R.; Qureshi, M.Y.; O’Leary, P.W.; Olson, T.M. Genetic Association Between Hypoplastic Left Heart Syndrome and Cardiomyopathies. Circ. Genom. Precis. Med. 2021, 14, e003126. [Google Scholar] [CrossRef]
- Levesque, K.; Morel, N.; Maltret, A.; Baron, G.; Masseau, A.; Orquevaux, P.; Piette, J.C.; Barriere, F.; Le Bidois, J.; Fermont, L.; et al. Description of 214 cases of autoimmune congenital heart block: Results of the French neonatal lupus syndrome. Autoimmun. Rev. 2015, 14, 1154–1160. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, L.; Fei, Y.; Li, Y.; Hao, D.; Liu, Y.; Zhao, Y. Pregnancy outcome of 126 anti-SSA/Ro-positive patients during the past 24 years—A retrospective cohort study. Clin. Rheumatol. 2015, 34, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Strzelecka, I.; Biedrzycka, M.; Karuga, F.F.; Szmyd, B.; Batarowicz, K.; Respondek-Liberska, M.J. Seasonality of Hypoplastic Left Heart Syndrome and Single Ventricle Heart in Poland in the Context of Air Pollution. Clin. Med. 2021, 10, 3207. [Google Scholar] [CrossRef] [PubMed]
- Eghtesady, P. Hypoplastic left heart syndrome: Rheumatic heart disease of the fetus? Med. Hypotheses. 2006, 66, 554–565. [Google Scholar] [CrossRef]
- Guimarães-Camboa, N.; Stowe, J.; Aneas, I.; Sakabe, N.; Cattaneo, P.; Henderson, L.; Kilberg, M.S.; Johnson, R.S.; Chen, J.; McCulloch, A.D.; et al. HIF1α Represses Cell Stress Pathways to Allow Proliferation of Hypoxic Fetal Cardiomyocytes. Dev. Cell. 2015, 33, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase activity in human germline and embryonic tissues and cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Rhinn, M.; Ritschka, B.; Keyes, W.M. Cellular senescence in development, regeneration and disease. Development 2019, 146, dev151837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snodgrass, B.; Chilakala, A. Forgotten knowledge of the coronary-cameral connections and the rediscovery of their dynamic nature in development: The result of a misnomer, ambiguous use of nomenclature, and varied evaluation methods. Congenit. Heart Dis. 2017, 12, 647–648. [Google Scholar] [CrossRef]
- Wearn, J.T.; Mettier, S.R.; Klumpp, T.G.; Zschiesche, L.J. The nature of the vascular communications between the coronary arteries and the chambers of the heart. Am. Heart J. 1933, 9, 143–164. [Google Scholar] [CrossRef] [Green Version]
- Stephens, E.H.; Gupta, D.; Bleiweis, M.; Backer, C.L.; Anderson, R.H.; Spicer, D.E. Coronary Arterial abnormalities in Hypoplastic Left Heart Syndrome: Pathologic Characteristics of Archived Specimens. Semin. Thorac. Cardiovasc. Surg. 2020, 32, 531–538. [Google Scholar] [CrossRef]
- Abuchaim, D.C.; Tanamati, C.; Jatene, M.B.; Marcial, M.L.; Aiello, V.D. Coronary dominance patterns in hypoplastic left heart syndrome. Rev. Bras. Cir. Cardiovasc. 2011, 26, 604–608. [Google Scholar] [CrossRef] [Green Version]
- Baffa, J.M.; Chen, S.L.; Guttenberg, M.E.; Norwood, W.I.; Weinberg, P.M. Coronary artery abnormalities and right ventricular histology in hypoplastic left heart syndrome. J. Am. Coll. Cardiol. 1992, 20, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Nathan, M.; Williamson, A.K.; Mayer, J.E.; Bacha, E.A.; Juraszek, A.L. Mortality in hypoplastic left heart syndrome: Review of 216 autopsy cases of aortic atresia with attention to coronary artery disease. J. Thorac. Cardiovasc. Surg. 2012, 144, 1301–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.H.; Uebing, A.; Scheewe, J.; Kramer, H.H.; Fischer, G. Angiographic evaluation of the coronary artery anatomy in patients with hypoplastic left heart syndrome. Eur. J. Cardiothorac. Surg. 2012, 41, 561–568. [Google Scholar] [CrossRef]
- Bartram, U.; Grünenfelder, J.; Van Praagh, R. Causes of death after the modified Norwood procedure: A study of 122 postmortem cases. Ann. Thorac. Surg. 1997, 64, 1795–1802. [Google Scholar] [CrossRef]
- Gooty, V.D.; Veeram Reddy, S.R.; Greer, J.S.; Blair, Z.; Zahr, R.A.; Arar, Y.; Castellanos, D.A.; Pimplawar, S.; Greil, G.F.; Dillenbeck, J.; et al. Lymphatic pathway evaluation in congenital heart disease using 3D whole-heart balanced steady state free precession and T2-weighted cardiovascular magnetic resonance. J. Cardiovasc. Magn. Reson. 2021, 23, 16. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhry, B.; Alqahtani, A.; Eley, L.; Coats, L.; Moldovan, C.; Annavarapu, S.R.; Henderson, D.J. The Left Ventricular Myocardium in Hypoplastic Left Heart Syndrome. J. Cardiovasc. Dev. Dis. 2022, 9, 279. https://doi.org/10.3390/jcdd9080279
Chaudhry B, Alqahtani A, Eley L, Coats L, Moldovan C, Annavarapu SR, Henderson DJ. The Left Ventricular Myocardium in Hypoplastic Left Heart Syndrome. Journal of Cardiovascular Development and Disease. 2022; 9(8):279. https://doi.org/10.3390/jcdd9080279
Chicago/Turabian StyleChaudhry, Bill, Ahlam Alqahtani, Lorraine Eley, Louise Coats, Corina Moldovan, Srinivas R. Annavarapu, and Deborah J. Henderson. 2022. "The Left Ventricular Myocardium in Hypoplastic Left Heart Syndrome" Journal of Cardiovascular Development and Disease 9, no. 8: 279. https://doi.org/10.3390/jcdd9080279
APA StyleChaudhry, B., Alqahtani, A., Eley, L., Coats, L., Moldovan, C., Annavarapu, S. R., & Henderson, D. J. (2022). The Left Ventricular Myocardium in Hypoplastic Left Heart Syndrome. Journal of Cardiovascular Development and Disease, 9(8), 279. https://doi.org/10.3390/jcdd9080279