

First Report of Diplodia quercivora and Neofusicoccum vitifusiforme Associated with Cankers and Necrosis of Holm Oak (Quercus ilex) in Declining Stands in Southern Italy

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Visual Inspection of Trees, Sampling, and Isolation
2.2. Morphological Identification of the Fungal Isolates
2.3. Pathogenicity Tests
2.4. Molecular Characterization of the Isolates
2.5. Statistical Analysis
3. Results
3.1. Morphological Characterization of the Fungal Isolates
3.2. Pathogenicity Test
3.3. Molecular Identification of the Fungal Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Gladieux, P.; Gavrilets, S. Linking the emergence of fungal plant diseases with ecological speciation. Trends Ecol. Evol. 2010, 25, 387–395. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Brauman, K.A.; Cunniffe, N.J.; Fedoroff, N.V.; Finegold, C.; Garrett, K.A.; Gilligan, C.A.; Jones, C.M.; et al. The persistent threat of emerging plant disease pandemics to global food security. Proc. Natl. Acad. Sci. USA 2021, 118, e2022239118. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Ghelardini, L.; Pepori, A.L.; Luchi, N.; Capretti, P.; Santini, A. Drivers of emerging fungal diseases of forest trees. For. Ecol. Manag. 2016, 381, 235–246. [Google Scholar] [CrossRef]
- Brasier, C.M. Episodic selection as a force in fungal microevolution, with special reference to clonal speciation and hybrid introgression. Can. J. Bot. 1995, 73, 1213–1221. [Google Scholar] [CrossRef]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- Thynne, E.; McDonald, M.C.; Solomon, P.S. Phytopathogen emergence in the genomics era. Trends Plant Sci. 2015, 20, 246–255. [Google Scholar] [CrossRef]
- Cai, L.; Giraud, T.; Zhang, N.; Begerow, D.; Cai, G.; Shivas, R.G. The evolution of species concepts and species recognition criteria in plant pathogenic fungi. Fungal Divers. 2011, 50, 121–133. [Google Scholar] [CrossRef]
- Stergiopoulos, L.; Gordon, T.R. Cryptic fungal infections: The hidden agenda of plant pathogens. Front. Plant Sci. 2014, 5, 506. [Google Scholar] [CrossRef]
- Fournier, E.; Giraud, T. Sympatric genetic differentiation of a generalist pathogenic fungus, Botrytis cinerea, on two different host plants, grapevine and bramble. J. Evol. Biol. 2008, 21, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Refrégier, G.; Le Gac, M.; de Vienne, D.M.; Hood, M.E. Speciation in fungi. Fungal Genet. Biol. 2008, 45, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Delaye, L.; García-Guzmán, G.; Heil, M. Endophytes versus biotrophic and necrotrophic pathogens-are fungal lifestyles evolutionarily stable traits? Fungal Divers. 2013, 60, 125–135. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Loayza, P.; White, J.F.; Torres, M.S.; Balslev, H.; Kristiansen, T.; Svenning, J.C.; Gil, N. Light converts endosymbiotic fungus to pathogen, influencing seedling survival and niche-space filling of a common tropical tree, Iriartea deltoidea. PLoS ONE 2011, 6, e16386. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, D.; Ghelardini, L.; Tondini, E.; Luchi, N.; Santini, A. The potential of symptomless potted plants for carrying invasive soilborne plant pathogens. Divers. Distrib. 2015, 21, 1218–1229. [Google Scholar] [CrossRef]
- Andjic, V.; Dell, B.; Barber, P.; Hardy, G.; Wingfield, M.; Burgess, T. Plants for planting; indirect evidence for the movement of a serious forest pathogen, Teratosphaeria destructans, in Asia. Eur. J. Plant Pathol. 2011, 131, 49–58. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Kraus, C.; Markakis, E.; Alves, A.; Armengol, J.; Eichmeier, A.; Compant, S.; Gramaje, D. Fungal trunk diseases of fruit trees in Europe: Pathogens, spread and future directions. Phytopathol. Mediterr. 2023, 61, 563–599. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- De Wet, J.; Burgess, T.; Slippers, B.; Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Multiple gene genealogies and microsatellite markers reflect relationships between morphotypes of Sphaeropsis sapinea and distinguish a new species of Diplodia. Mycol. Res. 2003, 107, 557–566. [Google Scholar] [CrossRef]
- Alves, A.; Correia, A.; Luque, J.; Phillips, A. Botryosphaeria corticola, sp. nov. on Quercus species, with notes and description of Botryosphaeria stevensii and its anamorph, Diplodia mutila. Mycologia 2004, 96, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Crous, P.W.; Fourie, P.H. Botryosphaeriaceae as potential pathogens of Prunus species in South Africa, with descriptions of Diplodia africana and Lasiodiplodia plurivora sp. nov. Mycologia 2007, 99, 664–680. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR—Protocols and Applications—A Laboratory Manual; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A Method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Batista, E.; Lopes, A.; Alves, A. What do we know about Botryosphaeriaceae? An overview of a worldwide cured dataset. Forests 2021, 12, 313. [Google Scholar] [CrossRef]
- Morse, S.S. Factors and determinants of disease emergence. OIE Rev. Sci. Tech. 2004, 23, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera Island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Franceschini, A.; Alves, A.; Phillips, A.J.L. Diplodia quercivora sp. nov.: A new species of Diplodia found on declining Quercus canariensis trees in Tunisia. Mycologia 2013, 105, 1266–1274. [Google Scholar] [CrossRef]
- Ferreira, S.; Stauder, C.; Martin, D.; Kasson, M. Morphological and phylogenetic resolution of Diplodia corticola and D. quercivora, emerging canker pathogens of oak (Quercus spp.), in the United States. Plant Dis. 2021, 105, 1298–1307. [Google Scholar] [CrossRef]
- Bragança, H.; Neno, J.; Henriques, J.; Diogo, E.; Alves, A. First report of Diplodia quercivora causing dieback on Quercus suber and in Europe. Plant Dis. 2016, 100, 2166. [Google Scholar] [CrossRef]
- Mahamedi, A.E.; Phillips, A.J.L.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, distribution and host association of Botryosphaeriaceae species causing oak decline across different forest ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765. [Google Scholar] [CrossRef]
- Ragazzi, A.; Fedi, I.D.; Mesturino, L. The oak decline: A new problem in Italy. Eur. J. For. Pathol. 1989, 19, 105–110. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Gullino, M.L. Emerging and re-emerging fungus and oomycete soil-borne plant diseases in Italy. Phytopathol. Mediterr. 2019, 58, 451–472. [Google Scholar] [CrossRef]
- Frisullo, S.; Lima, G.; Magnano di San Lio, G.; Camele, I.; Melissano, L.; Puglisi, I.; Pane, A.; Agosteo, G.; Prudente, L.; Cacciola, S.O. Phytophthora cinnamomi involved in the decline of holm oak (Quercus ilex) stands in southern Italy. For. Sci. 2018, 64, 290–298. [Google Scholar] [CrossRef]
- Conte, A.L.; Di Pietro, R.; Iamonico, D.; Di Marzio, P.; Cillis, G.; Lucia, D.; Fortini, P. Oak decline in the Mediterranean basin: A study case from the southern Apennines (Italy). Plant Sociol. 2019, 56, 69–80. [Google Scholar] [CrossRef]
- Smahi, H.; Belhoucine-Guezouli, L.; Berraf-Tebbal, A.; Chouih, S.; Arkam, M.; Franceschini, A.; Linaldeddu, B.T.; Phillips, A.J.L. Molecular characterization and pathogenicity of Diplodia corticola and other Botryosphaeriaceae species associated with canker and dieback of Quercus suber in Algeria. Mycosphere 2017, 8, 1261–1272. [Google Scholar] [CrossRef]
- Mondello, V.; Lo Piccolo, S.; Conigliaro, G.; Alfonzo, A.; Torta, L.; Burruano, S. First report of Neofusiccoccum vitifusiforme and presence of other Botryosphaeriaceae species associated with Botryosphaeria dieback of grapevine in Sicily (Italy). Phytopathol. Mediterr. 2013, 52, 388–396. [Google Scholar]
- Muñoz-Adalia, E.J.; Uppara, A.B.; Albó, D.; Meijer, A.; Colinas, C. Cork harvest planning and climate: High air humidity favors availability of airborne inoculum of Diplodia corticola. For. Ecol. Manag. 2023, 536, 120935. [Google Scholar] [CrossRef]
- Pinna, C.; Linaldeddu, B.T.; Deiana, V.; Maddau, L.; Montecchio, L.; Lentini, A. Plant pathogenic fungi associated with Coraebus florentinus (Coleoptera: Buprestidae) attacks in declining oak forests. Forests 2019, 10, 488. [Google Scholar] [CrossRef]
- Muñoz-Adalia, E.J.; Meijer, A.; Colinas, C. New qPCR protocol to detect Diplodia corticola shows phoretic association with the oak pinhole borer Platypus cylindrus. Pest Manag. Sci. 2022, 78, 3534–3539. [Google Scholar] [CrossRef]
- Panzavolta, T.; Panichi, A.; Bracalini, M.; Croci, F.; Ginetti, B.; Ragazzi, A.; Tiberi, R.; Moricca, S. Dispersal and propagule pressure of Botryosphaeriaceae species in a declining oak stand is affected by insect vectors. Forests 2017, 8, 228. [Google Scholar] [CrossRef]
- Franceschini, A.; Linaldeddu, B.T.; Marras, F. Occurrence and distribution of fungal endophytes in declining cork oak forests in Sardinia (Italy). IOBC Wprs Bull. 2005, 28, 67. [Google Scholar]
- Kuntzmann, P.; Villaume, S.; Bertsch, C. Conidia dispersal of Diplodia species in a French vineyard. Phytopathol. Mediterr. 2009, 48, 150–154. [Google Scholar]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
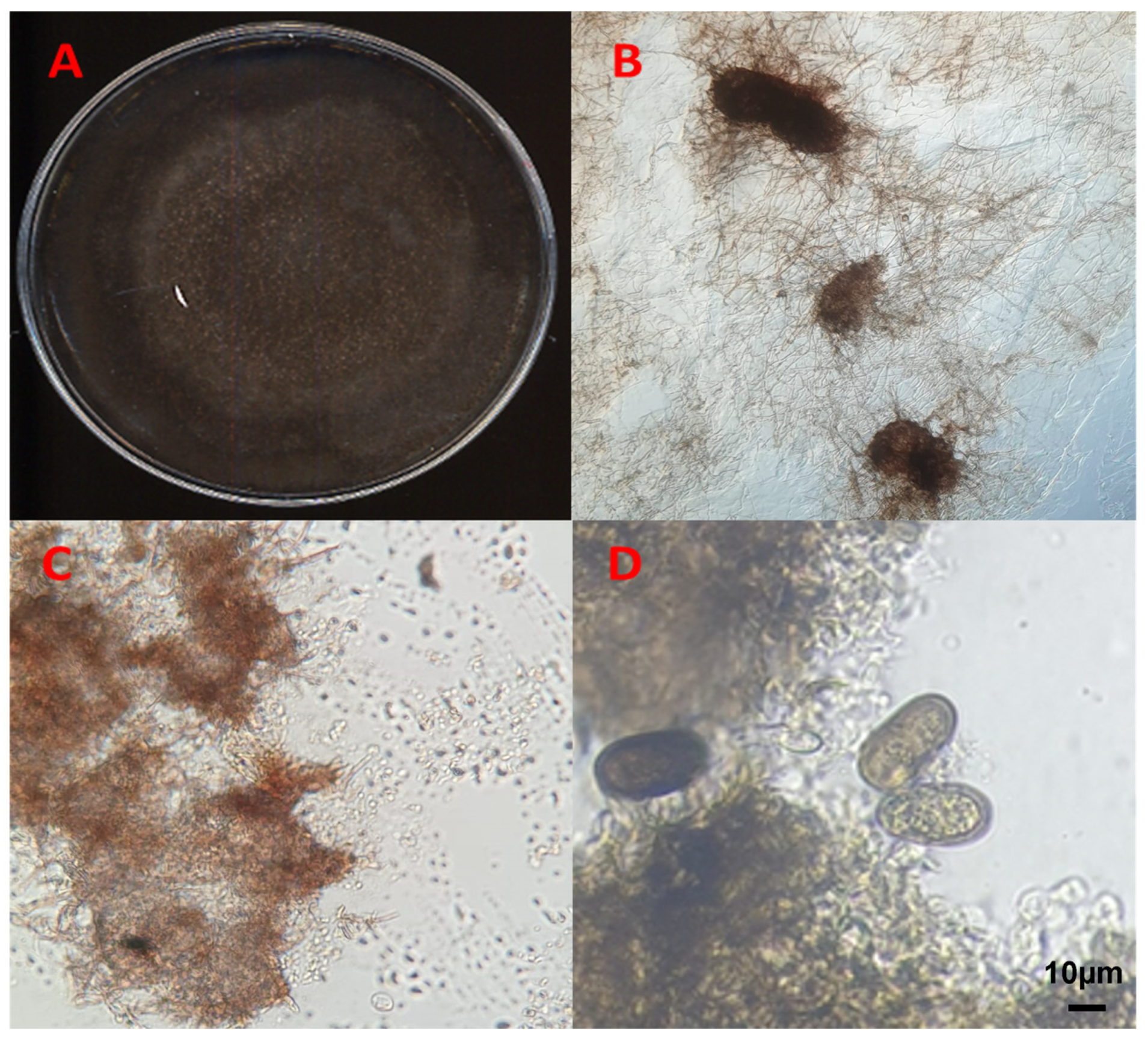{kind=link}
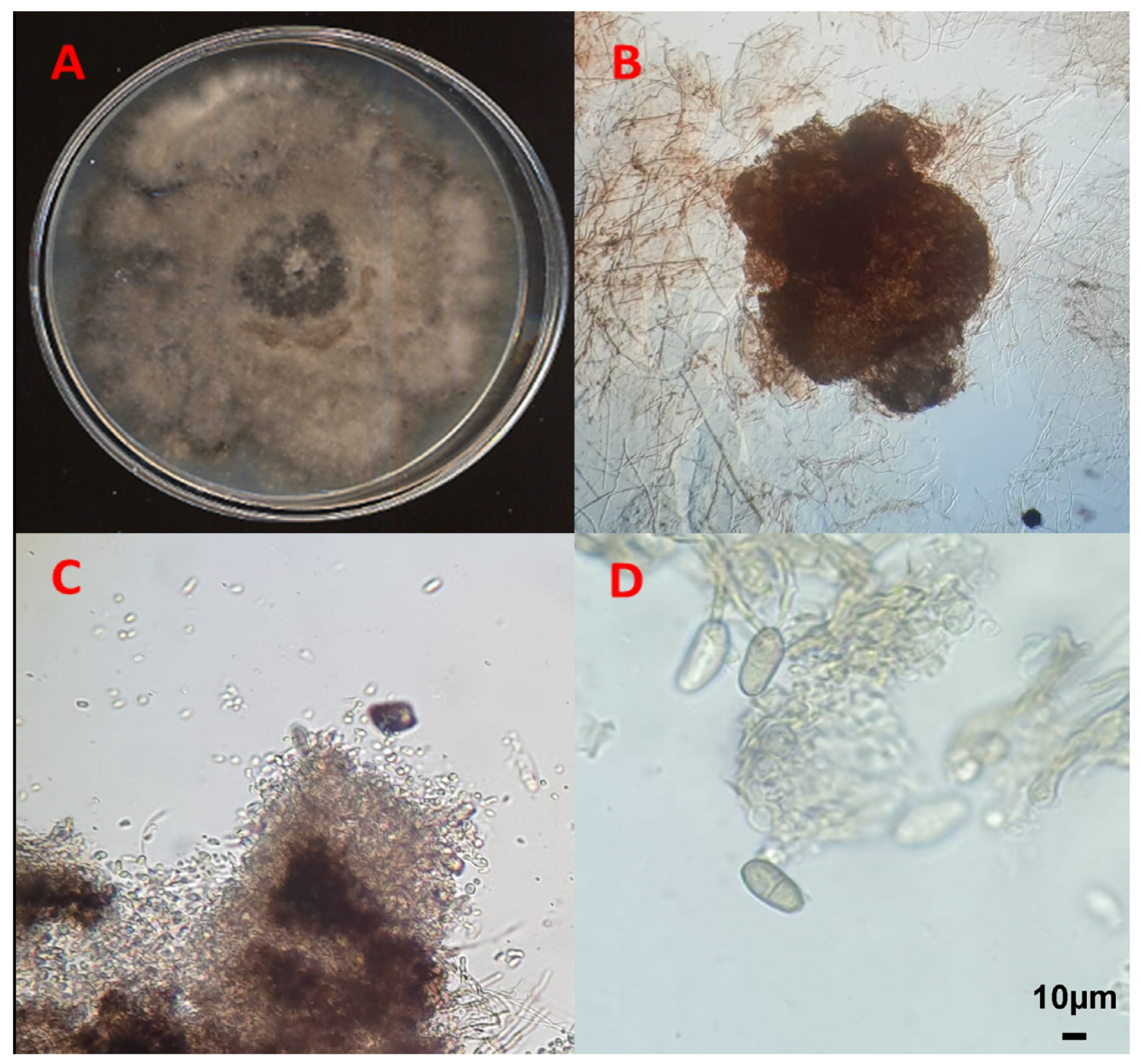{kind=link}
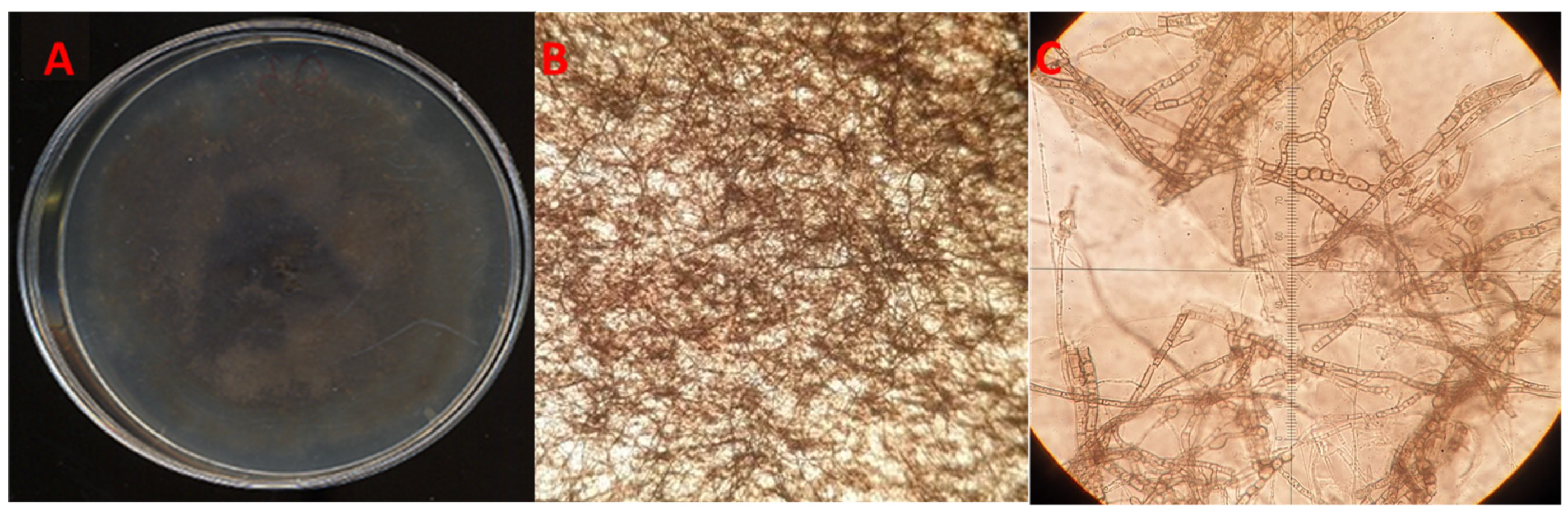{kind=link}
{kind=link}
{kind=link}
| Wood | Wood Name | Municipalities | Latitude (DD) | Longitude (DD) |
|---|---|---|---|---|
| 1 | Fraganite | Muro Leccese | 40.099761 | 18.313251 |
| 2 | Pozzo Mauro | Muro Leccese | 40.098289 | 18.346773 |
| 3 | Baia dei Turchi | Otranto | 40.191555 | 18.463726 |
| 4 | Maramonti | San Cassiano | 40.052636 | 18.315900 |
| 5 | Solicara | Lecce | 40.435958 | 18.201963 |
| 6 | Bosco Fiore | Lecce | 40.385044 | 18.255363 |
| 7 | Bosco San Biagio | Calimera | 40.253310 | 18.303520 |
| Accession n. (GenBank) | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Strain | Host | Country | Note | ITS | TEF-1 α |
| Neofusicoccum | vitifusiforme | QLE3 | Quercus ilex | Italy | Isolated in this study | OQ876772 | OQ750144 |
| Neofusicoccum | australe | CBS119046 | Rubus sp. | Portugal | / | DQ299244.1 | EU017541.1 |
| Neofusicoccum | australe | STE-U4425 | Vitis vinifera | South Africa | / | AY343388.1 | AY343347.1 |
| Neofusicoccum | luteum | CBS110299 | Vitis vinifera | Portugal | / | AY259091.1 | AY573217.1 |
| Neofusicoccum | luteum | CMW9076 | Malus x domestica | New Zealand | / | AY339257.1 | AY339265.1 |
| Neofusicoccum | vitifusiforme | STE-U5252 | Vitis vinifera | South Africa | / | AY343383.1 | AY343343.1 |
| Neofusicoccum | vitifusiforme | STE-U5050 | Vitis vinifera | South Africa | / | AY343382.1 | AY343344.1 |
| Neofusicoccum | viticlavatum | STE-U5044 | Vitis vinifera | South Africa | / | AY343381.1 | AY343342.1 |
| Neofusicoccum | viticlavatum | STE-U5041 | Vitis vinifera | South Africa | / | AY343380.1 | AY343341.1 |
| Neofusicoccum | parvum | CBS110301 | Vitis vinifera | Portugal | / | AY259098.2 | AY573221.1 |
| Neofusicoccum | parvum | CMW9081 | Populus nigra | New Zealand | / | AY236943.1 | AY236888.1 |
| Neofusicoccum | ribis | 96-187 | Ribes rubrum | Not known | / | AF241177.1 | AY236879.1 |
| Neofusicoccum | ribis | CMW7772 | Ribes sp. | USA | / | AY236935.1 | AY236877.1 |
| Neofusicoccum | mangiferae | CMW7024 | Mangifera indica | Australia | / | AY615185.1 | DQ093221.1 |
| Neofusicoccum | mangiferae | CMW7797 | Mangifera indica | Australia | / | AY615186.1 | DQ093220.1 |
| Neofusicoccum | eucalyptorum | CMW10125 | Eucalyptus grandis | South Africa | / | AF283686.1 | AY236891.1 |
| Neofusicoccum | eucalyptorum | CMW10126 | Eucalyptus grandis | South Africa | / | AF283687.1 | AY236892.1 |
| Neofusicoccum | eucalypticola | CMW6217 | Eucalyptus rossi | Australia | / | AY615143.1 | AY615135.1 |
| Neofusicoccum | eucalypticola | CMW6539 | Eucalyptus grandis | South Africa | / | AY615141.1 | AY615133.1 |
| Lasiodiplodia | Mediterranea | BL1 | Quercus ilex | Italy | Outgroup Genera | KJ638312.1 | KJ638331.1 |
| Diplodia | corticola | QLE1 | Quercus ilex | Italy | Isolated in this study | OQ876772 | OQ750142 |
| Diplodia | quercivora | QLE2 | Quercus ilex | Italy | Isolated in this study | OQ831537 | OQ750143 |
| Diplodia | corticola | CBS112072 | Quercus ilex | Spain | / | AY259108.1 | JX894221.1 |
| Diplodia | quercivora | BOT19 | Quercus suber | Algeria | / | MF535381.1 | MF535391.1 |
| Diplodia | corticola | BL10 | Quercus ilex | Italy | / | JX894191.1 | JX894210.1 |
| Diplodia | quercivora | BL8 | Quercus canariensis | Tunisia | / | JX894205.1 | JX894229.1 |
| Diplodia | scrobiculata | BL5 | Arbutus unedo | Italy | / | GU722102.1 | JX894231.1 |
| Diplodia | seriata | BL3 | Ulmus minor | Italy | / | JX894207.1 | JX894232.1 |
| Diplodia | mutila | CBS112553 | Vitis vinifera | Portugal | / | AY259093.2 | AY573219.1 |
| Diplodia | mutila | Sample301 | Juglans regia | Chile | / | MW412902.1 | MW574125.1 |
| Diplodia | sapinea | CMW39341 | Cedrus deodara | Montenegro | / | KF574998.1 | KF575028.1 |
| Diplodia | sapinea | CMW39338 | Cedrus atlantica | Serbia | / | KF574999.1 | KF575029.1 |
| Diplodia | seriata | CBS112555 | Vitis vinifera | Portugal | / | AY259094.2 | AY573220.1 |
| Diplodia | seriata | CBS119049 | Vitis sp. | Italy | / | DQ458889.1 | DQ458874.1 |
| Xylaria | hypoxylon | GB6391 | Isolated from soil | Mexico | Outgroup Order | AM993138.1 | AY327490.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Grosso, C.; Palmieri, D.; Marchese, L.; Melissano, L.; Lima, G. First Report of Diplodia quercivora and Neofusicoccum vitifusiforme Associated with Cankers and Necrosis of Holm Oak (Quercus ilex) in Declining Stands in Southern Italy. J. Fungi 2024, 10, 35. https://doi.org/10.3390/jof10010035
Del Grosso C, Palmieri D, Marchese L, Melissano L, Lima G. First Report of Diplodia quercivora and Neofusicoccum vitifusiforme Associated with Cankers and Necrosis of Holm Oak (Quercus ilex) in Declining Stands in Southern Italy. Journal of Fungi. 2024; 10(1):35. https://doi.org/10.3390/jof10010035
Chicago/Turabian StyleDel Grosso, Carmine, Davide Palmieri, Lucia Marchese, Luigi Melissano, and Giuseppe Lima. 2024. "First Report of Diplodia quercivora and Neofusicoccum vitifusiforme Associated with Cankers and Necrosis of Holm Oak (Quercus ilex) in Declining Stands in Southern Italy" Journal of Fungi 10, no. 1: 35. https://doi.org/10.3390/jof10010035
APA StyleDel Grosso, C., Palmieri, D., Marchese, L., Melissano, L., & Lima, G. (2024). First Report of Diplodia quercivora and Neofusicoccum vitifusiforme Associated with Cankers and Necrosis of Holm Oak (Quercus ilex) in Declining Stands in Southern Italy. Journal of Fungi, 10(1), 35. https://doi.org/10.3390/jof10010035