
Commercial Methods for Antifungal Susceptibility Testing of Saprophytic Molds: Can They Be Used to Detect Resistance?



Abstract
:1. Introduction
1.1. Development of Clinical Breakpoints and ECVs for Filamentous fungi
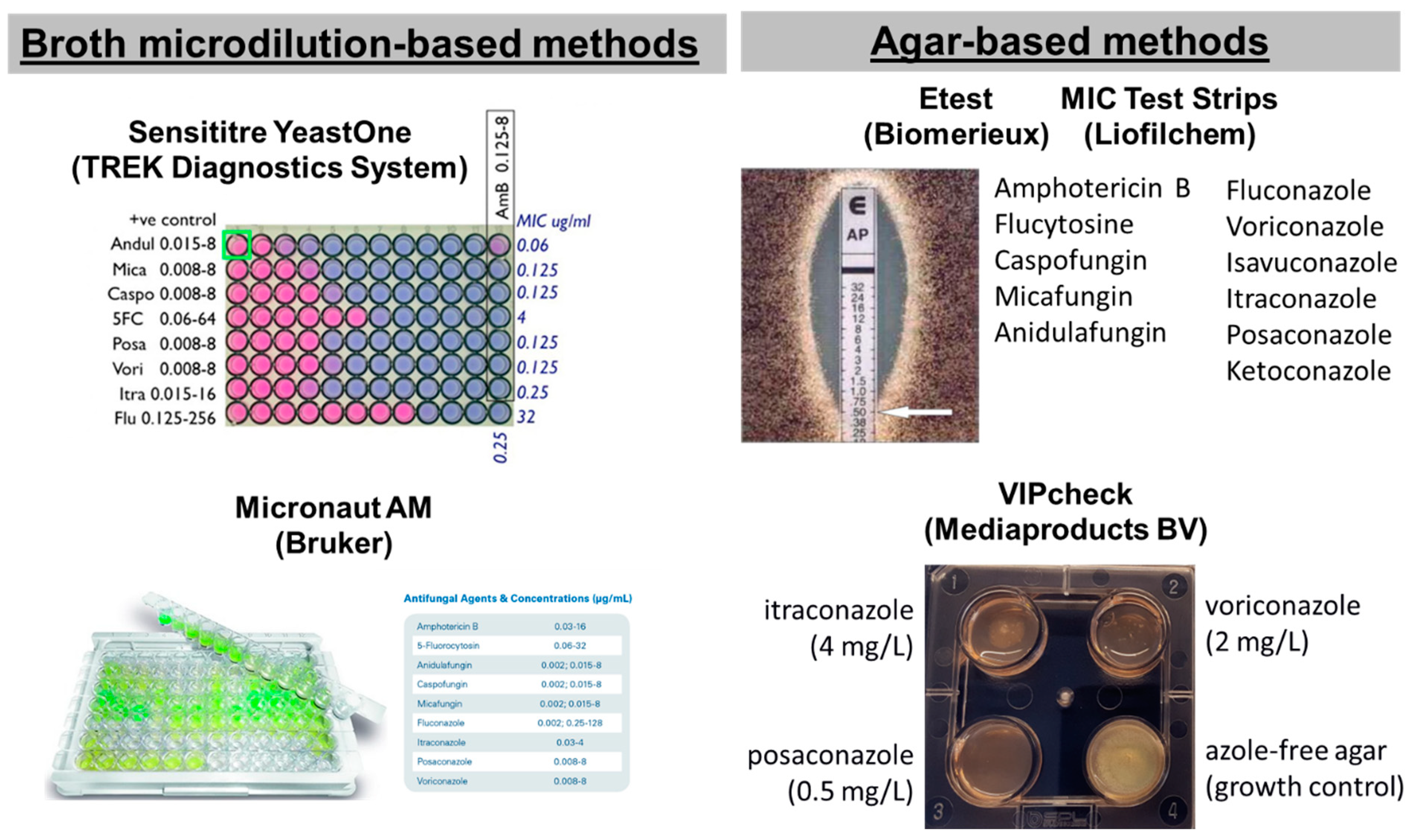
1.2. Commercial Methods for Antifungal Susceptibility Testing of Molds
2. Purpose of Review
2.1. Echinocandins
2.2. Triazoles
2.3. Polyenes
3. Gradient Concentration Strips
- Triazoles
- 2.
- Echinocandins
- 3.
- Polyenes
3.1. Sensititre YeastOne
- 1.
- Triazoles
- 2.
- Echinocandins
- 3.
- Polyenes
3.2. VIPcheck
3.3. Micronaut-AM
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and Management of Aspergillus Diseases: Executive Summary of the 2017 ESCMID-ECMM-ERS Guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi M38A, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; ISBN 978-1-56238-830-0. [Google Scholar]
- Guinea, J.; Meletiadis, J.; Arikan-Akdagli, S.; Muehlethaler, K.; Kahlmeter, G.; Arendrup, M. EUCAST Definitive Document E.Def 9.4. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Conidia Forming Moulds. 2022. Available online: https://eucast.org/astoffungi/ (accessed on 20 January 2024).
- Salsé, M.; Gangneux, J.P.; Cassaing, S.; Delhaes, L.; Fekkar, A.; Dupont, D.; Botterel, F.; Costa, D.; Bourgeois, N.; Bouteille, B.; et al. Multicentre Study to Determine the Etest Epidemiological Cut-off Values of Antifungal Drugs in Candida spp. and Aspergillus Fumigatus Species Complex. Clin. Microbiol. Infect. 2019, 25, 1546–1552. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Overview of Antifungal ECOFFs and Clinical Breakpoints for Yeasts, Moulds and Dermatophytes Using the EUCAST E.Def 7.3, E.Def 9.3 and E.Def 11.0 Procedures, Version 3.0. 2022. Available online: https://eucast.org/astoffungi/ (accessed on 20 January 2024).
- CLSI. M60-Ed2 June 2020 Replaces M60-Ed1 Performance Standards for Antifungal Susceptibility Testing of Yeasts, 2nd ed.; CLSI Supplement M60; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; ISBN 978-1-68440-083-6. [Google Scholar]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.G.; Meis, J.F. Azole Resistance in Aspergillus Fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Lionakis, M.S.; Lewis, R.E.; Chamilos, G.; Kontoyiannis, D.P. Aspergillus Susceptibility Testing in Patients with Cancer and Invasive Aspergillosis: Difficulties in Establishing Correlation between in Vitro Susceptibility Data and the Outcome of Initial Amphotericin B Therapy. Pharmacotherapy 2005, 25, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C.; Kofler, G.; Kropshofer, G.; Hermans, J.; Kreczy, A.; Dierich, M.P.; Niederwieser, D. In-Vitro Testing of Susceptibility to Amphotericin B Is a Reliable Predictor of Clinical Outcome in Invasive Aspergillosis. J. Antimicrob. Chemother. 1998, 42, 497–502. [Google Scholar] [CrossRef]
- Jiménez-Ortigosa, C.; Moore, C.; Denning, D.W.; Perlin, D.S. Emergence of Echinocandin Resistance Due to a Point Mutation in the Fks1 Gene of Aspergillus Fumigatus in a Patient with Chronic Pulmonary Aspergillosis. Antimicrob. Agents Chemother. 2017, 61, e01277-17. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Lewis, R.E.; Kontoyiannis, D.P. Role and Interpretation of Antifungal Susceptibility Testing for the Management of Invasive Fungal Infections. J. Fungi 2021, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Cuenca-Estrella, M.; Cantón, E. EUCAST and CLSI: Working Together Towards a Harmonized Method for Antifungal Susceptibility Testing. Curr. Fungal Infect. Rep. 2013, 7, 59–67. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing, Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 20 January 2024).
- Espinel-Ingroff, A.; Turnidge, J. The Role of Epidemiological Cutoff Values (ECVs/ECOFFs) in Antifungal Susceptibility Testing and Interpretation for Uncommon Yeasts and Moulds. Rev. Iberoam. Micol. 2016, 33, 63–75. [Google Scholar] [CrossRef]
- CLSI. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 4th ed.; CLSI Supplement M57S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; ISBN 978-1-68440-159-8. [Google Scholar]
- Espinel-Ingroff, A.; Alvarez-Fernandez, M.; Cantón, E.; Carver, P.L.; Chen, S.C.A.; Eschenauer, G.; Getsinger, D.L.; Gonzalez, G.M.; Govender, N.P.; Grancini, A.; et al. Multicenter Study of Epidemiological Cutoff Values and Detection of Resistance in Candida spp. to Anidulafungin, Caspofungin, and Micafungin Using the Sensititre YeastOne Colorimetric Method. Antimicrob. Agents Chemother. 2015, 59, 6725–6732. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Pfaller, M.A.; Bustamante, B.; Canton, E.; Fothergill, A.; Fuller, J.; Gonzalez, G.M.; Lass-Flörl, C.; Lockhart, S.R.; Martin-Mazuelos, E.; et al. Multilaboratory Study of Epidemiological Cutoff Values for Detection of Resistance in Eight Candida Species to Fluconazole, Posaconazole, and Voriconazole. Antimicrob. Agents Chemother. 2014, 58, 2006. [Google Scholar] [CrossRef]
- Turnidge, J.; Kahlmeter, G.; Kronvall, G. Statistical Characterisation of Bacterial Wild-Type MIC Value Distributions and the Determination of Epidemiological Cut-off Values. Clin. Microbiol. Infect. 2006, 12, 418–425. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Dannaoui, E. Should Etest MICs for Yeasts Be Categorized by Reference (BPs/ECVs) or by Etest (ECVs) Cutoffs as Determinants of Emerging Resistance? Curr. Fungal Infect. Rep. 2020, 14, 120–129. [Google Scholar] [CrossRef]
- Kahlmeter, G. The 2014 Garrod Lecture: EUCAST—Are We Heading towards International Agreement? J. Antimicrob. Chemother. 2015, 70, 2427–2439. [Google Scholar] [CrossRef]
- Dalhoff, A.; Ambrose, P.G.; Mouton, J.W. A Long Journey from Minimum Inhibitory Concentration Testing to Clinically Predictive Breakpoints: Deterministic and Probabilistic Approaches in Deriving Breakpoints. Infection 2009, 37, 296–305. [Google Scholar] [CrossRef]
- Turnidge, J.; Paterson, D.L. Setting and Revising Antibacterial Susceptibility Breakpoints. Clin. Microbiol. Rev. 2007, 20, 391–408. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Ghannoum, M.A.; Alexander, B.D. Establishment and Use of Epidemiological Cutoff Values for Molds and Yeasts by Use of the Clinical and Laboratory Standards Institute M57 Standard. J. Clin. Microbiol. 2017, 55, 1262. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Colombo, A.L.; Cordoba, S.; Dufresne, P.J.; Fuller, J.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J.; Kidd, S.E.; Meis, J.F.; et al. International Evaluation of MIC Distributions and Epidemiological Cutoff Value (ECV) Definitions for Fusarium Species Identified by Molecular Methods for the CLSI Broth Microdilution Method. Antimicrob. Agents Chemother. 2016, 60, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Arendrup, M.; Canton, E.; Cordob, S.; Dannaoui, E.; Garcia-Rodriguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter Study of Method-Dependent Epidemiological Cutoff Values for Detection of Resistance in Candida spp. and Aspergillus spp. to Amphotericin B and Echinocandins for the Etest Agar Diffusion Method. Antimicrob. Agents Chemother. 2016, 61, 1792–1808. [Google Scholar] [CrossRef] [PubMed]
- Dannaoui, E.; Espinel-Ingroff, A. Antifungal Susceptibly Testing by Concentration Gradient Strip Etest Method for Fungal Isolates: A Review. J. Fungi 2019, 5, 108. [Google Scholar] [CrossRef]
- CLSI. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 3rd ed.; CLSI Supplement M59; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing. Data from the EUCAST MIC Distribution Website. Available online: https://eucast.org/astoffungi/ (accessed on 20 January 2024).
- Espinel-Ingroff, A.; Turnidge, J.; Alastruey-Izquierdo, A.; Botterel, F.; Canton, E.; Castro, C.; Chen, Y.C.; Chen, Y.; Chryssanthou, E.; Dannaoui, E.; et al. Method-Dependent Epidemiological Cutoff Values for Detection of Triazole Resistance in Candida and Aspergillus Species for the Sensititre Yeastone Colorimetric Broth and Etest Agar Diffusion Methods. Antimicrob. Agents Chemother. 2019, 63, e01651-18. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Turnidge, J.; Alastruey-Izquierdo, A.; Dannaoui, E.; Garcia-Effron, G.; Guinea, J.; Kidd, S.; Pelaez, T.; Sanguinetti, M.; Meletiadis, J.; et al. Posaconazole MIC Distributions for Aspergillus Fumigatus Species Complex by Four Methods: Impact of Cyp51a Mutations on Estimation of Epidemiological Cutoff Values. Antimicrob. Agents Chemother. 2018, 62, e01916-17. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Sasso, M.; Turnidge, J.; Arendrup, M.; Botterel, F.; Bourgeois, N.; Bouteille, B.; Canton, E.; Cassaing, S.; Dannaoui, E.; et al. Etest ECVs/ECOFFs for Detection of Resistance in Prevalent and Three Nonprevalent Candida spp. to Triazoles and Amphotericin B and Aspergillus spp. to Caspofungin: Further Assessment of Modal Variability. Antimicrob. Agents Chemother. 2021, 65, e0109321. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. MIC Distributions and Epidemiological Cut-Off Value (ECOFF) Setting, EUCAST SOP 10.2. 2021. Available online: https://eucast.org/astoffungi/ (accessed on 20 January 2024).
- Borman, M.; Espinel-Ingroff, A. Commercial Methods for Antifungal Susceptibility Testing of Yeasts: Strengths and Limitations as Predictors of Resistance. J. Fungi 2022, 8, 309. [Google Scholar]
- Berkow, E.L.; Lockhart, S.R.; Ostrosky-Zeichner, L. Antifungal Susceptibility Testing: Current Approaches. Clin. Microbiol. Rev. 2020, 33, e00069-19. [Google Scholar] [CrossRef]
- Arastehfar, A.; Carvalho, A.; Houbraken, J.; Lombardi, L.; Garcia-Rubio, R.; Jenks, J.D.; Rivero-Menendez, O.; Aljohani, R.; Jacobsen, I.D.; Berman, J.; et al. Aspergillus Fumigatus and Aspergillosis: From Basics to Clinics. Stud. Mycol. 2021, 100, 100115. [Google Scholar] [CrossRef] [PubMed]
- Siopi, M.; Pournaras, S.; Meletiadis, J. Comparative Evaluation of Sensititre YeastOne and CLSI M38-A2 Reference Method for Antifungal Susceptibility Testing of Aspergillus spp. against Echinocandins. J. Clin. Microbiol. 2017, 55, 1714–1719. [Google Scholar] [CrossRef]
- Mello, E.; Posteraro, B.; Vella, A.; De Carolis, E.; Torelli, R.; D’Inzeo, T.; Verweij, P.E.; Sanguinetti, M. Susceptibility Testing of Common and Uncommon Aspergillus Species against Posaconazole and Other Mold-Active Antifungal Azoles Using the Sensititre Method. Antimicrob. Agents Chemother. 2017, 61, e00168-17. [Google Scholar] [CrossRef]
- Martín-Mazuelos, E.; Pemán, J.; Valverde, A.; Chaves, M.; Serrano, M.C.; Cantón, E. Comparison of the Sensititre YeastOne Colorimetric Antifungal Panel and Etest with the NCCLS M38-A Method to Determine the Activity of Amphotericin B and Itraconazole against Clinical Isolates of Aspergillus spp. J. Antimicrob. Chemother. 2003, 52, 365–370. [Google Scholar] [CrossRef]
- Torres-Narbona, M.; Guinea, J.; Martínez-Alarcón, J.; Peláez, T.; Bouza, E. In Vitro Activities of Amphotericin B, Caspofungin, Itraconazole, Posaconazole, and Voriconazole against 45 Clinical Isolates of Zygomycetes: Comparison of CLSI M38-A, Sensititre YeastOne, and the Etest. Antimicrob. Agents Chemother. 2007, 51, 1126. [Google Scholar] [CrossRef]
- Lamoth, F.; Alexander, B.D. Comparing Etest and Broth Microdilution for Antifungal Susceptibility Testing of the Most-Relevant Pathogenic Molds. J. Clin. Microbiol. 2015, 53, 3176–3181. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Lago, M.; Branco, L.; Vale-Silva, L.A.; Pinheiro, M.D. Evaluation of Etest Performed in Mueller-Hinton Agar Supplemented with Glucose for Antifungal Susceptibility Testing of Clinical Isolates of Filamentous Fungi. Mycopathologia 2014, 177, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.C.; Morilla, D.; Valverde, A.; Chávez, M.; Espinel-Ingroff, A.; Claro, R.; Ramírez, M.; Martín Mazuelos, E. Comparison of Etest with Modified Broth Microdilution Method for Testing Susceptibility of Aspergillus spp. to Voriconazole. J. Clin. Microbiol. 2003, 41, 5270. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Boyken, L.; Hollis, R.J.; Diekema, D.J. In Vitro Susceptibility Testing of Filamentous Fungi: Comparison of Etest and Reference M38-A Microdilution Methods for Determining Posaconazole MICs. Diagn. Microbiol. Infect. Dis. 2003, 45, 241–244. [Google Scholar] [CrossRef]
- Martos, A.I.; Romero, A.; González, M.T.; González, A.; Serrano, C.; Castro, C.; Pemán, J.; Cantón, E.; Martn-Mazuelos, E. Evaluation of the Etest Method for Susceptibility Testing of Aspergillus spp. and Fusarium spp. to Three Echinocandins. Med. Mycol. 2010, 48, 858–861. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Verweij, P.; Nielsen, H.V. Evaluation of MIC Strip Isavuconazole Test for Susceptibility Testing of Wild-Type and Non-Wild-Type Aspergillus Fumigatus Isolates. Antimicrob. Agents Chemother. 2017, 61, e01659-16. [Google Scholar] [CrossRef] [PubMed]
- Ozkutuk, A.; Ergon, C.; Metin, D.Y.; Yucesoy, M.; Polat, S.H. Comparison of Disk Diffusion, E-Test and Broth Microdilution Test in Determination of Susceptibility of Aspergillus Species to Amphotericin B, Itraconazole and Voriconazole. J. Chemother. 2008, 20, 87–92. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Normand, A.C.; Ranque, S.; Piarroux, R.; De Hoog, G.S.; Meletiadis, J.; Meis, J.F. Comparative Evaluation of Etest, EUCAST, and CLSI Methods for Amphotericin B, Voriconazole, and Posaconazole against Clinically Relevant Fusarium Species. Antimicrob. Agents Chemother. 2016, 61, e01671-16. [Google Scholar] [CrossRef]
- Nuh, A.; Ramadan, N.; Schelenz, S.; Armstrong-James, D. Comparative Evaluation of MIRONAUT-AM and CLSI Broth Microdilution Method for Antifungal Susceptibility Testing of Aspergillus Species against Four Commonly Used Antifungals. Med. Mycol. 2020, 58, 1085–1090. [Google Scholar] [CrossRef]
- Gyurtane Szabo, N.; Joste, V.; Houzé, S.; Dannaoui, E.; Bonnal, C. Comparison of the Micronaut-AM System and the EUCAST Broth Microdilution Reference Method for MIC Determination of Four Antifungals against Aspergillus Fumigatus. J. Fungi 2023, 9, 721. [Google Scholar] [CrossRef]
- Tsitsopoulou, A.; Posso, R.; Vale, L.; Bebb, S.; Johnson, E.; White, P.L. Determination of the Prevalence of Triazole Resistance in Environmental Aspergillus Fumigatus Strains Isolated in South Wales, UK. Front. Microbiol. 2018, 9, 386222. [Google Scholar] [CrossRef]
- Serrano-Lobo, J.; Gómez, A.; Rodríguez-Sánchez, B.; Muñoz, P.; Escribano, P.; Guinea, J. Azole-Resistant Aspergillus Fumigatus Clinical Isolate Screening in Azole-Containing Agar Plates (EUCAST E.Def 10.1): Low Impact of Plastic Trays Used and Poor Performance in Cryptic Species. Antimicrob. Agents Chemother. 2021, 65, e0048221. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Principles and Procedures for the Development of Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 1st ed.; CLSI Document M57; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Arendrup, M.C.; Garcia-Effron, G.; Lass-Flörl, C.; Lopez, A.G.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Perlin, D.S. Echinocandin Susceptibility Testing of Candida Species: Comparison of EUCAST EDef 7.1, CLSI M27-A3, Etest, Disk Diffusion, and Agar Dilution Methods with RPMI and IsoSensitest Media. Antimicrob. Agents Chemother. 2010, 54, 426. [Google Scholar] [CrossRef] [PubMed]
- Imhof, A.; Balajee, S.A.; Marr, K.A. New Methods to Assess Susceptibilities of Aspergillus Isolates to Caspofungin. J. Clin. Microbiol. 2003, 41, 5683–5688. [Google Scholar] [CrossRef]
- e Silva, A.P.; Miranda, I.M.; Branco, J.; Oliveira, P.; Faria-Ramos, I.; Silva, R.M.; Rodrigues, A.G.; Costa-de-Oliveira, S. FKS1 Mutation Associated with Decreased Echinocandin Susceptibility of Aspergillus Fumigatus Following Anidulafungin Exposure. Sci. Rep. 2020, 10, 1–6. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Garcia-Effron, G.; Buzina, W.; Mortensen, K.L.; Reiter, N.; Lundin, C.; Jensen, H.E.; Lass-Florl, C.; Perlin, D.S.; Bruun, B. Breakthrough Aspergillus Fumigatus and Candida Albicans Double Infection during Caspofungin Treatment: Laboratory Characteristics and Implication for Susceptibility Testing. Antimicrob. Agents Chemother. 2009, 53, 1185–1193. [Google Scholar] [CrossRef]
- Gardiner, R.E.; Souteropoulos, P.; Park, S.; Perlin, D.S. Characterization of Aspergillus Fumigatus Mutants with Reduced Susceptibility to Caspofungin. Med. Mycol. 2005, 43, S299–S305. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.M.F.; Garcia-Effron, G.; Park, S.; Perlin, D.S. A Ser678Pro Substitution in Fks1p Confers Resistance to Echinocandin Drugs in Aspergillus Fumigatus. Antimicrob. Agents Chemother. 2007, 51, 4174. [Google Scholar] [CrossRef]
- Satish, S.; Jiménez-Ortigosa, C.; Zhao, Y.; Lee, M.H.; Dolgov, E.; Krüger, T.; Park, S.; Denning, D.W.; Kniemeyer, O.; Brakhage, A.A.; et al. Stress-Induced Changes in the Lipid Microenvironment of β-(1,3)-d-Glucan Synthase Cause Clinically Important Echinocandin Resistance in Aspergillus Fumigatus. mBio 2019, 10, e00779-19. [Google Scholar] [CrossRef]
- Antachopoulos, C.; Meletiadis, J.; Sein, T.; Roilides, E.; Walsh, T.J. Concentration-Dependent Effects of Caspofungin on the Metabolic Activity of Aspergillus Species. Antimicrob. Agents Chemother. 2007, 51, 881–887. [Google Scholar] [CrossRef]
- Jimeńez-Ortigosa, C.; Aimanianda, V.; Muszkieta, L.; Mouyna, I.; Alsteens, D.; Pire, S.; Beau, R.; Krappmann, S.; Beauvais, A.; Dufrêne, Y.F.; et al. Chitin Synthases with a Myosin Motor-Like Domain Control the Resistance of Aspergillus Fumigatus to Echinocandins. Antimicrob. Agents Chemother. 2012, 56, 6121. [Google Scholar] [CrossRef] [PubMed]
- Fortwendel, J.R.; Juvvadi, P.R.; Perfect, B.Z.; Rogg, L.E.; Perfect, J.R.; Steinbach, W.J. Transcriptional Regulation of Chitin Synthases by Calcineurin Controls Paradoxical Growth of Aspergillus Fumigatus in Response to Caspofungin. Antimicrob. Agents Chemother. 2010, 54, 1555. [Google Scholar] [CrossRef] [PubMed]
- Ancuceanu, R.; Hovaneț, M.V.; Cojocaru-Toma, M.; Anghel, A.I.; Dinu, M. Potential Antifungal Targets for Aspergillus sp. from the Calcineurin and Heat Shock Protein Pathways. Int. J. Mol. Sci. 2022, 23, 12543. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Chowdhary, A. Molecular Bases of Antifungal Resistance in Filamentous Fungi. Int. J. Antimicrob. Agents 2017, 50, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Juvvadi, P.R.; Gehrke, C.; Asfaw, Y.G.; Steinbach, W.J. Transcriptional Activation of Heat Shock Protein 90 Mediated via a Proximal Promoter Region as Trigger of Caspofungin Resistance in Aspergillus Fumigatus. J. Infect. Dis. 2014, 209, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Satish, S.; Perlin, D.S. Echinocandin Resistance in Aspergillus Fumigatus Has Broad Implications for Membrane Lipid Perturbations That Influence Drug-Target Interactions. Microbiol. Insights 2019, 12, 117863611989703. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole Resistance in Aspergillus Species: An Emerging Problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Dabas, Y.; Xess, I.; Bakshi, S.; Mahapatra, M.; Seth, R. Emergence of Azole-Resistant Aspergillus Fumigatus from Immunocompromised Hosts in India. Antimicrob. Agents Chemother. 2018, 62, e02264-17. [Google Scholar] [CrossRef]
- Berkow, E.L.; Nunnally, N.S.; Bandea, A.; Kuykendall, R.; Beer, K.; Lockhart, S.R. Detection of TR34/L98H CYP51A Mutation through Passive Surveillance for Azole-Resistant Aspergillus Fumigatus in the United States from 2015 to 2017. Antimicrob Agents Chemother 2018, 62, e02240-17. [Google Scholar] [CrossRef]
- Wiederhold, N.P.; Gil, V.G.; Gutierrez, F.; Lindner, J.R.; Albataineh, M.T.; McCarthy, D.I.; Sanders, C.; Fan, H.; Fothergill, A.W.; Sutton, D.A. First Detection of TR34 L98H and TR46 Y121F T289A Cyp51 Mutations in Aspergillus Fumigatus Isolates in the United States. J. Clin. Microbiol. 2016, 54, 168–171. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Verweij, P.E.; Mouton, J.W.; Lagrou, K.; Meletiadis, J. Multicentre Validation of 4-Well Azole Agar Plates as a Screening Method for Detection of Clinically Relevant Azole-Resistant Aspergillus Fumigatus. J. Antimicrob. Chemother. 2017, 72, 3325–3333. [Google Scholar] [CrossRef]
- Gonzalez-Jimenez, I.; Lucio, J.; Amich, J.; Cuesta, I.; Arroyo, R.S.; Alcazar-Fuoli, L.; Mellado, E. A Cyp51B Mutation Contributes to Azole Resistance in Aspergillus Fumigatus. J. Fungi 2020, 6, 315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The Fungal CYP51s: Their Functions, Structures, Related Drug Resistance, and Inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef]
- Buied, A.; Moore, C.B.; Denning, D.W.; Bowyer, P. High-Level Expression of Cyp51B in Azole-Resistant Clinical Aspergillus Fumigatus Isolates. J. Antimicrob. Chemother. 2013, 68, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Handelman, M.; Meir, Z.; Scott, J.; Shadkchan, Y.; Liu, W.; Ben-Ami, R.; Amich, J.; Osherov, N. Point Mutation or Overexpression of Aspergillus Fumigatus, Encoding Lanosterol 14α-Sterol Demethylase, Leads to Triazole Resistance. Antimicrob. Agents Chemother. 2021, 65, AAC0125221. [Google Scholar] [CrossRef]
- Sharma, C.; Kumar, R.; Kumar, N.; Masih, A.; Gupta, D.; Chowdhary, A. Investigation of Multiple Resistance Mechanisms in Voriconazole-Resistant Aspergillus Flavus Clinical Isolates from a Chest Hospital Surveillance in Delhi, India. Antimicrob. Agents Chemother. 2018, 62, e01928-17. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sun, Y.; Chen, W.; Liu, W.; Wan, Z.; Bu, D.; Li, R. The T788G Mutation in the Cyp51C Gene Confers Voriconazole Resistance in Aspergillus Flavus Causing Aspergillosis. Antimicrob. Agents Chemother. 2012, 56, 2598. [Google Scholar] [CrossRef]
- Lucio, J.; Gonzalez-Jimenez, I.; Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Pelaez, T.; Alcazar-Fuoli, L.; Mellado, E. Point Mutations in the 14-α Sterol Demethylase Cyp51A or Cyp51C Could Contribute to Azole Resistance in Aspergillus Flavus. Genes 2020, 11, 1217. [Google Scholar] [CrossRef]
- Chen, M.; Zhong, G.; Wang, S.; Chen, P.; Li, L. Deletion of Cox7c Results in Pan-Azole Resistance in Aspergillus Fumigatus. Antimicrob. Agents Chemother. 2022, 66, e0015122. [Google Scholar] [CrossRef]
- Tobin, M.B.; Peery, R.B.; Skatrud, P.L. Genes Encoding Multiple Drug Resistance-like Proteins in Aspergillus Fumigatus and Aspergillus Flavus. Gene 1997, 200, 11–23. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Hagen, F.; Meis, J.F. Exploring Azole Antifungal Drug Resistance in Aspergillus Fumigatus with Special Reference to Resistance Mechanisms. Future Med. 2014, 9, 697–711. [Google Scholar] [CrossRef] [PubMed]
- da Silva Ferreira, M.E.; Malavazi, I.; Savoldi, M.; Brakhage, A.A.; Goldman, M.H.S.; Kim, H.S.; Nierman, W.C.; Goldman, G.H. Transcriptome Analysis of Aspergillus Fumigatus Exposed to Voriconazole. Curr. Genet. 2006, 50, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The Cdr1B Efflux Transporter Is Associated with Non-Cyp51a-Mediated Itraconazole Resistance in Aspergillus Fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Camps, S.M.T.; Dutilh, B.E.; Arendrup, M.C.; Rijs, A.J.M.M.; Snelders, E.; Huynen, M.A.; Verweij, P.E.; Melchers, W.J.G. Discovery of a HapE Mutation That Causes Azole Resistance in Aspergillus Fumigatus through Whole Genome Sequencing and Sexual Crossing. PLoS ONE 2012, 7, e50034. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Chen, P.; Gao, R.; Li, Y.; Zhang, A.; Liu, F.; Lu, L. Screening and Characterization of a Non-Cyp51A Mutation in an Aspergillus Fumigatus Cox10 Strain Conferring Azole Resistance. Antimicrob. Agents Chemother. 2017, 61, e02101-16. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.R.H.; Bicanic, T.; Salim, R.; Hope, W. Liposomal Amphotericin B (AmBisome®): A Review of the Pharmacokinetics, Pharmacodynamics, Clinical Experience and Future Directions. Drugs 2016, 76, 485. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Dietl, A.M.; Kontoyiannis, D.P.; Brock, M. Aspergillus Terreus Species Complex. Clin. Microbiol. Rev. 2021, 34, e0031120. [Google Scholar] [CrossRef]
- Blum, G.; Hörtnagl, C.; Jukic, E.; Erbeznik, T.; Pümpel, T.; Dietrich, H.; Nagl, M.; Speth, C.; Rambach, G.; Lass-Flörl, C. New Insight into Amphotericin B Resistance in Aspergillus Terreus. Antimicrob. Agents Chemother. 2013, 57, 1583. [Google Scholar] [CrossRef]
- Fakhim, H.; Badali, H.; Dannaoui, E.; Nasirian, M.; Jahangiri, F.; Raei, M.; Vaseghi, N.; Ahmadikia, K.; Vaezi, A. Trends in the Prevalence of Amphotericin B-Resistance (AmBR) among Clinical Isolates of Aspergillus Species. J. Mycol. Med. 2022, 32, 101310. [Google Scholar] [CrossRef]
- Walsh, T.J.; Petraitis, V.; Petraitiene, R.; Field-Ridley, A.; Sutton, D.; Ghannoum, M.; Sein, T.; Schaufele, R.; Peter, J.; Bacher, J.; et al. Experimental Pulmonary Aspergillosis Due to Aspergillus Terreus: Pathogenesis and Treatment of an Emerging Fungal Pathogen Resistant to Amphotericin B. J. Infect. Dis. 2003, 188, 305–319. [Google Scholar] [CrossRef]
- Fan, Y.; Korfanty, G.A.; Xu, J. Genetic Analyses of Amphotericin b Susceptibility in Aspergillus Fumigatus. J. Fungi 2021, 7, 860. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Shi, G.H.; Dai, Y.; Fang, W.X.; Wu, Q. Identifying Genetic Variants Associated with Amphotericin B (AMB) Resistance in Aspergillus Fumigatus via k-Mer—Based GWAS. Front. Genet. 2023, 14, 1133593. [Google Scholar] [CrossRef]
- Hadrich, I.; Makni, F.; Neji, S.; Cheikhrouhou, F.; Bellaaj, H.; Elloumi, M.; Ayadi, A.; Ranque, S. Amphotericin B in Vitro Resistance Is Associated with Fatal Aspergillus Flavus Infection. Med. Mycol. 2012, 50, 829–834. [Google Scholar] [CrossRef]
- Seo, K.; Akiyoshi, H.; Ohnishi, Y. Alteration of Cell Wall Composition Leads to Amphotericin B Resistance in Aspergillus Flavus. Microbiol. Immunol. 1999, 43, 1017–1025. [Google Scholar] [CrossRef]
- Chowdhary, A.; Masih, A.; Sharma, C. Azole Resistance in Moulds—Approach to Detection in a Clinical Laboratory. Curr. Fungal Infect. Rep. 2016, 10, 96–106. [Google Scholar] [CrossRef]
- Kathuria, S.; Sharma, C.; Singh, P.K.; Agarwal, P.; Agarwal, K.; Hagen, F.; Meis, J.F.; Chowdhary, A. Molecular Epidemiology and In-Vitro Antifungal Susceptibility of Aspergillus Terreus Species Complex Isolates in Delhi, India: Evidence of Genetic Diversity by Amplified Fragment Length Polymorphism and Microsatellite Typing. PLoS ONE 2015, 10, e0118997. [Google Scholar] [CrossRef]
- Jukic, E.; Blatzer, M.; Posch, W.; Steger, M.; Binder, U.; Lass-Flörl, C.; Wilflingseder, D. Oxidative Stress Response Tips the Balance in Aspergillus Terreus Amphotericin B Resistance. Antimicrob. Agents Chemother. 2017, 61, e00670-17. [Google Scholar] [CrossRef]
- Blum, G.; Perkhofer, S.; Haas, H.; Schrettl, M.; Würzner, R.; Dierich, M.P.; Lass-Flörl, C. Potential Basis for Amphotericin B Resistance in Aspergillus Terreus. Antimicrob. Agents Chemother. 2008, 52, 1553–1555. [Google Scholar] [CrossRef]
- Burgel, P.R.; Baixench, M.T.; Amsellem, M.; Audureau, E.; Chapron, J.; Kanaan, R.; Honoré, I.; Dupouy-Camet, J.; Dusser, D.; Klaassen, C.H.; et al. High Prevalence of Azole-Resistant Aspergillus Fumigatus in Adults with Cystic Fibrosis Exposed to Itraconazole. Antimicrob. Agents Chemother. 2012, 56, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Kano, R.; Sobukawa, H.; Murayama, S.Y.; Hirose, D.; Tanaka, Y.; Kosuge, Y.; Hasegawa, A.; Kamata, H. In Vitro Resistance of Aspergillus Fumigatus to Azole Farm Fungicide. J. Infect. Chemother. 2016, 22, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Siopi, M.; Perlin, D.S.; Arendrup, M.C.; Pournaras, S.; Meletiadis, J. Comparative Pharmacodynamics of Echinocandins against Aspergillus Fumigatus Using an In Vitro Pharmacokinetic/Pharmacodynamic Model That Correlates with Clinical Response to Caspofungin Therapy: Is There a Place for Dose Optimization? Antimicrob. Agents Chemother. 2021, 65, e01618-20. [Google Scholar] [CrossRef] [PubMed]
- Aruanno, M.; Glampedakis, E.; Lamoth, F. Echinocandins for the Treatment of Invasive Aspergillosis: From Laboratory to Bedside. Antimicrob. Agents Chemother. 2019, 63, e00399-1. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C.; Arendrup, M.C.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Donnelly, P.; Hope, W. EUCAST Technical Note on Amphotericin B. Clin. Microbiol. Infect. 2011, 17, E27–E29. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Cuenca-Estrella, M.; Fothergill, A.; Fuller, J.; Ghannoum, M.; Johnson, E.; Pelaez, T.; Pfaller, M.A.; Turnidge, J. Wild-Type MIC Distributions and Epidemiological Cutoff Values for Amphotericin B and Aspergillus spp. for the CLSI Broth Microdilution Method (M38-A2 Document). Antimicrob. Agents Chemother. 2011, 55, 5150–5154. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Hsieh, M.I.; Choi, P.C.; Wu, C.J. Comparison of the Sensititre YeastOne and CLSI M38-A2 Microdilution Methods in Determining the Activity of Amphotericin B, Itraconazole, Voriconazole, and Posaconazole against Aspergillus Species. J. Clin. Microbiol. 2018, 56, e00780-18. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.J.; Subedi, S.; Halliday, C.L.; Hibbs, D.E.; Lai, F.; Lopez-Ruiz, F.J.; Harper, L.; Park, R.F.; Cuddy, W.S.; Biswas, C.; et al. Surveillance for Azole Resistance in Clinical and Environmental Isolates of Aspergillus Fumigatus in Australia and Cyp51A Homology Modelling of Azole-Resistant Isolates. J. Antimicrob. Chemother. 2018, 73, 2347–2351. [Google Scholar] [CrossRef]
- Chen, Y.C.; Kuo, S.F.; Wang, H.C.; Wu, C.J.; Lin, Y.S.; Li, W.S.; Lee, C.H. Azole Resistance in Aspergillus Species in Southern Taiwan: An Epidemiological Surveillance Study. Mycoses 2019, 62, 1174–1181. [Google Scholar] [CrossRef]
- Guinea, J.; Verweij, P.E.; Meletiadis, J.; Mouton, J.W.; Barchiesi, F.; Arendrup, M.C.; Arikan-Akdagli, S.; Castanheira, M.; Chryssanthou, E.; Friberg, N.; et al. How to: EUCAST Recommendations on the Screening Procedure E.Def 10.1 for the Detection of Azole Resistance in Aspergillus Fumigatus Isolates Using Four-Well Azole-Containing Agar Plates. Clin. Microbiol. Infect. 2019, 25, 681–687. [Google Scholar] [CrossRef]
- Buil, J.B.; Van Der Lee, H.A.L.; Rijs, A.J.M.M.; Zoll, J.; Hovestadt, J.A.M.F.; Melchers, W.J.G.; Verweij, P.E. Single-Center Evaluation of an Agar-Based Screening for Azole Resistance in Aspergillus Fumigatus by Using VIPcheck. Antimicrob. Agents Chemother. 2017, 61, e01250-17. [Google Scholar] [CrossRef]
- van der Linden, J.W.M.; Arendrup, M.C.; Warris, A.; Lagrou, K.; Pelloux, H.; Hauser, P.M.; Chryssanthou, E.; Mellado, E.; Kidd, S.E.; Tortorano, A.M.; et al. Prospective Multicenter International Surveillance of Azole Resistance in Aspergillus Fumigatus. Emerg. Infect. Dis. 2015, 21, 1041. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, K.L.; Mellado, E.; Lass-Flörl, C.; Rodriguez-Tudela, J.L.; Johansen, H.K.; Arendrup, M.C. Environmental Study of Azole-Resistant Aspergillus Fumigatus and Other Aspergilli in Austria, Denmark, and Spain. Antimicrob. Agents Chemother. 2010, 54, 4545–4549. [Google Scholar] [CrossRef] [PubMed]
- Abdolrasouli, A.; Scourfield, A.; Rhodes, J.; Shah, A.; Elborn, J.S.; Fisher, M.C.; Schelenz, S.; Armstrong-James, D. High Prevalence of Triazole Resistance in Clinical Aspergillus Fumigatus Isolates in a Specialist Cardiothoracic Centre. Int. J. Antimicrob. Agents 2018, 52, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Lestrade, P.P.; van der Velden, W.J.F.M.; Bouwman, F.; Stoop, F.J.; Blijlevens, N.M.A.; Melchers, W.J.G.; Verweij, P.E.; Donnelly, J.P. Epidemiology of Invasive Aspergillosis and Triazole-Resistant Aspergillus Fumigatus in Patients with Haematological Malignancies: A Single-Centre Retrospective Cohort Study. J. Antimicrob. Chemother. 2018, 73, 1389–1394. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Drug and Species | Agent/Method-Dependent ECVs (mg/L) | |||
|---|---|---|---|---|
| CLSI a | EUCAST b | Etest c | SYO d | |
| Micafungin | ||||
| Aspergillus fumigatus SC | 0.016 | |||
| Caspofungin | ||||
| Aspergillus fumigatus SC | 0.5 | 0.25 | ||
| Aspergillus terreus SC | 0.125 | 2 | ||
| Aspergillus flavus SC | 0.5 | 0.5 | ||
| Aspergillus niger SC | 0.25 | 0.25 | ||
| Isavuconazole | ||||
| Aspergillus fumigatus SC | 1 | 2 | ||
| Aspergillus terreus SC | 1 | (1) | ||
| Aspergillus flavus SC | 1 | 2 | ||
| Aspergillus niger SC | 4 | 4 | ||
| Aspergillus versicolor SC | 1 | |||
| Aspergillus nidulans SC | 0.25 | |||
| Voriconazole | ||||
| Aspergillus fumigatus SC | 1 | 1 | 0.5 | 1 |
| Aspergillus terreus SC | 2 | 2 | 1 | |
| Aspergillus flavus SC | 2 | 2 | 0.5 | 1 |
| Aspergillus niger SC | 2 | 2 | 1 | |
| Aspergillus nidulans SC | 1 | |||
| Fusarium solani SC | 32 | |||
| Fusarium verticillioides SC | 4 | |||
| Fusarium oxyporum SC | 16 | |||
| Posaconazole | ||||
| Aspergillus fumigatus SC | 0.25 | 0.25 | 0.25 | 0.06 |
| Aspergillus terreus SC | 1 | 0.25 | 0.25 | |
| Aspergillus flavus SC | 0.5 | 0.5 | 0.5 | |
| Aspergillus niger SC | 2 | 0.5 | 0.5 | |
| Aspergillus nidulans SC | 0.5 | |||
| Fusarium solani SC | 32 | |||
| Fusarium verticillioides SC | 2 | |||
| Fusarium oxyporum SC | 8 | |||
| Lichtheimia corymbifera | 2 | |||
| Mucor circinelloides | 4 | |||
| Rhizopus arrhizus | 2 | |||
| Rhizopus microsporus | 2 | |||
| Itraconazole | ||||
| Aspergillus fumigatus SC | 1 | 1 | 2 | |
| Aspergillus terreus SC | 2 | (0.5) | ||
| Aspergillus flavus SC | 1 | 1 | 1 | |
| Aspergillus niger SC | 4 | 2 | 4 | 1 |
| Aspergillus nidulans SC | (1) | 1 | ||
| Fusarium solani SC | 32 | |||
| Fusarium oxyporum SC | 32 | |||
| Rhizopus arrhizus | 2 | |||
| Amphotericin B | ||||
| Aspergillus fumigatus SC | 2 | 11 | 2 a | |
| Aspergillus terreus SC | 4 | 8 | 16 a | |
| Aspergillus flavus SC | 4 | 4 | 8 a | |
| Aspergillus niger SC | 2 | (0.5) | 2 a | |
| Aspergillus versicolor SC | 2 | |||
| Aspergillus nidulans SC | (4) | |||
| Fusarium (Gibberella) fujikuroi SC | (8) | |||
| Fusarium solani SC | 8 | (8) | ||
| Fusarium verticillioides SC | 4 | |||
| Fusarium oxyporum SC | 8 | |||
| Lichtheimia corymbifera | 2 | |||
| Mucor circinelloides | 2 | |||
| Rhizopus arrhizus | 4 | |||
| Rhizopus microsporus | 2 | |||
| Method | Species | No. of Isolates | Anidulafungin | Micafungin | Caspofungin | Isavuconazole | Voriconazole | Posaconazole | Itraconazole | Amphotericin B | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SYO | A. fumigatusSC | 17 | 100% (0) | 41% (3) | [36] | ||||||
| 21 | 100% (+1) | 100% (0) | 95.2% (−2) | [37] | |||||||
| 24 | 95.8% (NA) | 95.8% (NA) | [38] | ||||||||
| A. flavus SC | 9 | 89% (−1) | 0% (3) | [36] | |||||||
| 19 | 100% (+1) | 100% (0) | 94.7% (−1) | [37] | |||||||
| 23 | 78.3% (NA) | 91.3% (NA) | [38] | ||||||||
| A. terreus SC | 13 | 77% (−2) | 54% (2) | [36] | |||||||
| 12 | 100% (+1) | 100% (−2) | 91.7% (−1) | [37] | |||||||
| A. niger SC | 7 | 100% (0) | 85.7% (0) | 100% (−1) | [37] | ||||||
| 7 | 100% (NA) | 100% (NA) | [38] | ||||||||
| A. nidulans SC | 5 | 100% (+1) | 100% (+1) | 100% (−2) | [37] | ||||||
| Aspergillus spp. | 61 | 90.2% (NA) | 93.4% (NA) | [38] | |||||||
| Zygomycetes a | 45 | 93.3% (NA) | 74.4% (NA) | 81.8% (NA) | 79.5% (NA) | 29.5% (NA) | [39] | ||||
| Etest | A. fumigatus SC | 74 | 100% (0) | 96% (0) | 95% (−1) | 89% (−1) | 99% (+1) | [40] | |||
| 24 | 45.8% (NA) | 79.2% (NA) | [38] | ||||||||
| 25 | 100% (NA) | 100% (NA) | 100% (NA) | 100% (NA) | [41] | ||||||
| 26 | 65% (+2) | [42] | |||||||||
| 38 | 82% (NA) | [43] | |||||||||
| 21 | 100% (NA) | 100% (NA) | 90.4% (NA) | [44] | |||||||
| Wild type | 40 | 98 b | [45] | ||||||||
| G54 alterations | 10 | 100% b | [45] | ||||||||
| M220 alterations | 10 | 90% b | [45] | ||||||||
| TR34/L98H | 9 | 33% b | [45] | ||||||||
| TR46/Y121F/T289A | 10 | 100% (NA)b | [45] | ||||||||
| A. flavus SC | 29 | 100% (0) | 100% (0) | 100% (0) | 83% (−1) | 97% (+1) | [40] | ||||
| 23 | 26.1% (NA) | 60.9% (NA) | [38] | ||||||||
| 21 | 100% (+1) | [42] | |||||||||
| 18 | 100% (NA) | 100% (NA) | 77.8% (NA) | [44] | |||||||
| A. terreus SC | 25 | 100% (0) | 96% (0) | 100% (−1) | 64% (−2) | 16% (+2) | [40] | ||||
| 10 | 100% (+1) | [42] | |||||||||
| 17 | 100% (NA) | 100% (NA) | 82.4% (NA) | [44] | |||||||
| A. niger SC | 12 | 100% (0) | 100% (0) | 100% (−1) | 83% (−1) | 100% (+1) | [40] | ||||
| 7 | 71.4% (NA) | 100% (NA) | [38] | ||||||||
| 13 | 100% (NA) | 92% (NA) | 100% (NA) | 100% (NA) | [41] | ||||||
| 9 | 88% (+1) | [42] | |||||||||
| 7 | 100% (NA) | 100% (NA) | 100% (NA) | [44] | |||||||
| A. nidulans SC | 5 | 100% (0) | [42] | ||||||||
| A. glaucus SC | 4 | 50% (NA) | [42] | ||||||||
| 4 | 100% (NA) | 100% (NA) | 50% (NA) | [44] | |||||||
| A. flavipes SC | 2 | 100% (NA) | [42] | ||||||||
| Aspergillus spp. | 32 | 81% (+1) | 72% (0) | 69% (+5) | [46] | ||||||
| 61 | 42.6% (NA) | 73.8% (NA) | [38] | ||||||||
| 77 | 84.4% (+2) | [42] | |||||||||
| 17 | 88% (NA) | [43] | |||||||||
| Fusarium spp. | 20 | 95% (0) | 85% (−1) | 90% (−1) | [47] | ||||||
| 34 | 100% (0) | 100% (0) | 100% (0) | 100% (0) | 94% (0) | [40] | |||||
| Zygomycetes a | 45 | 100% (NA) | 100% (NA) | 81.8% (NA) | 65.9% (NA) | 54.5% (NA) | [39] | ||||
| S. apiospermum SC | 20 | 37% (−2) | 63% (+1) | 90% (−2) | 90% (0) | 80% (+1) | [40] | ||||
| S. prolificans SC | 5 | 100% (0) | 100% (+1) | 100% (0) | 100% (0) | 100% (0) | [40] | ||||
| Scopulariopsis spp. | 8 | 29% (4) | 43% (+5) | 100% (0) | 100% (0) | 75% (+1) | [40] | ||||
| P. lilacium SC | 20 | 100% (0) | 100% (−1) | 100% (0) | 97% (−1) | 100% (0) | [40] | ||||
| Micronaut-AM | Aspergillus spp. | 78 | 99% (0) | 90% (−1) | 87% (−2) | 100% (0) | [48] | ||||
| A. fumigatus SC c | 77 | 58% (−2) | 30% (−3) | 62% (−2) | 100% (0) | [49] | |||||
| VIPcheck d | A. fumigatus SC | 30 | 80% | 83.3% | 80% | [50] | |||||
| A. fumigatus | 91 | 96.7% | 73.6% | 97.8% | [51] | ||||||
| A. fumigatus cryptic species | 30 | 78.8% | 55.8% | 69.2% | [51] |
| Drug (Method-Specific ECV) | Mutations | MIC of All Mutant Isolates (mg/L) | No. Mutant Isolates with MIC > ECV | Refs. |
|---|---|---|---|---|
| Isavuconazole (2 mg/L a) | ||||
| G54 alterations G54E, G54R, G54V, G54W, G54R N248K | 0.125–1 | 0/10 | [45] | |
| M220 alterations M220K, M220T, M220V, M220I, M220I V101F | 0.5–>32 | 2/10 | [45] | |
| TR34/L98H | >32 | 9/9 | [45] | |
| TR46/Y121F T289A | >32 | 10/10 | [45] | |
| Total | 0.125–>32 | 21/39 (53.8%) | ||
| Itraconazole (2 mg/L) | ||||
| cyp51A mutants | 0.06–>16 | 78/81 | [29] | |
| G54E, M220R, M220I, TR/L98H | 12–>32 | 6/6 | [99] | |
| cyp51A mutants | 1.5 | 0/3 | [100] | |
| G448S | 0.5 | 0/1 | [29] | |
| M220K | 2 | 0/1 | [29] | |
| I301T | ≤0.06 | 0/1 | [29] | |
| Total | ≤0.06–>32 | 84/93 (90.3%) | ||
| Voriconazole (0.5 mg/L) | ||||
| cyp51A mutants | 0.06–>16 | 49/75 | [29] | |
| G54E, M220R, M220I, TR/L98H | 0.047–1.5 | 3/6 | [99] | |
| cyp51A mutants | 0.047 | 0/3 | [100] | |
| I301T | <0.06 | 0/1 | [29] | |
| TR34 | 0.125–>4 | 35/38 | [29] | |
| G54E/R/W | ≤0.06–0.5 | 0/12 | [29] | |
| M220I/K//R/T/V | 0.125–>4 | 3/11 | [29] | |
| G138C | 0.25 | 0/1 | [29] | |
| Total | 0.047–>16 | 90/147 (61.2%) | ||
| Posaconazole (0.25 mg/L) | ||||
| G54E, M220R, M220I, TR/L98H | 1–>16 | 6/6 | [99] | |
| TR34/L98H | 0.5–>16 | 13/13 | [30] | |
| G54E/R/V/W | 2–>16 | 5/5 | [30] | |
| M220I/R/T/V/K | 0.25–>16 | 3/4 | [30] | |
| G448S | 0.25–1 | 4/5 | [30] | |
| cyp51A mutants | 0.023 | 0/3 | [100] | |
| Total | 0.023–>16 | 31/36 (86.1%) | ||
| Micafungin (0.016 mg/L) b | ||||
| fks alterations S678P | 0.004–1 | 2/3 | [101] | |
| Total | 0.004–1 | 2/3 (66.7%) | ||
| Caspofungin (0.25 mg/L) b | ||||
| fks alterations S678P | 2–8 | 3/3 | [101] | |
| Total | 2–8 | 3/3 (100%) |
| Drug (Method-Specific ECV) | Mutations | MIC of All Mutant Isolates (mg/L) | No. Mutant Isolates with MIC > ECV | Refs. |
|---|---|---|---|---|
| Itraconazole (0.5 mg/L a) | ||||
| TR34/L98H | 2–>16 | 5/5 | [37] | |
| TR46/Y121F T289A | 0.5–1 | 4/5 | [37] | |
| TR34 | >16 | 24/24 | [105] | |
| G54R | 1 | 2/2 | [106] | |
| TR34/L98H | >16 | 1/1 | [106] | |
| cyp51A mutants | 0.125–>8 | 9/21 | [37] | |
| TR34/L98H, S297T, F495I, TR46/Y121F T289A | 0.125–>16 | 4/7 | [107] | |
| Total | 0.125–>16 | 45/65 (69.2%) | ||
| Voriconazole (1 mg/L) | ||||
| cyp51A mutants | 0.125–>16 | 21/39 | [29] | |
| I301T | 0.125 | 0/1 | [29] | |
| TR34 | 1 | 0/1 | [29] | |
| G54E/R/W | 0.125–0.5 | 0/5 | [29] | |
| M220I/K//R/T/V | 0.25–0.5 | 0/5 | [29] | |
| G138C | 0.25 | 0/1 | [29] | |
| TR34/L98H | 1–8 | 4/5 | [37] | |
| TR46/Y121F T289A | 8–>8 | 5/5 | [37] | |
| TR34 | 1–8 | >12/24 * | [105] | |
| G54R | 0.125 | 0/2 | [106] | |
| TR34/L98H | 8 | 1/1 | [106] | |
| cyp51A mutants | 0.125–>8 | 10/21 | [37] | |
| TR34/L98H, S297T, F495I, TR46/Y121F T289A | 1–>8 | 5/7 | [107] | |
| Total | 0.125–>16 | 67/116 (57.7%) | ||
| Posaconazole (0.06 mg/L) | ||||
| TR34/L98H | 0.5–1 | 5/5 | [37] | |
| TR46/Y121F T289A | 1 | 5/5 | [37] | |
| TR34 | 0.25–1 | 24/24 | [105] | |
| G54R | 1 | 2/2 | [106] | |
| TR34/L98H | 1 | 1/1 | [106] | |
| cyp51A mutants | 0.5–1 | 10/10 | [37] | |
| TR34/L98H, S297T, F495I, TR46/Y121F T289A | 0.125–0.25 | 7/7 | [107] | |
| Total | 0.25–1 | 54/54 (100%) |
| Drug | Mutations | MIC of All Mutant Isolates a (mg/L) | No. Mutant Isolates Detected | Refs. |
|---|---|---|---|---|
| Itraconazole | ||||
| TR34/L98H (28/30) | 2–>16 | 25/30 | [50] | |
| TR/L98H | >8 | 3/4 | [111] | |
| TR34/L98H G54R, N248K | 1–>16 | 3/3 | [106] | |
| TR34/L98H, TR46/Y121F T289A | 1–>8 | 47/47 | [110] | |
| TR34/L98H, TR46/Y121F T289A, G54, M220 | 0.5–>16 | 32/39 | [71] | |
| TR34/L98H, TR46/Y121F, TR53 T289A, G54, M220, P216, G138, G448 | NA | 29/33 | [109] | |
| cyp51A mutations | >16 | 20/20 | [109] | |
| Total | 0.5–>32 | 159/176 (90.3%) | ||
| Voriconazole | ||||
| TR34/L98H (28/30) | 2–4 | 23/30 | [50] | |
| TR/L98H | 4 | 1/4 | [111] | |
| TR34/L98H G54R, N248K | 0.125–8 | 2/3 | [106] | |
| TR34/L98H, TR46/Y121F T289A | 2 | 47/47 | [110] | |
| TR34/L98H, TR46/Y121F T289A, G54, M220 | 0.125–>16 | 23/39 | [71] | |
| TR34/L98H, TR46/Y121F, TR53 T289A, G54, M220, P216, G138, G448 | NA | 29/33 | [109] | |
| cyp51A mutations | 0.5–>16 | 16/20 | [109] | |
| Total | 0.03–>16 | 141/176 (80.1%) | ||
| Posaconazole | ||||
| TR34/L98H (28/30) | 0.5–1 | 7/30 | [50] | |
| TR/L98H | 0.5 | 1/4 | [111] | |
| TR34/L98H G54R, N248K | 1 | 2/3 | [106] | |
| TR34/L98H, TR46/Y121F T289A | 0.25–1 | 47/47 | [110] | |
| TR34/L98H, TR46/Y121F T289A, G54, M220 | 0.125–>16 | 15/39 | [71] | |
| TR34/L98H, TR46/Y121F, TR53 T289A, G54, M220, P216, G138, G448 | NA | 29/33 | [109] | |
| cyp51A mutations | 0.5–2 | 14/20 | [109] | |
| Total | 0.125–>16 | 115/174 (66%) |
| Method | Isavuconazole | Itraconazole | Voriconazole | Posaconazole | Micafungin | Caspofungin |
|---|---|---|---|---|---|---|
| Etest | 53.8% (21/39) | 90.3% (84/93) | 61.2% (90/147) | 86.1% (31/36) | 66.7% (2/3) | 100% (3/3) |
| SYO | 69.2% (45/65) | 57.7% (67/116) | 100% (54/54) | |||
| VIPcheck | 90.3% (159/176) | 80.1% (141/176) | 66% (115/174) | |||
| Micronaut-AM | 88% (22/25) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paranos, P.; Espinel-Ingroff, A.; Meletiadis, J. Commercial Methods for Antifungal Susceptibility Testing of Saprophytic Molds: Can They Be Used to Detect Resistance? J. Fungi 2024, 10, 214. https://doi.org/10.3390/jof10030214
Paranos P, Espinel-Ingroff A, Meletiadis J. Commercial Methods for Antifungal Susceptibility Testing of Saprophytic Molds: Can They Be Used to Detect Resistance? Journal of Fungi. 2024; 10(3):214. https://doi.org/10.3390/jof10030214
Chicago/Turabian StyleParanos, Paschalis, Ana Espinel-Ingroff, and Joseph Meletiadis. 2024. "Commercial Methods for Antifungal Susceptibility Testing of Saprophytic Molds: Can They Be Used to Detect Resistance?" Journal of Fungi 10, no. 3: 214. https://doi.org/10.3390/jof10030214
APA StyleParanos, P., Espinel-Ingroff, A., & Meletiadis, J. (2024). Commercial Methods for Antifungal Susceptibility Testing of Saprophytic Molds: Can They Be Used to Detect Resistance? Journal of Fungi, 10(3), 214. https://doi.org/10.3390/jof10030214