
The Oxidative Stress Response Highly Depends on Glucose and Iron Availability in Aspergillus fumigatus



Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Manipulation | Function of the Gene(s) | Phenotype of the Mutant | Reference | |
|---|---|---|---|---|
| ΔoxrA | oxidation resistance protein | decreased H2O2 tolerance | hypovirulence (mice) | [10] |
| Δaspf3 | peroxiredoxin | decreased H2O2 and O2− tolerance | hypovirulence (mice) | [11] |
| Δpes1 | nonribosomal peptide synthetase | decreased H2O2 and HOCl tolerance | hypovirulence (Galleria mellonella) | [12] |
| ΔppoA, ΔppoB, ΔppoC | fatty acid oxygenases | increased H2O2 tolerance | hypervirulence (mice) | [13] |
| Δsod1Δsod2Δsod3 | SODs | decreased menadione tolerance | no effect on virulence (mice) | [14] |
| ΔcatA | catalase | decreased H2O2 tolerance | no effect on virulence (rat) | [15] |
| Δcat1, Δcat2 | catalase and catalase-peroxidase | slightly decreased H2O2 tolerance | slightly delayed infection (rat) | [15] |
| Δsho1 | transmembrane osmosensor | decreased H2O2 and menadione tolerance | no effect on virulence (mice) | [19] |
| pkcAG579R | protein kinase | decreased paraquat and menadione tolerance | no effect on virulence (mice) | [21] |
| ΔmpkA | protein kinase | increased H2O2 and decreased menadione tolerance | no effect on virulence (mice) | [20] |
| Δskn7 | transcription factor | decreased H2O2 and t-butyl hydroperoxide tolerance | no effect on virulence (mice) | [17] |
| DyapA | transcription factor | decreased H2O2 and menadione tolerance | no effect on virulence (mice) | [18] |
2. Materials and Methods
2.1. Strain, Culturing Conditions
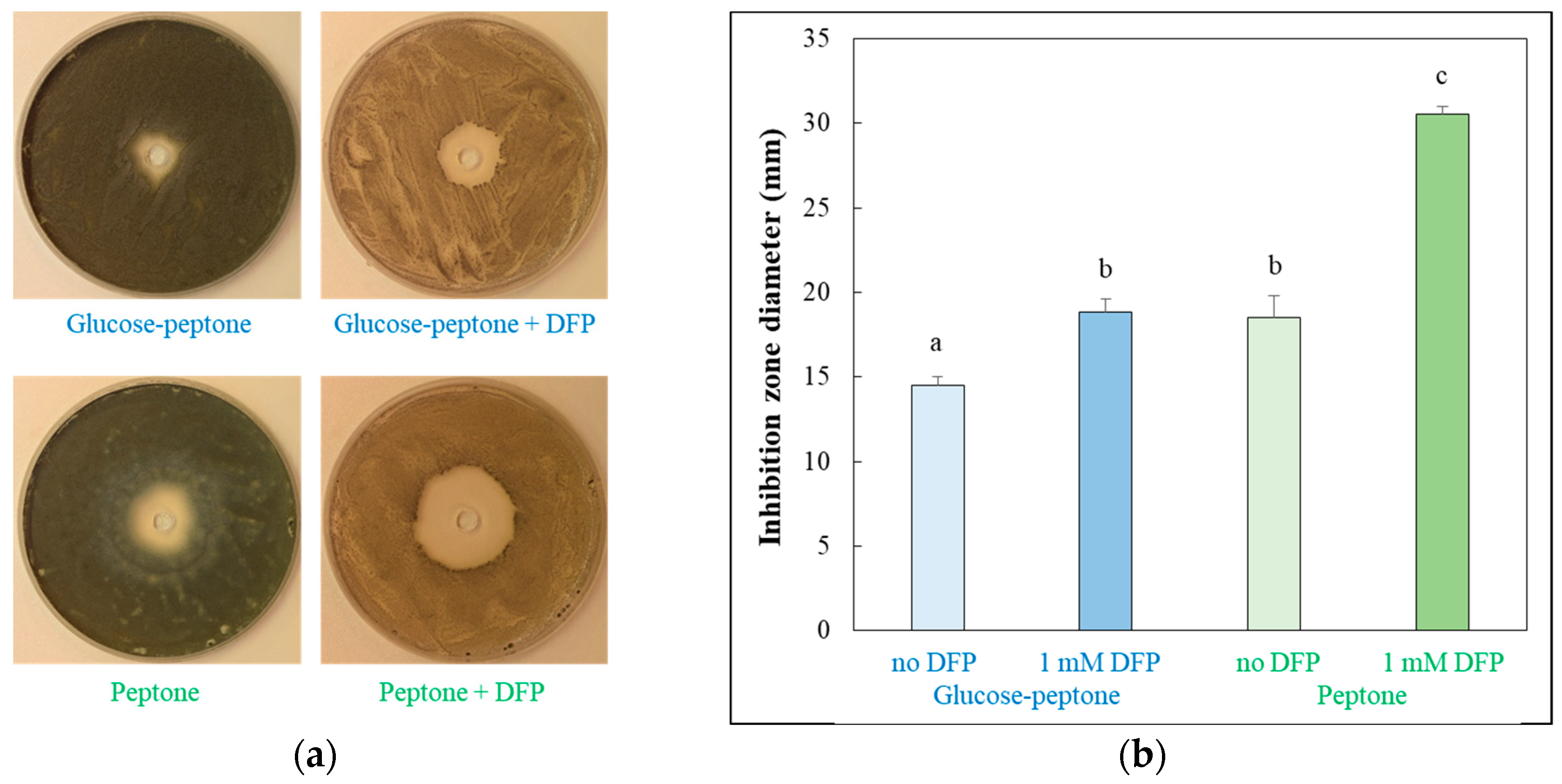
2.2. Measuring H2O2 Stress Sensitivity of Surface Cultures with Well Diffusion Assay
2.3. Detecting Growth, Redox Imbalance and Triacetylfusarinine C Production
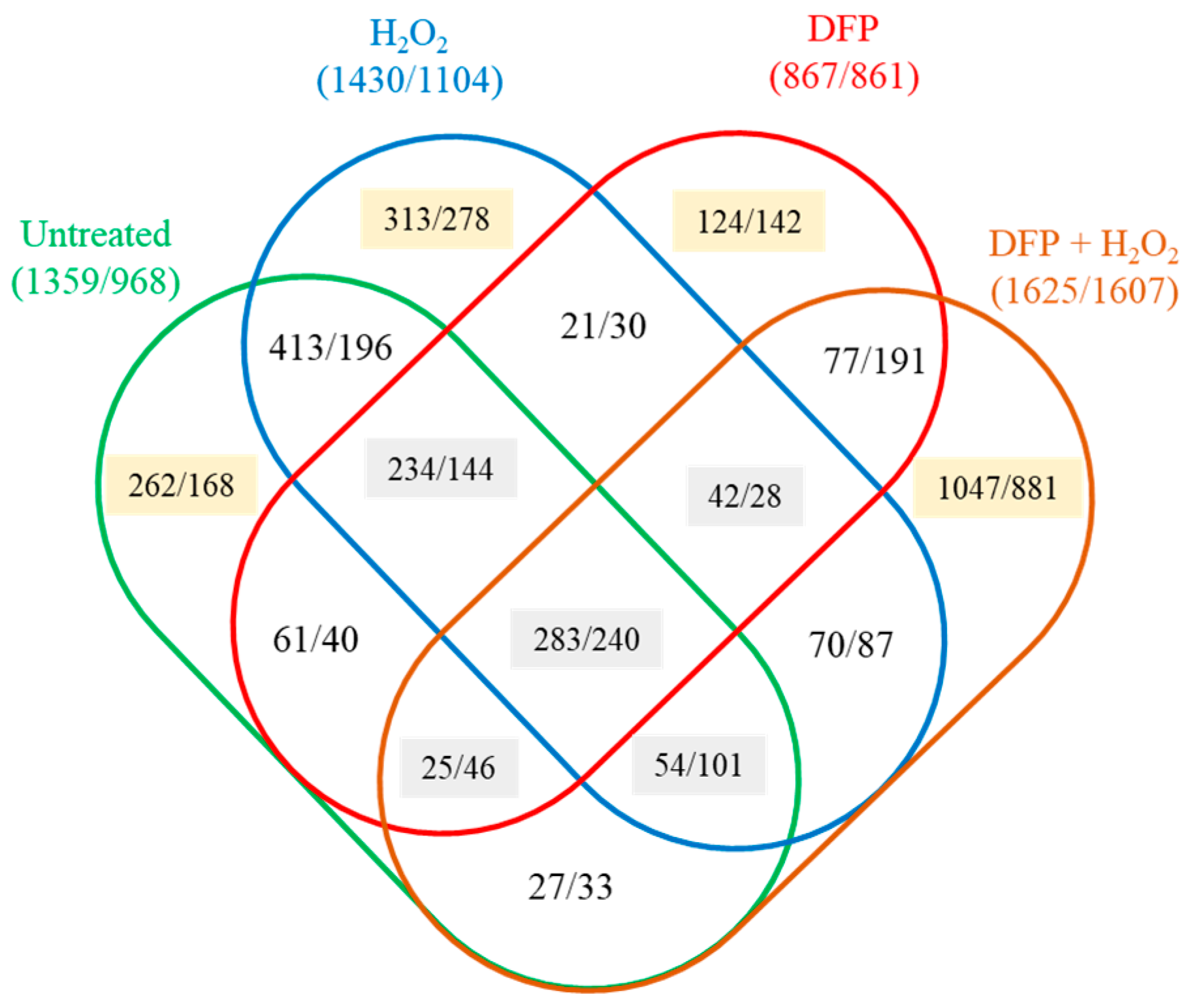
2.4. High Throughput RNA Sequencing
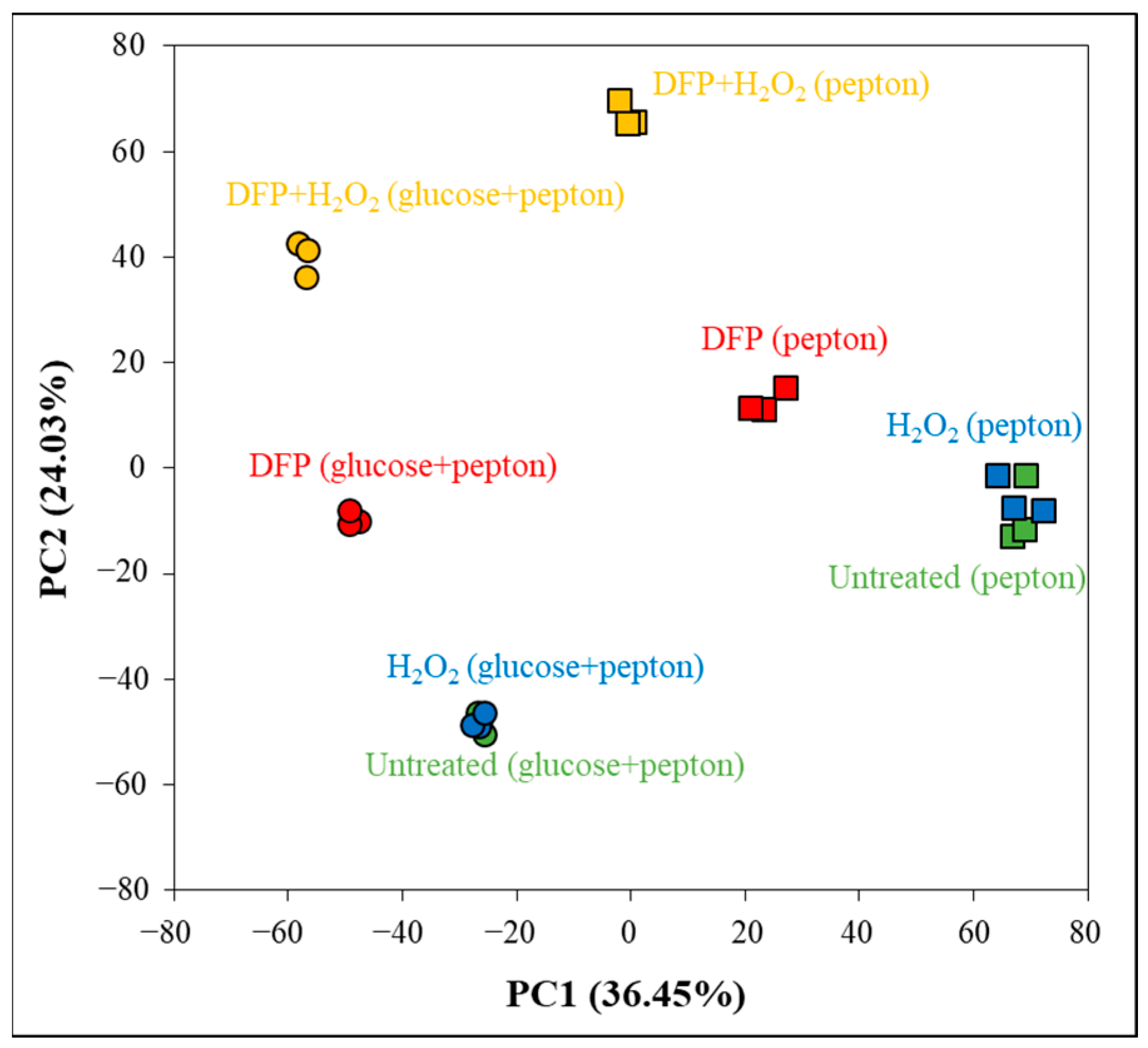
2.5. Evaluation of Transcriptome Data
3. Results
3.1. Effects of Glucose Withdrawal
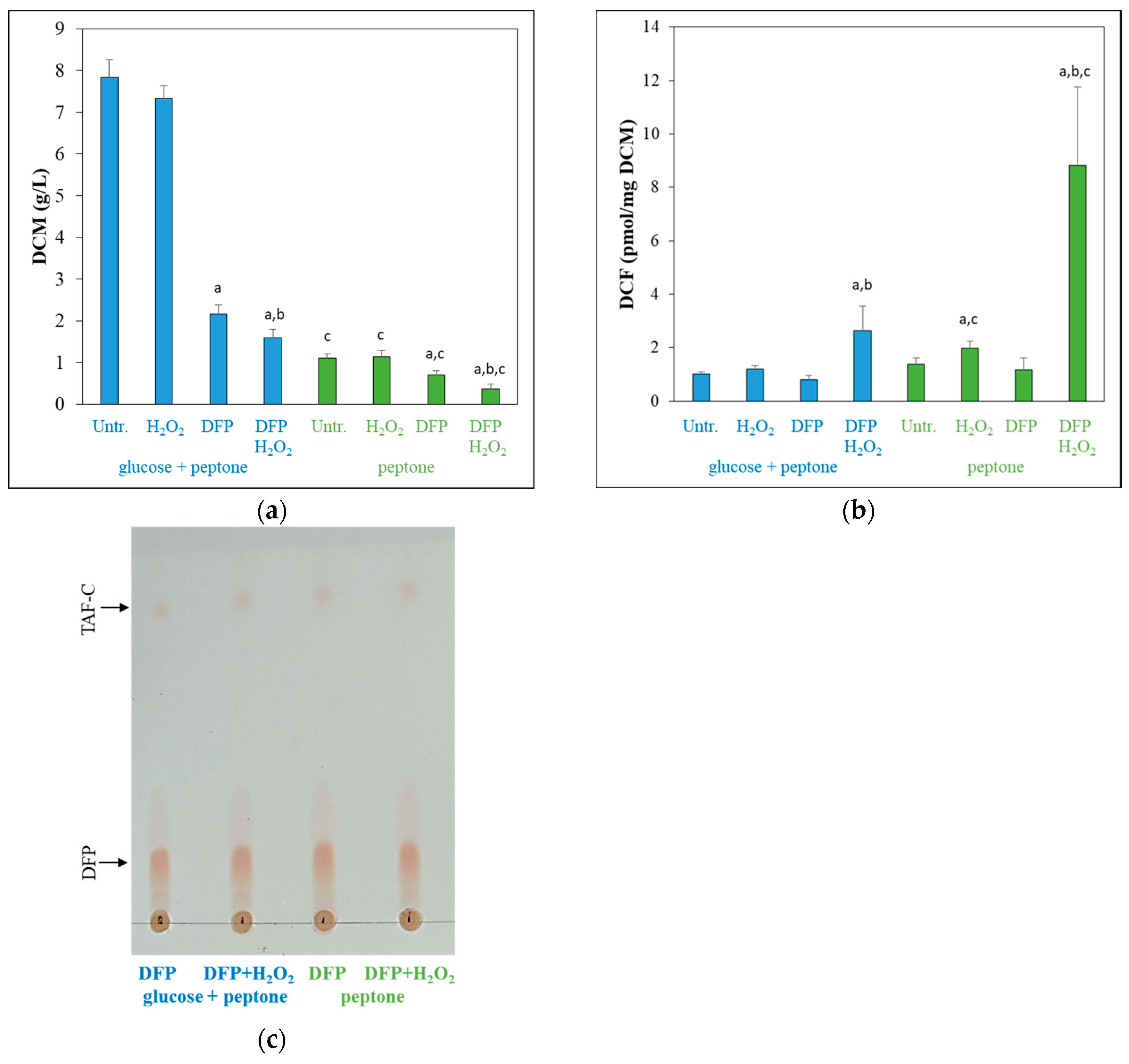
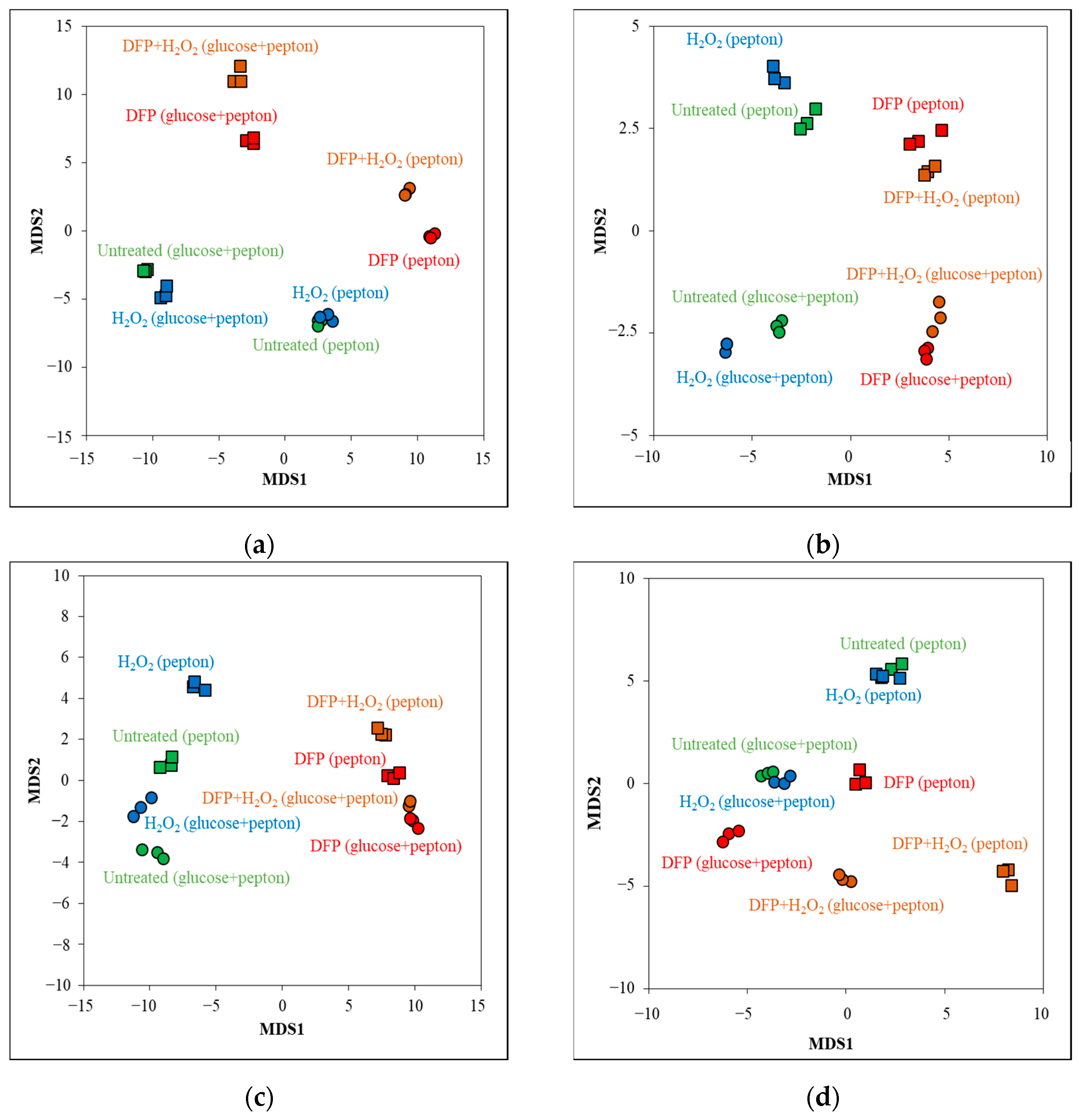
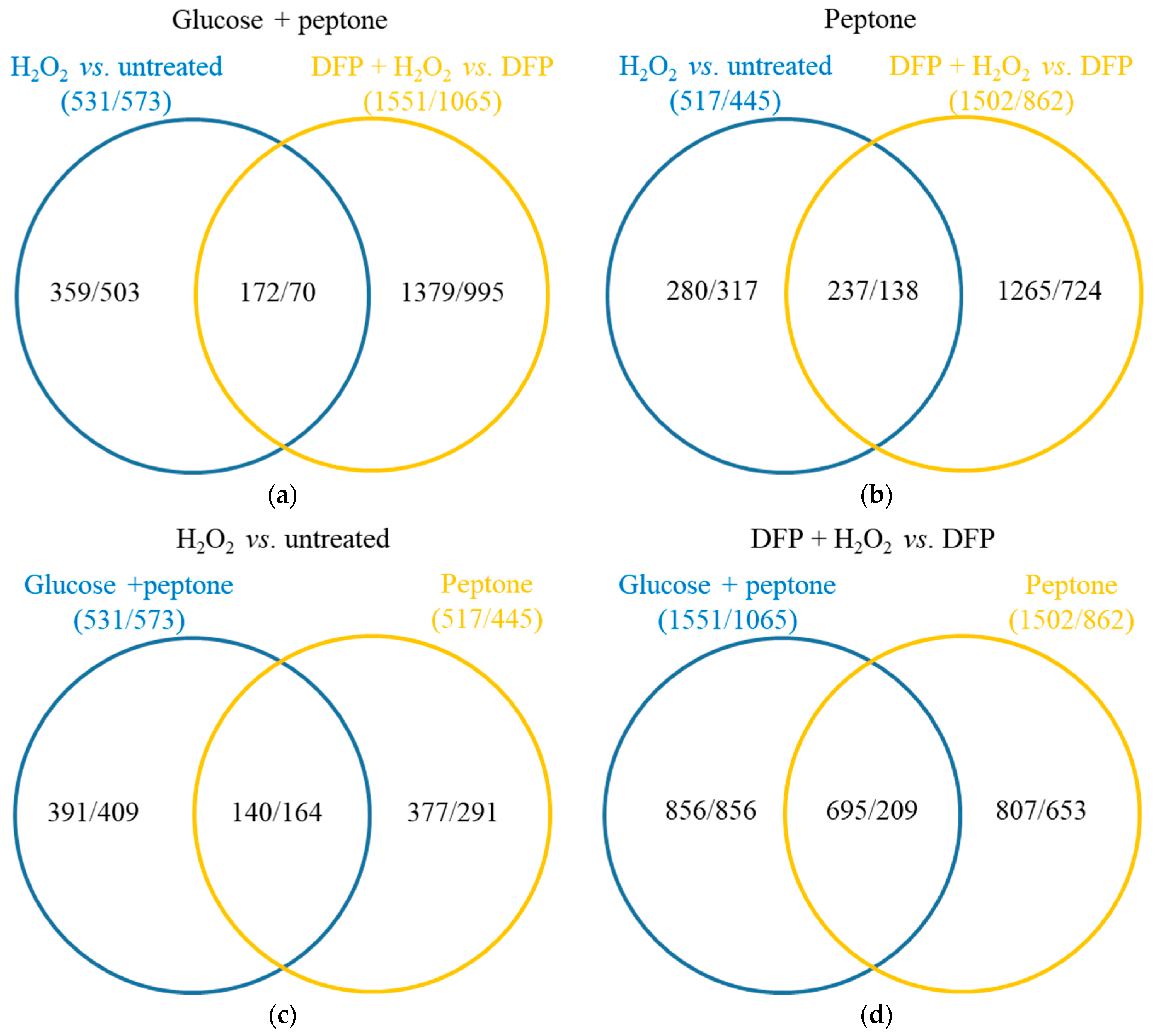
3.2. Effects of DFP Treatment in the Presence and Absence of Glucose
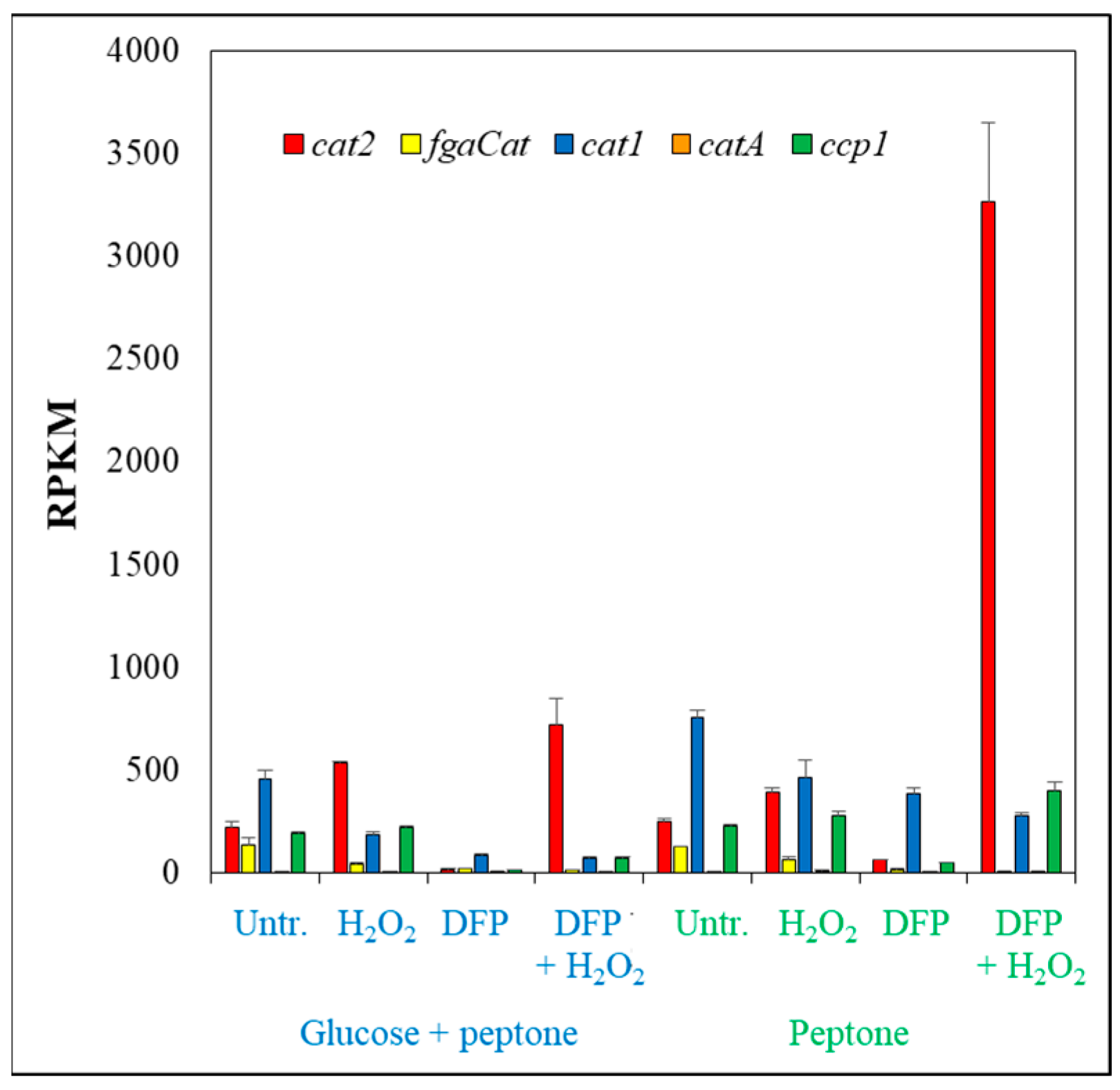
3.3. Oxidative Stress Responses of the Cultures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases-estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, T.R.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef]
- Howard, S.J.; Arendrup, M.C. Acquired antifungal drug resistance in Aspergillus fumigatus: Epidemiology and detection. Med. Mycol. 2011, 49, S90–S95. [Google Scholar] [CrossRef] [PubMed]
- Burks, C.; Darby, A.; Gómez Londoño, L.; Momany, M.; Brewer, M.T. Azole-resistant Aspergillus fumigatus in the environment: Identifying key reservoirs and hotspots of antifungal resistance. PLoS Pathog. 2021, 17, e1009711. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Goldman, G.H. The contribution of Aspergillus fumigatus stress responses to virulence and antifungal resistance. J. Microbiol. 2016, 54, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, S.; Weber, M.; Stein, P.; Bosmann, M.; Stassen, M.; Kreft, A.; Schild, H.; Radsak, M.P. Oxidative burst and neutrophil elastase contribute to clearance of Aspergillus fumigatus pneumonia in mice. Immunobiology 2014, 219, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hatinguais, R.; Pradhan, A.; Brown, G.D.; Brown, A.J.P.; Warris, A.; Shekhova, E. Mitochondrial reactive oxygen species regulate immune responses of macrophages to Aspergillus fumigatus. Front. Immunol. 2021, 12, 641495. [Google Scholar] [CrossRef] [PubMed]
- Warris, A.; Ballou, E.R. Oxidative responses and fungal infection biology. Semin. Cell Dev. Biol. 2019, 89, 34–46. [Google Scholar] [CrossRef]
- Henriet, S.; Verweij, P.E.; Holland, S.M.; Warris, A. Invasive fungal infections in patients with chronic granulomatous disease. Adv. Exp. Med. Biol. 2013, 764, 27–55. [Google Scholar] [CrossRef]
- Zhai, P.; Shi, L.; Zhong, G.; Jiang, J.; Zhou, J.; Chen, X.; Dong, G.; Zhang, L.; Li, R.; Song, J. The OxrA protein of Aspergillus fumigatus is required for the oxidative stress response and fungal pathogenesis. Appl. Environ. Microbiol. 2021, 87, e0112021. [Google Scholar] [CrossRef]
- Hillmann, F.; Bagramyan, K.; Straßburger, M.; Heinekamp, T.; Hong, T.B.; Bzymek, K.P.; Williams, J.C.; Brakhage, A.A.; Kalkum, M. The crystal structure of peroxiredoxin asp f3 provides mechanistic insight into oxidative stress resistance and virulence of Aspergillus fumigatus. Sci. Rep. 2016, 6, 33396. [Google Scholar] [CrossRef]
- Reeves, E.P.; Reiber, K.; Neville, C.; Scheibner, O.; Kavanagh, K.; Doyle, S. A nonribosomal peptide synthetase (Pes1) confers protection against oxidative stress in Aspergillus fumigatus. FEBS J. 2009, 273, 3038–3053. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Bok, J.W.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Keller, N.P. Aspergillus cyclooxygenase-like enzymes are associated with prostaglandin production and virulence. Infect. Immun. 2005, 73, 4548–4559. [Google Scholar] [CrossRef] [PubMed]
- Lambou, K.; Lamarre, C.; Beau, R.; Dufour, N.; Latge, J.P. Functional analysis of the superoxide dismutase family in Aspergillus fumigatus. Mol. Microbiol. 2010, 75, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Paris, S.; Wysong, D.; Debeaupuis, J.P.; Shibuya, K.; Philippe, B.; Diamond, R.D.; Latgé, J.P. Catalases of Aspergillus fumigatus. Infect. Immun. 2003, 71, 3551–3562. [Google Scholar] [CrossRef] [PubMed]
- Calera, J.A.; Paris, S.; Monod, M.; Hamilton, A.J.; Debeaupuis, J.P.; Diaquin, M.; López-Medrano, R.; Leal, F.; Latgé, J.P. Cloning and disruption of the antigenic catalase gene of Aspergillus fumigatus. Infect. Immun. 1997, 65, 4718–4724. [Google Scholar] [CrossRef]
- Lamarre, C.; Ibrahim-Granet, O.; Du, C.; Calderone, R.; Latgé, J.P. Characterization of the SKN7 ortholog of Aspergillus fumigatus. Fungal Genet. Biol. 2007, 44, 682–690. [Google Scholar] [CrossRef]
- Lessing, F.; Kniemeyer, O.; Wozniok, I.; Loeffler, J.; Kurzai, O.; Haertl, A.; Brakhage, A.A. The Aspergillus fumigatus transcriptional regulator AfYap1 represents the major regulator for defense against reactive oxygen intermediates but is dispensable for pathogenicity in an intranasal mouse infection model. Eukaryot. Cell 2007, 6, 2290–2302. [Google Scholar] [CrossRef]
- Ma, Y.; Qiao, J.; Liu, W.; Wan, Z.; Wang, X.; Calderone, R.; Li, R. The sho1 sensor regulates growth, morphology, and oxidant adaptation in Aspergillus fumigatus but is not essential for development of invasive pulmonary aspergillosis. Infect. Immun. 2008, 76, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Valiante, V.; Heinekamp, T.; Jain, R.; Härtl, A.; Brakhage, A.A. The mitogen-activated protein kinase MpkA of Aspergillus fumigatus regulates cell wall signaling and oxidative stress response Fungal Genet. Biol. 2008, 45, 618–627. [Google Scholar] [CrossRef]
- Rocha, M.C.; Godoy, K.F.; de Castro, P.A.; Hori, J.I.; Bom, V.L.; Brown, N.A.; Cunha, A.F.; Goldman, G.H.; Malavazi, I. The Aspergillus fumigatus pkcAG579R mutant is defective in the activation of the cell wall integrity pathway but is dispensable for virulence in a neutropenic mouse infection model. PLoS ONE 2015, 10, e0135195. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Sümegi-Győri, V.M.; Páll, K.; Gila, B.C.; Pócsi, I. Effect of the combinatorial iron-chelation and oxidative stress on the growth of Aspergillus species. Res. Microbiol. 2022, 173, 103969. [Google Scholar] [CrossRef] [PubMed]
- Desoubeaux, G.; Cray, C. Animal Models of Aspergillosis. Comp. Med. 2018, 68, 109–123. [Google Scholar] [PubMed]
- Ene, I.V.; Adya, A.K.; Wehmeier, S.; Brand, A.C.; MacCallum, D.M.; Gow, N.A.; Brown, A.J. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Antal, K.; Gila, B.; Jónás, A.P.; Pócsi, I. Stress responses elicited by glucose withdrawal in Aspergillus fumigatus. J. Fungi 2022, 8, 1226. [Google Scholar] [CrossRef]
- He, R.; Wei, P.; Odiba, A.S.; Gao, L.; Usman, S.; Gong, X.; Wang, B.; Wang, L.; Jin, C.; Lu, G.; et al. Amino sugars influence Aspergillus fumigatus cell wall polysaccharide biosynthesis, and biofilm formation through interfering galactosaminogalactan deacetylation. Carbohydr. Polym. 2024, 324, 121511. [Google Scholar] [CrossRef]
- Kurucz, V.; Krüger, T.; Antal, K.; Dietl, A.M.; Haas, H.; Pócsi, I.; Kniemeyer, O.; Emri, T. Additional oxidative stress reroutes the global response of Aspergillus fumigatus to iron depletion. BMC Genom. 2018, 19, 357. [Google Scholar] [CrossRef]
- Brown, A.J.P.; Cowen, L.E.; di Pietro, A.; Quinn, J. Stress adaptation. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Forgács, K.; Pócsi, I. Biologia futura: Combinatorial stress responses in fungi. Biol. Futur. 2022, 73, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shi, L.; Wang, S.; Wang, Y.; Zhu, Y.; Jiang, J.; Li, R. Acidic/alkaline stress mediates responses to azole drugs and oxidative stress in Aspergillus fumigatus. Microbiol. Spectr. 2022, 10, e0199921. [Google Scholar] [CrossRef] [PubMed]
- Brantl, V.; Boysen, J.M.; Yap, A.; Golubtsov, E.; Ruf, D.; Heinekamp, T.; Straßburger, M.; Dichtl, K.; Haas, H.; Hillmann, F.; et al. Peroxiredoxin Asp f3 is essential for Aspergillus fumigatus to overcome iron limitation during infection. mBio 2021, 12, e0097621. [Google Scholar] [CrossRef]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild-type and mutant stocks of Aspergillus nidulans. Genetics 1965, 52, 233–246. [Google Scholar] [CrossRef]
- Emri, T.; Molnár, Z.; Pócsi, I. The appearances of autolytic and apoptotic markers are concomitant but differently regulated in carbon-starving Aspergillus nidulans cultures. FEMS Microbiol. Lett. 2005, 251, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Sámi, L.; Emri, T.; Pócsi, I. Autolysis and aging of Penicillium chrysogenum cultures under carbon starvation: III: Glutathione metabolism and formation of reactive oxygen species. Mycol. Res. 2001, 105, 1246–1250. [Google Scholar] [CrossRef]
- Chomczynski, P.A. Reagent for the Single-Step Simultaneous Isolation of RNA, DNA and Proteins from Cell and Tissue Samples. Biotechniques 1993, 15, 532–534, 536–537. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Sabiha, Y.; Loss, O.; Eisendle, M.; Wallner, A.; Arst, H.N., Jr.; Haynes, K.; Haas, H. Distinct roles for intra- and extracellular siderophores during Aspergillus fumigatus infection. PLoS Pathog. 2007, 3, e128. [Google Scholar] [CrossRef]
- Haas, H. Iron—A key nexus in the virulence of Aspergillus fumigatus. Front. Microbiol. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Magnani, T.; Soriani, F.M.; Martins, V.P.; Nascimento, A.M.; Tudella, V.G.; Curti, C.; Uyemura, S.A. Cloning and functional expression of the mitochondrial alternative oxidase of Aspergillus fumigatus and its induction by oxidative stress. FEMS Microbiol. Lett. 2007, 271, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Magnani, T.; Soriani, F.M.; de Paulo Martins, V.; Policarpo, A.C.; Sorgi, C.A.; Faccioli, L.H.; Curti, C.; Uyemura, S.A. Silencing of mitochondrial alternative oxidase gene of Aspergillus fumigatus enhances reactive oxygen species production and killing of the fungus by macrophages. J. Bioenerg. Biomembr. 2008, 40, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Grahl, N.; Dinamarco, T.M.; Willger, S.D.; Goldman, G.H.; Cramer, R.A. Aspergillus fumigatus mitochondrial electron transport chain mediates oxidative stress homeostasis, hypoxia responses and fungal pathogenesis. Mol. Microbiol. 2012, 84, 383–399. [Google Scholar] [CrossRef]
- Qiao, J.; Liu, W.; Li, R. Truncated Afyap1 attenuates antifungal susceptibility of Aspergillus fumigatus to voriconazole and confers adaptation of the fungus to oxidative stress. Mycopathologia 2010, 170, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.P.; Horta, M.A.C.; Goldman, G.H. Genetic interactions between Aspergillus fumigatus basic leucine zipper (bZIP) transcription factors AtfA, AtfB, AtfC, and AtfD. Front. Fungal Biol. 2021, 2, 632048. [Google Scholar] [CrossRef]
- Fones, H.; Preston, G.M. Reactive oxygen and oxidative stress tolerance in plant pathogenic Pseudomonas. FEMS Microbiol. Lett. 2012, 327, 1–8. [Google Scholar] [CrossRef]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster—From microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Sun, J.; Xiao, S.; Xue, C. The tug-of-war on iron between plant and pathogen. Phytopathol. Res. 2023, 5, 61. [Google Scholar] [CrossRef]
- Allert, S.; Brunke, S.; Hube, B. In vivo transcriptional profiling of human pathogenic fungi during infection: Reflecting the real life? PLoS Pathog. 2016, 12, e1005471. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim-Granet, O.; Dubourdeau, M.; Latgé, J.P.; Ave, P.; Huerre, M.; Brakhage, A.A.; Brock, M. Methylcitrate synthase from Aspergillus fumigatus is essential for manifestation of invasive aspergillosis. Cell. Microbiol. 2008, 10, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol. Asp. Med. 2020, 75, 100864. [Google Scholar] [CrossRef] [PubMed]
- Misslinger, M.; Hortschansky, P.; Brakhage, A.A.; Haas, H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118885. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.; Shadkchan, Y.; Osherov, N.; Krappmann, S. The essential thioredoxin reductase of the human pathogenic mold Aspergillus fumigatus is a promising antifungal target. Front. Microbiol. 2020, 11, 1383. [Google Scholar] [CrossRef] [PubMed]
- Dlouhy, A.C.; Outten, C.E. The iron metallome in eukaryotic organisms. Met. Ions Life Sci. 2013, 12, 241–278. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Orasch, T.; Shadkchan, Y.; Caballero, P.; Pfister, J.; Sastré-Velásquez, L.E.; Gsaller, F.; Decristoforo, C.; Osherov, N.; Haas, H. Uptake of the siderophore triacetylfusarinine C, but not fusarinine C, is crucial for virulence of Aspergillus fumigatus. mBio 2022, 13, e0219222. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Joechl, C.; Rogers, T.; Arst, H.N.; Haynes, K.; Haas, H. Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence. J. Exp. Med. 2004, 200, 1213–1219. [Google Scholar] [CrossRef]
- Yasmin, S.; Alcazar-Fuoli, L.; Gründlinger, M.; Puempel, T.; Cairns, T.; Blatzer, M.; Lopez, J.F.; Grimalt, J.O.; Bignell, E.; Haas, H. Mevalonate governs interdependency of ergosterol and siderophore biosyntheses in the fungal pathogen Aspergillus fumigatus. Proc. Natl. Acad. Sci. USA 2012, 109, E497–E504. [Google Scholar] [CrossRef]
- Beinert, H.; Holm, R.H.; Münck, E. Iron–sulfur clusters: Nature’s modular, multipurpose structures. Science 1997, 277, 653–659. [Google Scholar] [CrossRef]
- Stiban, J.; So, M.; Kaguni, L.S. Iron-sulfur clusters in mitochondrial metabolism: Multifaceted roles of a simple cofactor. Biochemistry 2016, 81, 1066–1080. [Google Scholar] [CrossRef]
- Read, A.D.; Bentley, R.E.; Archer, S.L.; Dunham-Snary, K.J. Mitochondrial iron-sulfur clusters: Structure, function, and an emerging role in vascular biology. Redox Biol. 2021, 47, 102164. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.M.; Martínez-Pastor, M.T.; Puig, S. Iron in translation: From the beginning to the end. Microorganisms 2021, 9, 1058. [Google Scholar] [CrossRef] [PubMed]
- Mettert, E.L.; Kiley, P.J. Fe-S proteins that regulate gene expression. Biochim. Biophys. Acta 2015, 1853, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Outten, C.E.; Albetel, A.N. Iron sensing and regulation in Saccharomyces cerevisiae: Ironing out the mechanistic details. Curr. Opin. Microbiol. 2013, 16, 662–668. [Google Scholar] [CrossRef]








| Gene Sets | Peptone vs. Glucose + Peptone | DFP Pretreated vs. Not Pretreated | H2O2 Treated vs. Untreated | ||||
|---|---|---|---|---|---|---|---|
| Glucose + Peptone | Peptone | Iron Supplemented | DFP Pretreated | ||||
| Glucose + Peptone | Peptone | Glucose + Peptone | Peptone | ||||
| Iron acquisition | - | upregulated | upregulated | downregulated | upregulated | - | - |
| RIA * | - | upregulated | upregulated | downregulated | - | - | - |
| Siderophore cluster | up/downregulated | upregulated | upregulated | upregulated | upregulated | upregulated | upregulated |
| Fe-S cluster assembly | - | - | - | - | - | upregulated | upregulated |
| Fe-S cluster protein | - | downregulated | downregulated | upregulated | - | - | upregulated |
| Heme biosynthesis | - | - | - | - | - | - | - |
| Heme binding proteins | up/downregulated | downregulated | up/downregulated | downregulated | - | - | downregulated |
| Catalases and peroxidases | - | downregulated | downregulated | downregulated | - | - | upregulated |
| TGG * system | - | - | - | - | - | upregulated | upregulated |
| TCA * cycle | - | downregulated | downregulated | upregulated | - | - | - |
| Respiration | - | downregulated | - | - | - | - | - |
| Ergosterol biosynthesis | - | upregulated | upregulated | - | - | - | downregulated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emri, T.; Antal, K.; Varga, K.; Gila, B.C.; Pócsi, I. The Oxidative Stress Response Highly Depends on Glucose and Iron Availability in Aspergillus fumigatus. J. Fungi 2024, 10, 221. https://doi.org/10.3390/jof10030221
Emri T, Antal K, Varga K, Gila BC, Pócsi I. The Oxidative Stress Response Highly Depends on Glucose and Iron Availability in Aspergillus fumigatus. Journal of Fungi. 2024; 10(3):221. https://doi.org/10.3390/jof10030221
Chicago/Turabian StyleEmri, Tamás, Károly Antal, Kinga Varga, Barnabás Csaba Gila, and István Pócsi. 2024. "The Oxidative Stress Response Highly Depends on Glucose and Iron Availability in Aspergillus fumigatus" Journal of Fungi 10, no. 3: 221. https://doi.org/10.3390/jof10030221
APA StyleEmri, T., Antal, K., Varga, K., Gila, B. C., & Pócsi, I. (2024). The Oxidative Stress Response Highly Depends on Glucose and Iron Availability in Aspergillus fumigatus. Journal of Fungi, 10(3), 221. https://doi.org/10.3390/jof10030221