
Five New Species of the Genus Hymenogaster (Hymenogastraceae, Agaricales) from Northern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collections
2.2. DNA Extraction, PCR Amplification, Sequencing and Nucleotide Alignment
2.3. Phylogenetic Analysis
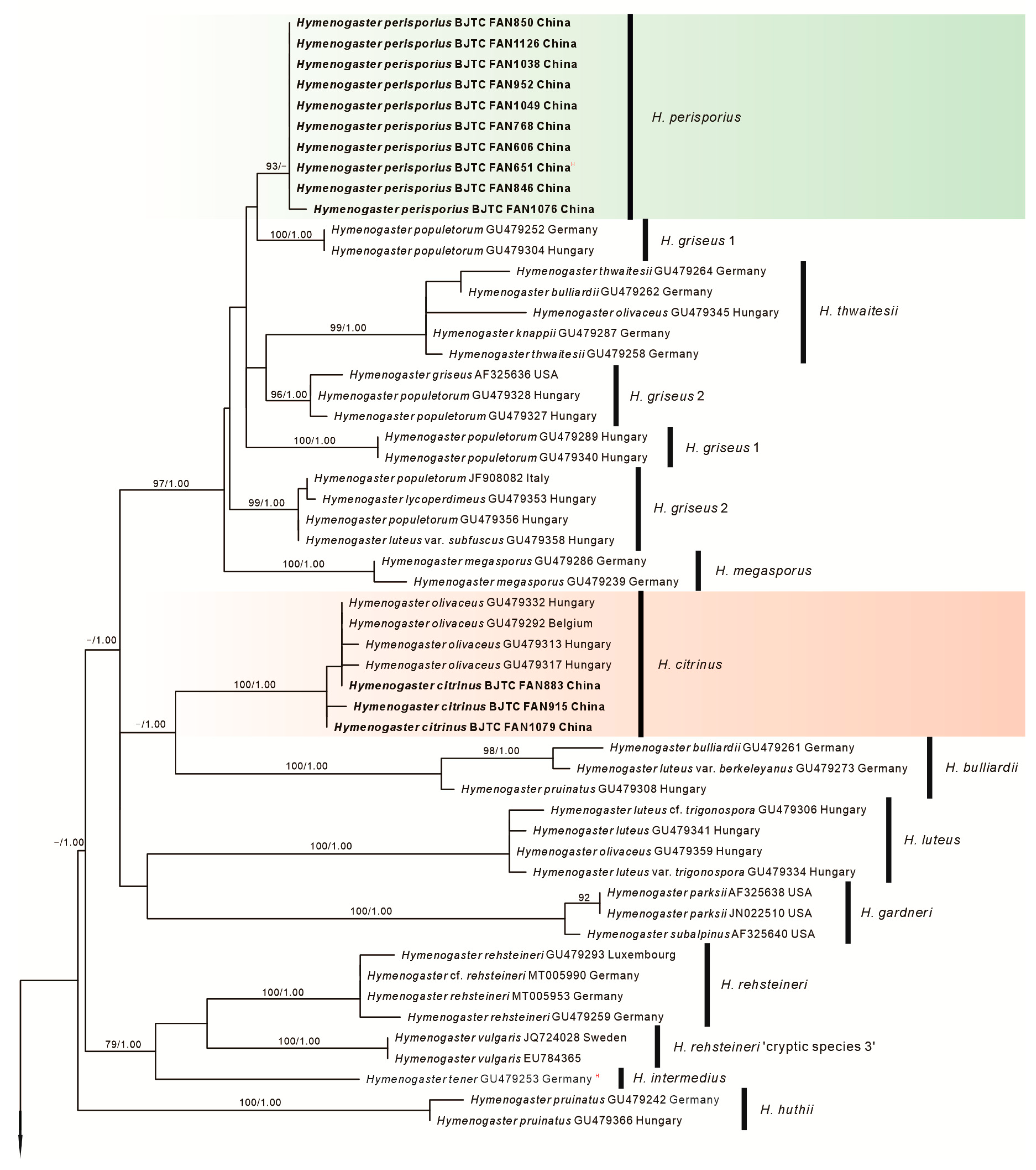
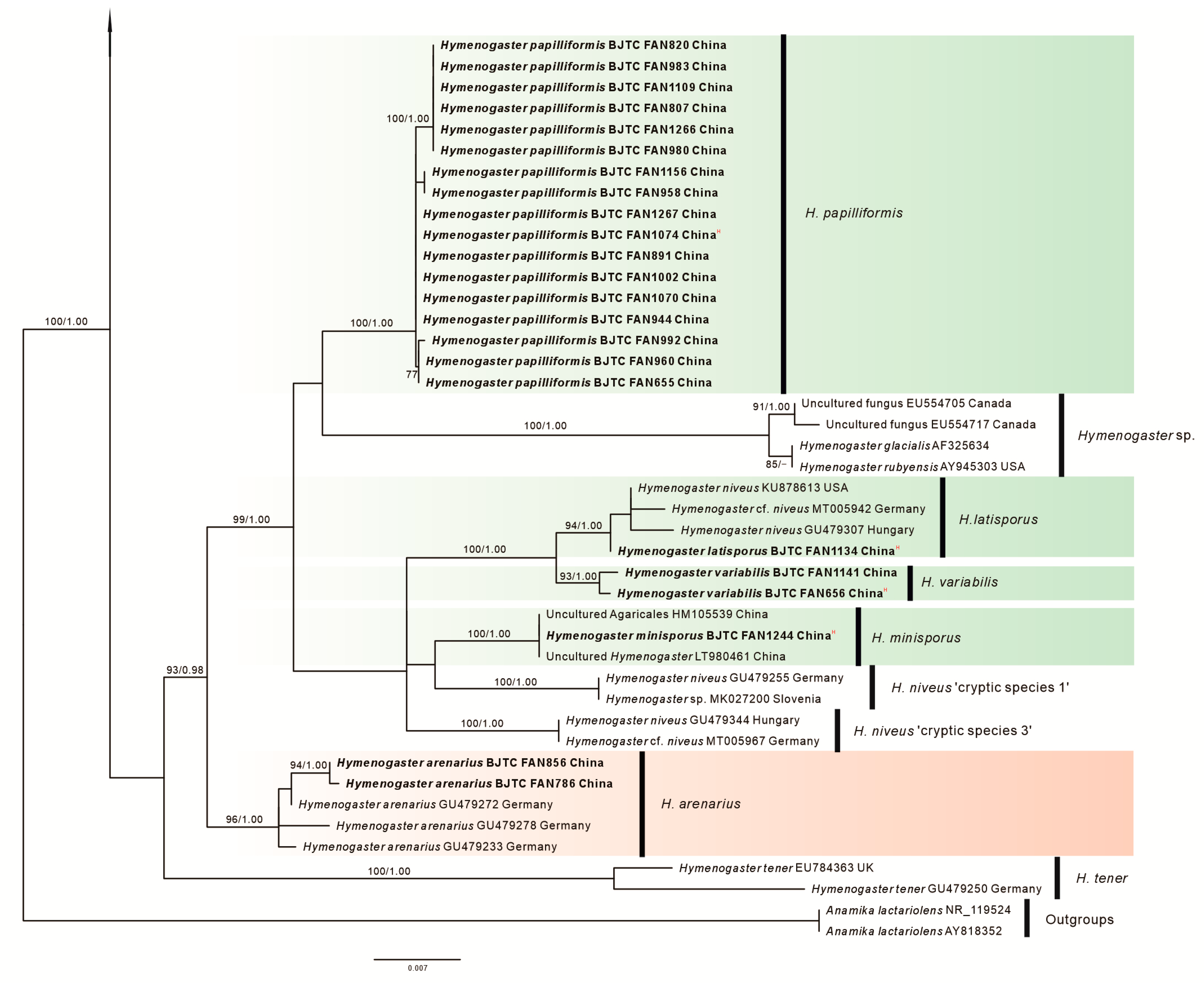
3. Results
3.1. Molecular Phylogenetics
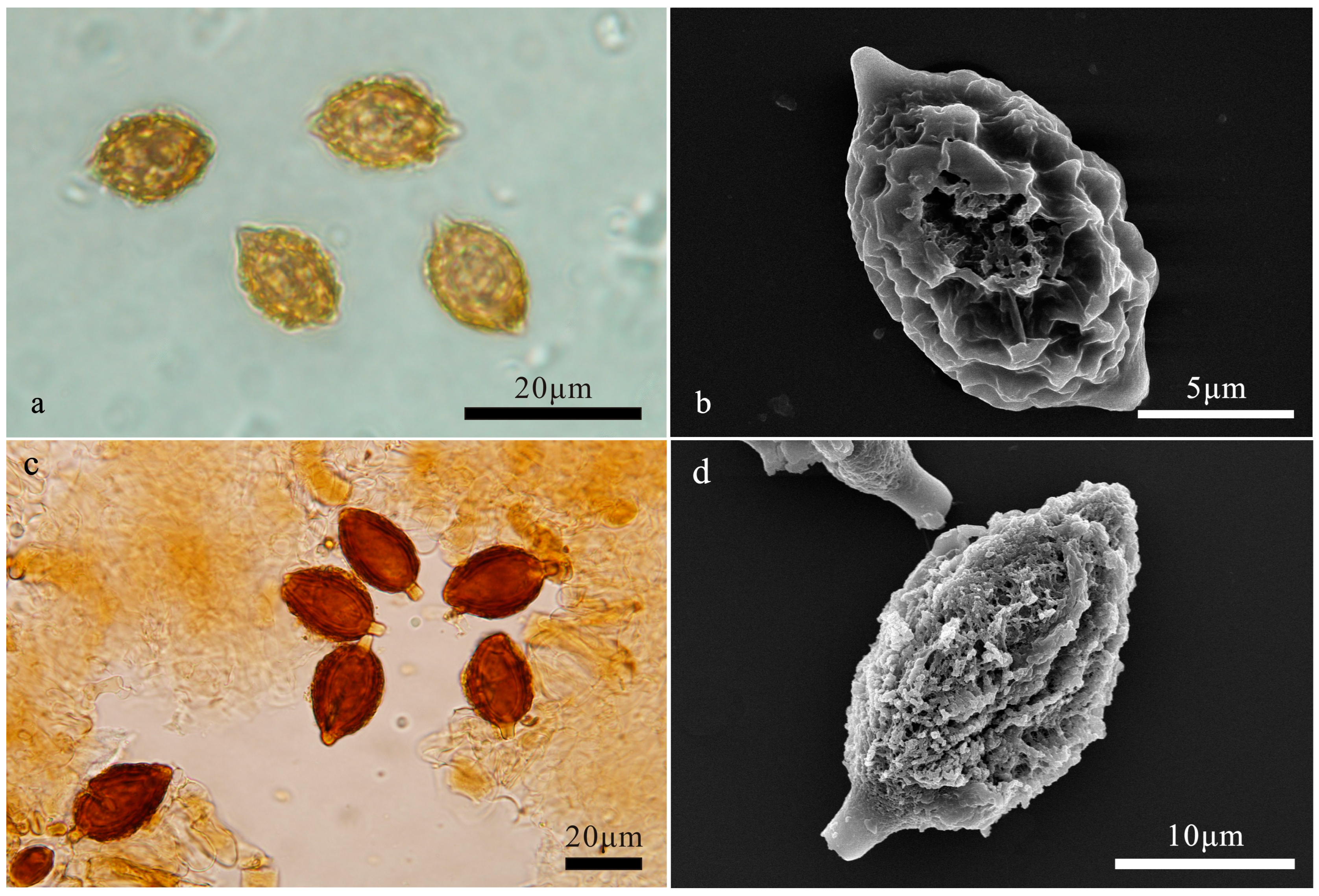
3.2. Taxonomy
4. Discussion
| 1 | Basidiome pale yellow, white to dirty white when fresh | 2 |
| 1 | Basidiome earth yellow to yellow-brown when fresh | 3 |
| 2 | Gleba reddish brown to brown when fresh | 4 |
| 2 | Gleba light brown when fresh | H. minisporus |
| 3 | Peridium very variable in thickness and more than >130 μm thick | 5 |
| 3 | Peridium without major changes in thickness and <130 μm thick | 6 |
| 4 | Basidiospores 21–26 × 14–18.5 μm | H. citrinus |
| 4 | Basidiospores 17–22 × 12–15 μm | H. perisporius |
| 5 | Basidiospores broad ellipsoidal to subglobose, Q = 1.1–1.3 | H. variabilis |
| 5 | Basidiospores broad fusiform to citriform, Q = 1.3–1.4 | H. apilliformis |
| 6 | Basidiospores fusiform, Q = 1.2–1.4 | H. arenarius |
| 6 | Basidiospores broadly ellipsoidal to broadly ovoid, Q = 1.1–1.3 | H. latisporus |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vittadini, C. Monographia Tuberacearum; Ex Typographia, F. Rusconi: Milian, Italy, 1831; pp. 1–88. [Google Scholar]
- Stielow, B.; Bratek, Z.; Orczán, A.K.I.; Rudnoy, S.; Hensel, G.; Hoffmann, P.; Klenk, H.P.; Göker, M. Species delimitation in taxonomically difficult fungi: The case of Hymenogaster. PLoS ONE 2011, 6, e15614. [Google Scholar] [CrossRef] [PubMed]
- Montecchi, A.; Sarasini, M. Funghi Ipogei d’Europa; AMB Fondazione Centro Studi Micologici: Vicenza, Italy, 2000; pp. 52–501. [Google Scholar]
- Pegler, D.N.; Spooner, B.M.; Young, T.W.K. British truffles: A revision of British Hypogeous fungi. Kew Bull. 1993, 49, 167. [Google Scholar]
- Dodge, C.W.; Zeller, S.M. Hymenogaster and related genera. Ann. Mo. Bot. Gard. 1934, 21, 625–708. [Google Scholar] [CrossRef]
- Harkness, H.W. Californian hypogaeous fungi. Calif. Acad. Sci. 1899, 1, 241–291. [Google Scholar]
- Smith, A.H. Notes on Dendrogaster, Gymnoglossum, Protoglossum and species of Hymenogaster. Mycologia 1966, 58, 100–124. [Google Scholar] [CrossRef]
- Cázares, E.; Trappe, J.M. Alpine and subalpine fungi of the Cascade Mountains. I. Hymenogaster glacialis sp. nov. Mycotaxon 1990, 38, 245–249. [Google Scholar]
- Fogel, R.; States, J. Materials for a hypogeous mycoflora of the Great Basin and adjacent cordilleras of the western United States VI: Hymenogaster rubyensis sp. nov. (Basidiomycota, Cortinariaceae). Mycotaxon 2001, 80, 333–337. [Google Scholar]
- Julou, T.; Burghardt, B.; Gebauer, G.; Berveiller, D.; Damesin, C.; Selosse, M.A. Mixotrophy in orchids: Insights from a comparative study of green individuals and nonphotosynthetic individuals of Cephalantera damasonium. N. Phytol. 2005, 166, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L.; Trappe, J.M. Diversity, Ecology, and Conservation of Truffle Fungi in Forests of the Pacific Northwest; Pacific Northwest Research Station, USDA Forest Service: Corvallis, OR, USA, 2009; pp. 1–194.
- Türkoğlu, A.; Castellano, M.A. New records of truffle fungi (Basidiomycetes) from Turkey. Turk. J. Bot. 2013, 37, 970–976. [Google Scholar] [CrossRef]
- Pecoraro, L.; Huang, L.; Caruso, T.; Perotto, S.; Girlandam, M.; Cai, L.; Liu, Z.J. Fungal diversity and specificity in Cephalanthera damasonium and C. longifolia (Orchidaceae) mycorrhizas. J. Syst. Evol. 2017, 55, 158–169. [Google Scholar] [CrossRef]
- Rudawska, M.; Kujawska, M.; Leski, T.; Janowski, D.; Karliński, L.; Wilgan, R. Ectomycorrhizal community structure of the admixture tree species Betula pendula, Carpinus betulus, and Tilia cordata grown in bare-root forest nurseries. For. Ecol. Manag. 2019, 437, 113–125. [Google Scholar] [CrossRef]
- Hawker, L.E. British hypogeous fungi. Philos. Trans. R. Soc. Biol. Sci. 1954, 237, 429–546. [Google Scholar]
- Soehner, V.E. Die Gattung Hymenogaster Vitt: Eine monographische Studie mit besonderer. Nova Hedwigia. 1962, 2, 1–113. [Google Scholar]
- Bougher, N.L.; Castellano, M.A. Delimitation of Hymenogaster sensu stricto and four new segregate genera. Mycologia 1993, 85, 273–293. [Google Scholar] [CrossRef]
- Smith, M.E.; Castellano, M.A.; Frank, J.L. Hymenogaster macmurphyi and Splanchnomyces behrii are sequestrate species of Xerocomellus from the western United States. Mycologia 2018, 110, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Y.H.; Liu, J.H. The hypogeous macrofungi of China. Edible Fungi China 2002, 21, 14–15. [Google Scholar]
- Dring, D.M. Techniques for microscopic preparation. In Methods in Microbiology; Booth, C., Ed.; Academic Press Inc.: New York, NY, USA, 1971; pp. 95–111. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, F.J.R.M. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, A Guide to Methods and Applications; Seliger, H., Gelfand, D.H., Sinsky, J.J., White, T.J., Eds.; Academic: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Frith, M.C. Adding unaligned sequences into an existing alignment using MAFFT and LAST. Bioinformatics 2012, 28, 3144–3146. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Estimating the rate of molecular evolution: Incorporating non-contemporaneous sequences into maximum likelihood phylogenies. Bioinformatics 2000, 16, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D.M. TreeView; Glasgow University: Glasgow, UK, 2001. [Google Scholar]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. MrModeltest 2.2. Computer Software Distributed by the University of Uppsala; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name in Analysis | Taxon Name | Collection | Country | GenBank Accession Number | |
|---|---|---|---|---|---|
| ITS | nrLSU | ||||
| Anamika lactariolens AY818352 | Anamika lactariolens | taxon:301353 | AY818352 | - | |
| Anamika lactariolens NR_119524 | Anamika lactariolens | HC 88/95 | NR_119524 | - | |
| Hymenogaster arenariusBJTC FAN786 China | Hymenogaster arenarius | BJTC FAN786 | China | PP467413 | PP467449 |
| Hymenogaster arenariusBJTC FAN856 China | Hymenogaster arenarius | BJTC FAN856 | China | PP467414 | PP467450 |
| Hymenogaster arenarius GU479233 Germany | Hymenogaster arenarius | it10_26_2 | Germany | GU479233 | - |
| Hymenogaster arenarius GU479272 Germany | Hymenogaster arenarius | it5_2 | Germany | GU479272 | - |
| Hymenogaster arenarius GU479278 Germany | Hymenogaster arenarius | it6_3 | Germany | GU479278 | - |
| Hymenogaster bulliardii GU479261 Germany | Hymenogaster bulliardii | it20_4 | Germany | GU479261 | - |
| Hymenogaster bulliardii GU479262 Germany | Hymenogaster thwaitesii | it20_4_1 | Germany | GU479262 | - |
| Hymenogaster cf. niveus MT005942 Germany | Hymenogaster xxx | KR-M-0044217 | Germany | MT005942 | - |
| Hymenogaster cf. niveus MT005967 Germany | Hymenogaster niveus ‘cryptic species 3’ | KR-M-0044314 | Germany | MT005967 | - |
| Hymenogaster cf. rehsteineri MT005990 Germany | Hymenogaster rehsteineri | KR-M-0044423 | Germany | MT005990 | - |
| Hymenogaster citrinusBJTC FAN1079 China | Hymenogaster citrinus | BJTC FAN1079 | China | PP467412 | PP467448 |
| Hymenogaster citrinusBJTC FAN883 China | Hymenogaster citrinus | BJTC FAN883 | China | PP467410 | PP467446 |
| Hymenogaster citrinusBJTC FAN915 China | Hymenogaster citrinus | BJTC FAN915 | China | PP467411 | PP467447 |
| Hymenogaster glacialis AF325634 | Hymenogaster sp. | GP 5302 | - | AF325634 | - |
| Hymenogaster griseus AF325636 USA | Hymenogaster griseus 2 | Trappe 12841 | USA | AF325636 | - |
| Hymenogaster knappii GU479287 Germany | Hymenogaster thwaitesii | it9_2 | Germany | GU479287 | - |
| Hymenogaster latisporusBJTC FAN1134 China | Hymenogaster latisporus | BJTC FAN1134, holotype | China | PP467404 | PP467440 |
| Hymenogaster luteus cf. trigonospora GU479306 Hungary | Hymenogaster luteus | zb1457 | Hungary | GU479306 | - |
| Hymenogaster luteus GU479341 Hungary | Hymenogaster luteus | zb2603 | Hungary | GU479341 | - |
| Hymenogaster luteus var. berkeleyanus GU479273 Germany | Hymenogaster bulliardii | it5_21 | Germany | GU479273 | - |
| Hymenogaster luteus var. subfuscus GU479358 Hungary | Hymenogaster griseus 2 | zb37 | Hungary | GU479358 | - |
| Hymenogaster luteus var. trigonospora GU479334 Hungary | Hymenogaster luteus | zb235 | Hungary | GU479334 | - |
| Hymenogaster lycoperdineus GU479353 Hungary | Hymenogaster griseus 2 | zb3533 | Hungary | GU479353 | - |
| Hymenogaster megasporus GU479239 Germany | Hymenogaster megasporus | it12_1 | Germany | GU479239 | - |
| Hymenogaster megasporus GU479286 Germany | Hymenogaster megasporus | it8_5_1 | Germany | GU479286 | - |
| Hymenogaster minisporusBJTC FAN1244 China | Hymenogaster minisporus | BJTC FAN1244, holotype | China | PP467407 | PP467443 |
| Hymenogaster niveus GU479255 Germany | Hymenogaster niveus ‘cryptic species 1’ | it17_3 | Germany | GU479255 | - |
| Hymenogaster niveus GU479307 Hungary | Hymenogaster xxx | zb1461 | Hungary | GU479307 | - |
| Hymenogaster niveus GU479344 Hungary | Hymenogaster niveus ‘cryptic species 3’ | zb28 | Hungary | GU479344 | - |
| Hymenogaster niveus KU878613 USA | Hymenogaster xxx | SC14_3 | USA | KU878613 | - |
| Hymenogaster olivaceus GU479292 Belgium | Hymenogaster citrinus | dt8293 | Belgium | GU479292 | - |
| Hymenogaster olivaceus GU479313 Hungary | Hymenogaster citrinus | zb1645 | Hungary | GU479313 | - |
| Hymenogaster olivaceus GU479317 Hungary | Hymenogaster citrinus | zb1817 | Hungary | GU479317 | - |
| Hymenogaster olivaceus GU479332 Hungary | Hymenogaster citrinus | zb2300 | Hungary | GU479332 | - |
| Hymenogaster olivaceus GU479345 Hungary | Hymenogaster thwaitesii | zb2804 | Hungary | GU479345 | - |
| Hymenogaster olivaceus GU479359 Hungary | Hymenogaster luteus | zb3721 | Hungary | GU479359 | - |
| Hymenogaster papilliformisBJTC FAN1002 China | Hymenogaster papilliformis | BJTC FAN1002 | China | PP467396 | PP467432 |
| Hymenogaster papilliformisBJTC FAN1070 China | Hymenogaster papilliformis | BJTC FAN1070 | China | PP467399 | PP467435 |
| Hymenogaster papilliformisBJTC FAN1074 China | Hymenogaster papilliformis | BJTC FAN1074, holotype | China | PP467400 | PP467436 |
| Hymenogaster papilliformisBJTC FAN1109 China | Hymenogaster papilliformis | BJTC FAN1109 | China | PP467402 | PP467438 |
| Hymenogaster papilliformisBJTC FAN1156 China | Hymenogaster papilliformis | BJTC FAN1156 | China | PP467406 | PP467442 |
| Hymenogaster papilliformisBJTC FAN1266 China | Hymenogaster papilliformis | BJTC FAN1266 | China | PP467408 | PP467444 |
| Hymenogaster papilliformisBJTC FAN1267 China | Hymenogaster papilliformis | BJTC FAN1267 | China | PP467409 | PP467445 |
| Hymenogaster papilliformisBJTC FAN655 China | Hymenogaster papilliformis | BJTC FAN655 | China | PP467381 | PP467417 |
| Hymenogaster papilliformisBJTC FAN807 China | Hymenogaster papilliformis | BJTC FAN807 | China | PP467384 | PP467420 |
| Hymenogaster papilliformisBJTC FAN820 China | Hymenogaster papilliformis | BJTC FAN820 | China | PP467385 | PP467421 |
| Hymenogaster papilliformisBJTC FAN891 China | Hymenogaster papilliformis | BJTC FAN891 | China | PP467388 | PP467424 |
| Hymenogaster papilliformisBJTC FAN944 China | Hymenogaster papilliformis | BJTC FAN944 | China | PP467389 | PP467425 |
| Hymenogaster papilliformisBJTC FAN958 China | Hymenogaster papilliformis | BJTC FAN958 | China | PP467391 | PP467427 |
| Hymenogaster papilliformisBJTC FAN960 China | Hymenogaster papilliformis | BJTC FAN960 | China | PP467392 | PP467428 |
| Hymenogaster papilliformisBJTC FAN980 China | Hymenogaster papilliformis | BJTC FAN980 | China | PP467393 | PP467429 |
| Hymenogaster papilliformisBJTC FAN983 China | Hymenogaster papilliformis | BJTC FAN983 | China | PP467394 | PP467430 |
| Hymenogaster papilliformisBJTC FAN992 China | Hymenogaster papilliformis | BJTC FAN992 | China | PP467395 | PP467431 |
| Hymenogaster parksii AF325638 USA | Hymenogaster gardneri | Trappe 13296 | USA | AF325638 | - |
| Hymenogaster parksii JN022510 USA | Hymenogaster gardneri | SOC1643 | USA | JN022510 | - |
| Hymenogaster perisporiusBJTC FAN1038 China | Hymenogaster perisporius | BJTC FAN1038 | China | PP467397 | PP467433 |
| Hymenogaster perisporiusBJTC FAN1049 China | Hymenogaster perisporius | BJTC FAN1049 | China | PP467398 | PP467434 |
| Hymenogaster perisporiusBJTC FAN1076 China | Hymenogaster perisporius | BJTC FAN1076 | China | PP467401 | PP467437 |
| Hymenogaster perisporiusBJTC FAN1126 China | Hymenogaster perisporius | BJTC FAN1126 | China | PP467403 | PP467439 |
| Hymenogaster perisporiusBJTC FAN606 China | Hymenogaster perisporius | BJTC FAN606 | China | PP467379 | PP467415 |
| Hymenogaster perisporiusBJTC FAN651 China | Hymenogaster perisporius | BJTC FAN651, holotype | China | PP467380 | PP467416 |
| Hymenogaster perisporiusBJTC FAN768 China | Hymenogaster perisporius | BJTC FAN768 | China | PP467383 | PP467419 |
| Hymenogaster perisporiusBJTC FAN846 China | Hymenogaster perisporius | BJTC FAN846 | China | PP467386 | PP467422 |
| Hymenogaster perisporiusBJTC FAN850 China | Hymenogaster perisporius | BJTC FAN850 | China | PP467387 | PP467423 |
| Hymenogaster perisporiusBJTC FAN952 China | Hymenogaster perisporius | BJTC FAN952 | China | PP467390 | PP467426 |
| Hymenogaster populetorum GU479252 Germany | Hymenogaster griseus 1 | it16_1_1 | Germany | GU479252 | - |
| Hymenogaster populetorum GU479289 Hungary | Hymenogaster griseus 1 | aszodvt_1991 | Hungary | GU479289 | - |
| Hymenogaster populetorum GU479304 Hungary | Hymenogaster griseus 1 | zb1436 | Hungary | GU479304 | - |
| Hymenogaster populetorum GU479327 Hungary | Hymenogaster griseus 2 | zb2097 | Hungary | GU479327 | - |
| Hymenogaster populetorum GU479328 Hungary | Hymenogaster griseus 2 | zb2105 | Hungary | GU479328 | - |
| Hymenogaster populetorum GU479340 Hungary | Hymenogaster griseus 1 | zb2576 | Hungary | GU479340 | - |
| Hymenogaster populetorum GU479356 Hungary | Hymenogaster griseus 2 | zb3594 | Hungary | GU479356 | - |
| Hymenogaster populetorum JF908082 Italy | Hymenogaster griseus 2 | 17022 | Italy | JF908082 | - |
| Hymenogaster pruinatus GU479242 Germany | Hymenogaster huthii | it12_3_1 | Germany | GU479242 | - |
| Hymenogaster pruinatus GU479308 Hungary | Hymenogaster bulliardii | zb1485 | Hungary | GU479308 | - |
| Hymenogaster pruinatus GU479366 Hungary | Hymenogaster huthii | zb95 | Hungary | GU479366 | - |
| Hymenogaster rehsteineri GU479259 Germany | Hymenogaster rehsteineri | it2_4_1 | Germany | GU479259 | - |
| Hymenogaster rehsteineri GU479293 Luxembourg | Hymenogaster rehsteineri | dt8455 | Luxembourg | GU479293 | - |
| Hymenogaster rehsteineri MT005953 Germany | Hymenogaster rehsteineri | KR-M-0044018 | Germany | MT005953 | - |
| Hymenogaster rubyensis AY945303 USA | Hymenogaster sp. | Fogel 2698 | USA | AY945303 | - |
| Hymenogaster sp. MK027200 Slovenia | Hymenogaster niveus ‘cryptic species 1’ | FV4_04 | Slovenia | MK027200 | - |
| Hymenogaster subalpinus AF325640 USA | Hymenogaster gardneri | Trappe 22752 | USA | AF325640 | - |
| Hymenogaster tener EU784363 UK | Hymenogaster tener | RBG Kew K(M)102406 | UK | EU784363 | - |
| Hymenogaster tener GU479250 Germany | Hymenogaster tener | it15_3 | Germany | GU479250 | - |
| Hymenogaster tener GU479253 Germany | Hymenogaster intermedius | it16_2, holotype | Germany | GU479253 | - |
| Hymenogaster thwaitesii GU479258 Germany | Hymenogaster thwaitesii | it2_2 | Germany | GU479258 | - |
| Hymenogaster thwaitesii GU479264 Germany | Hymenogaster thwaitesii | it3_2 | Germany | GU479264 | - |
| Hymenogaster variabilisBJTC FAN1141 China | Hymenogaster variabilis | BJTC FAN1141 | China | PP467405 | PP467441 |
| Hymenogaster variabilisBJTC FAN656 China | Hymenogaster variabilis | BJTC FAN656, holotype | China | PP467382 | PP467418 |
| Hymenogaster vulgaris EU784365 | Hymenogaster rehsteineri ‘cryptic species 3’ | RBG Kew K(M)27363 | - | EU784365 | - |
| Hymenogaster vulgaris JQ724028 Sweden | Hymenogaster rehsteineri ‘cryptic species 3’ | GN_4d_I | Sweden | JQ724028 | - |
| Uncultured Agaricales HM105539 China | Hymenogaster minisporus | QL054 | China | HM105539 | - |
| Uncultured fungus EU554705 Canada | Hymenogaster sp. | A2N_88 | Canada | EU554705 | - |
| Uncultured fungus EU554717 Canada | Hymenogaster sp. | A3E_60 | Canada | EU554717 | - |
| Uncultured Hymenogaster LT980461 China | Hymenogaster minisporus | taxon:522720 | China | LT980461 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Mao, N.; Fu, H.; Zhang, Y.; Fan, L. Five New Species of the Genus Hymenogaster (Hymenogastraceae, Agaricales) from Northern China. J. Fungi 2024, 10, 272. https://doi.org/10.3390/jof10040272
Li T, Mao N, Fu H, Zhang Y, Fan L. Five New Species of the Genus Hymenogaster (Hymenogastraceae, Agaricales) from Northern China. Journal of Fungi. 2024; 10(4):272. https://doi.org/10.3390/jof10040272
Chicago/Turabian StyleLi, Ting, Ning Mao, Haoyu Fu, Yuxin Zhang, and Li Fan. 2024. "Five New Species of the Genus Hymenogaster (Hymenogastraceae, Agaricales) from Northern China" Journal of Fungi 10, no. 4: 272. https://doi.org/10.3390/jof10040272
APA StyleLi, T., Mao, N., Fu, H., Zhang, Y., & Fan, L. (2024). Five New Species of the Genus Hymenogaster (Hymenogastraceae, Agaricales) from Northern China. Journal of Fungi, 10(4), 272. https://doi.org/10.3390/jof10040272