
Host–Pathogen Interactions and Correlated Factors That Are Affected in Replicative-Aged Cryptococcus neoformans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Isolation of Old C. neoformans Cells
2.2. Macrophage Cell Line Culture
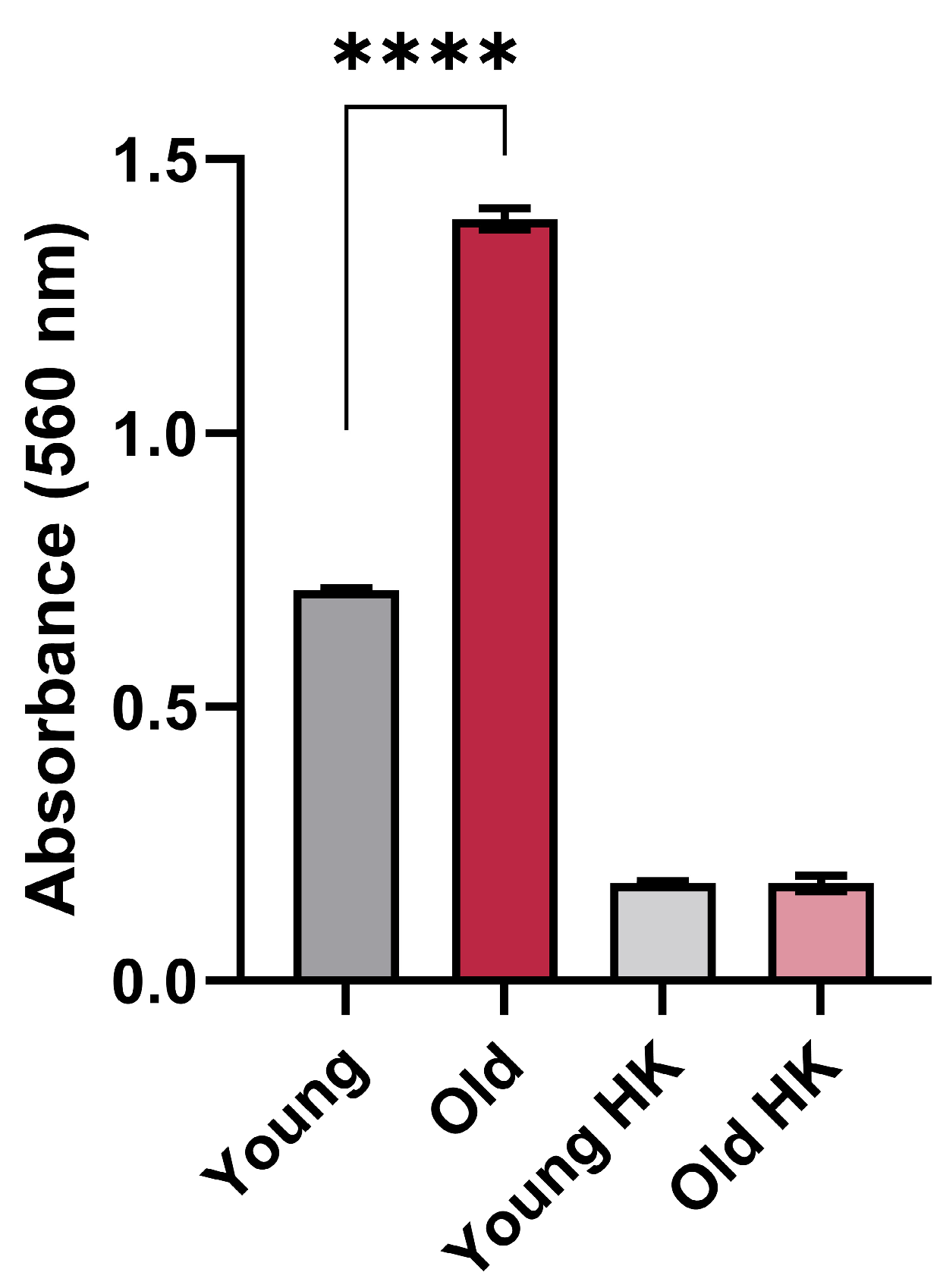
2.3. Urease Activity
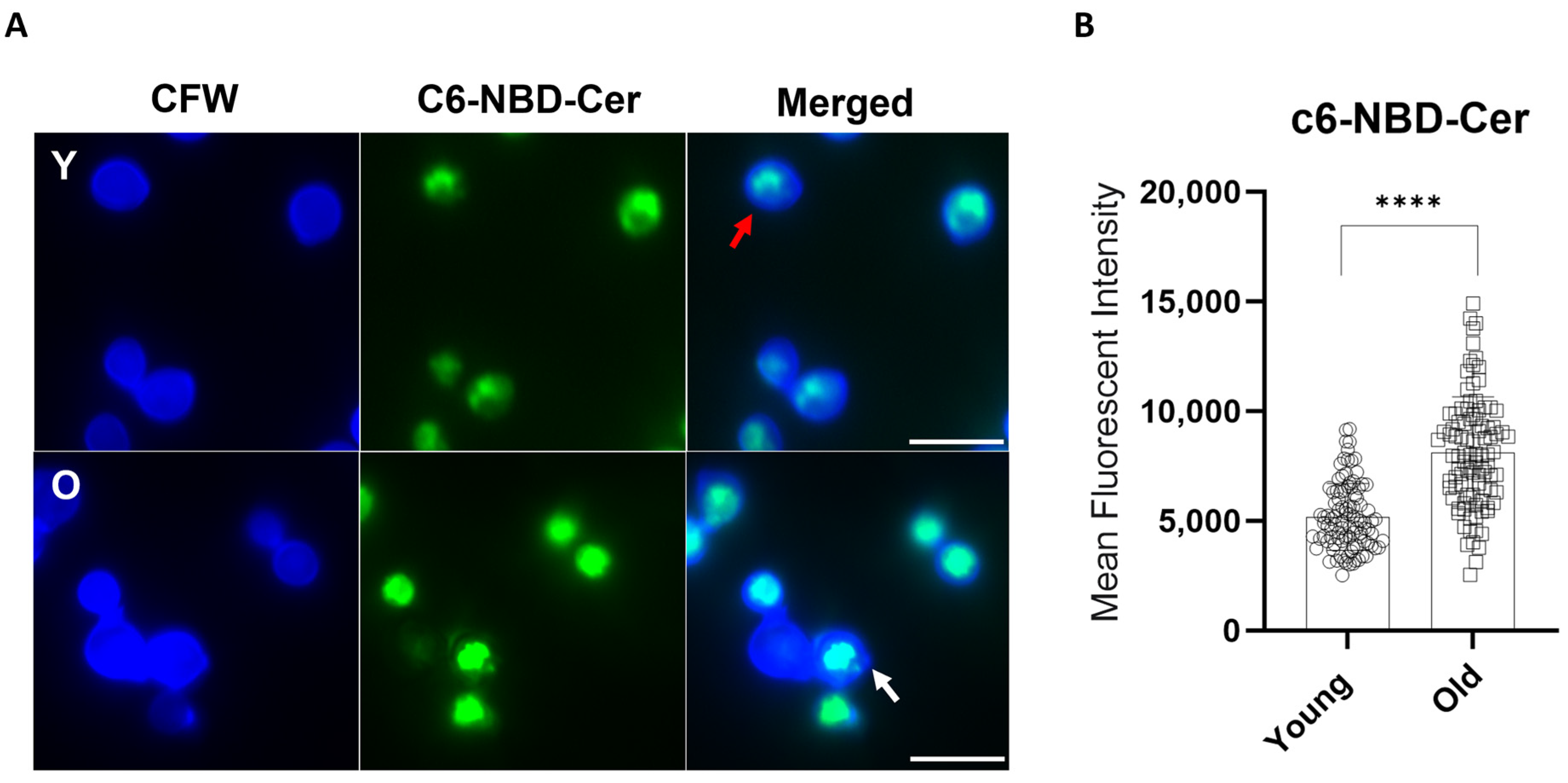
2.4. Analysis of the Golgi Apparatus
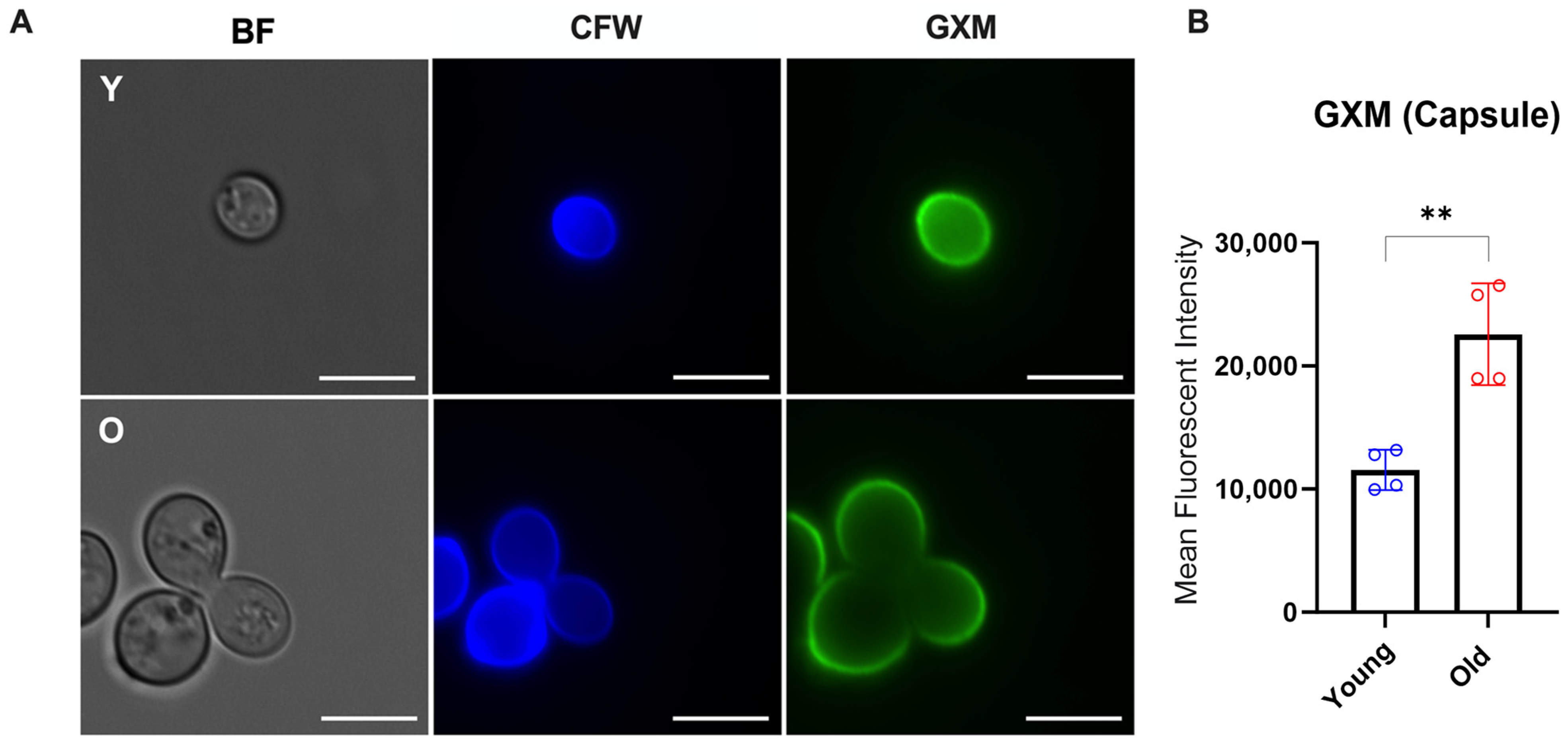
2.5. Polysaccharide Capsule Analysis
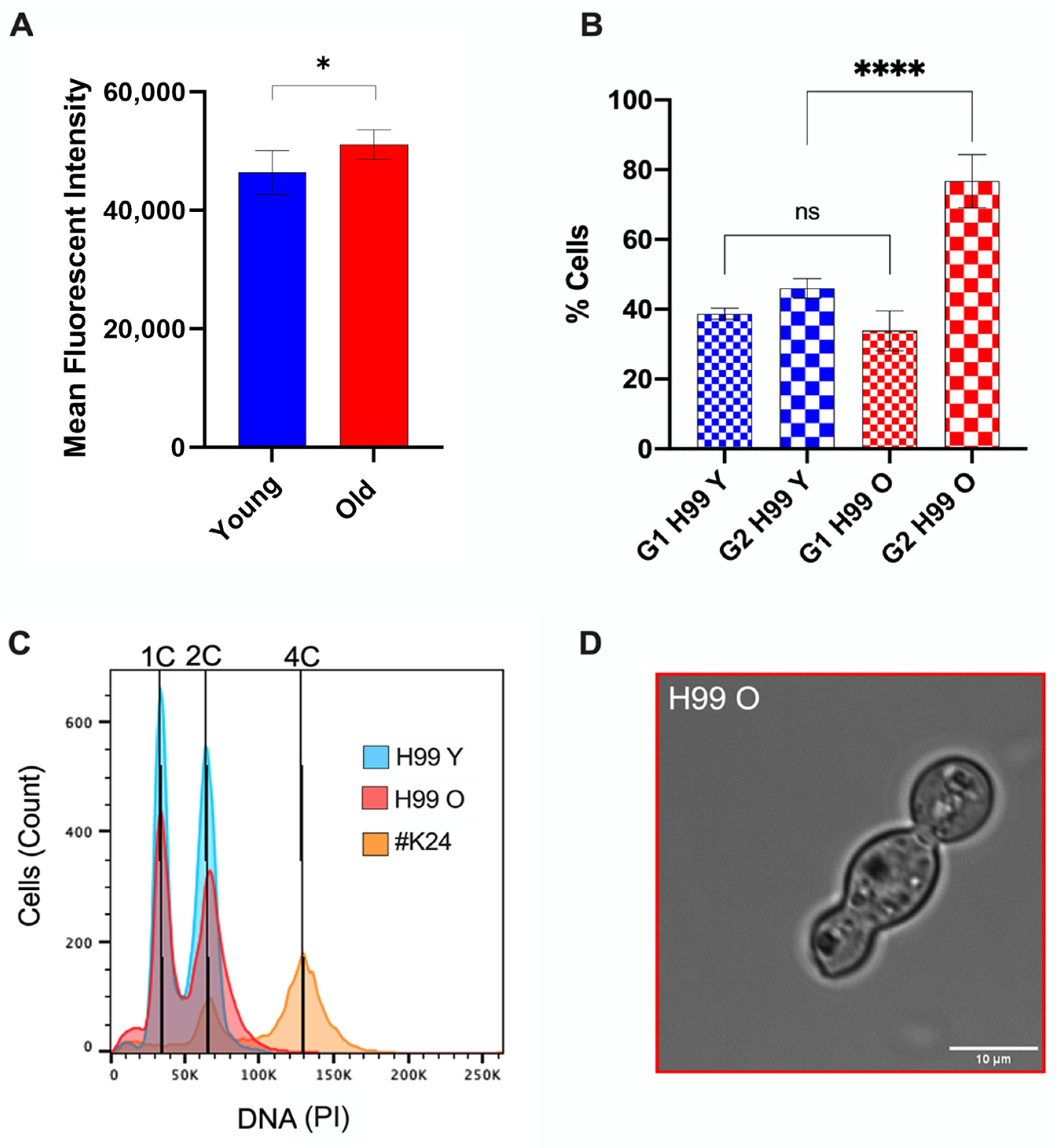
2.6. DNA Staining and Cell Cycle Analysis
2.7. Phagocytosis Assay, Phagosomal Acidification, and Phagosomal Permeability
2.8. Time-Lapse Microscopy and Vomocytosis Rate
2.9. Data Analysis
3. Results
3.1. Effects of Aging on Urease Activity in C. neoformans
3.2. Longevity in C. neoformans Affects Golgi Apparatus and Capsular Properties
3.3. Old C. neoformans Presented an Increase in DNA Content and G2 Arrest
3.4. Vomocytosis Phenomenon during Replicative Aging in C. neoformans
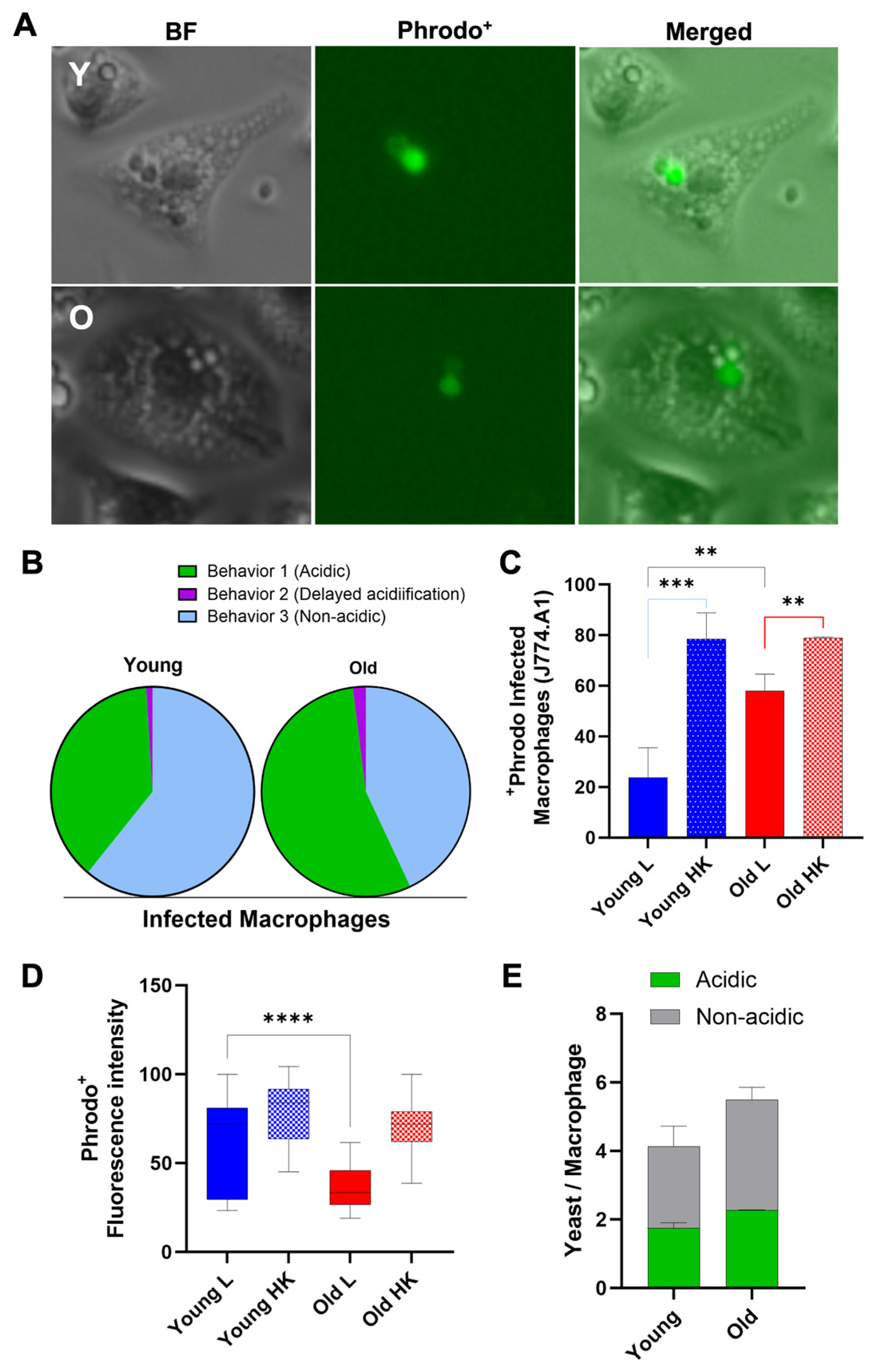
3.5. Old C. neoformans Prefer to Reside in Acidified Phagosomes
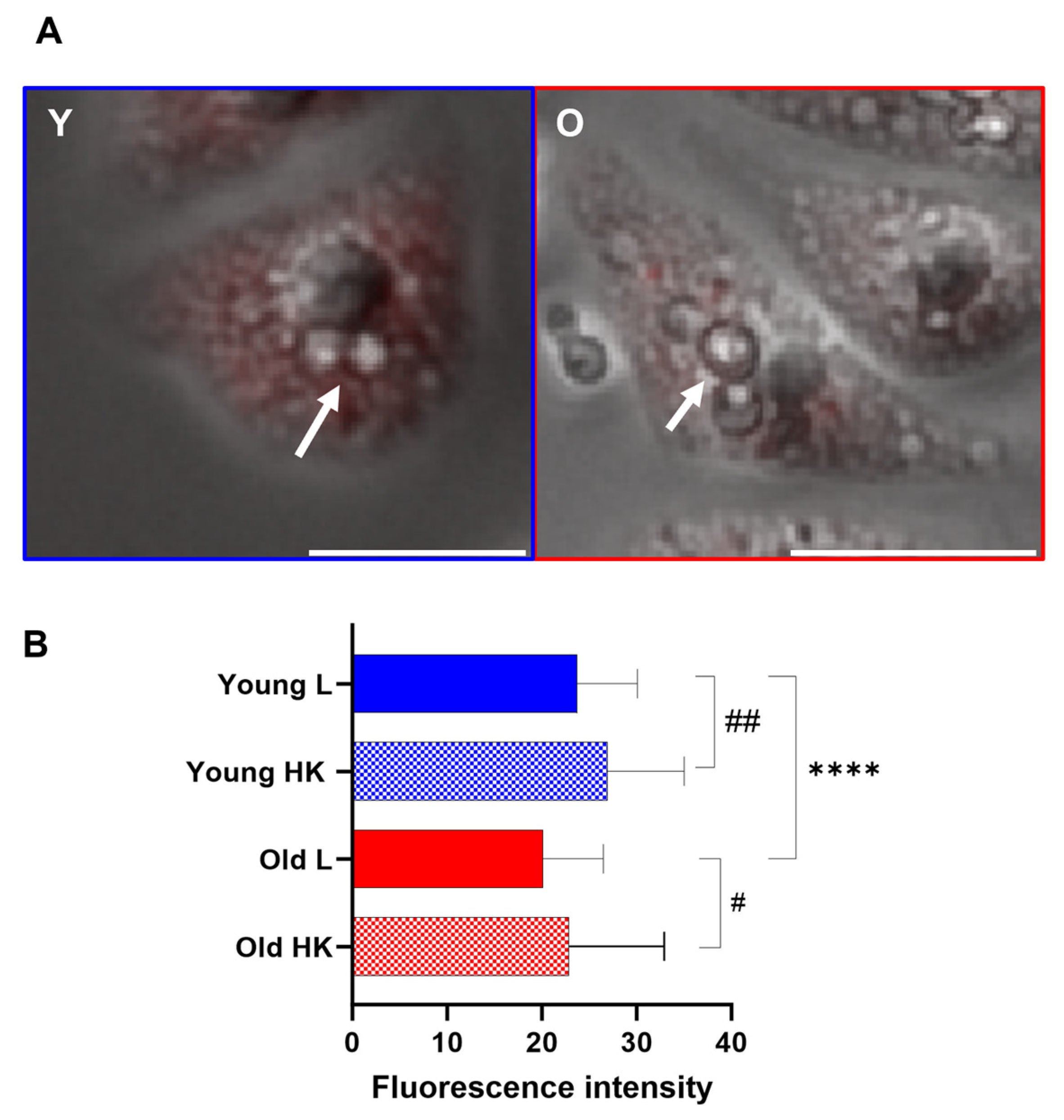
3.6. Generational Age in C. neoformans Influences Lysosomal Permeabilization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casadevall, A.; Fang, F.C. The intracellular pathogen concept. Mol. Microbiol. 2020, 113, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.S.; Seoane, P.I.; Sephton-Clark, P.; Bojarczuk, A.; Hotham, R.; Giurisato, E.; Sarhan, A.R.; Hillen, A.; Velde, G.V.; Gray, N.S.; et al. Vomocytosis of live pathogens from macrophages is regulated by the atypical MAP kinase ERK5. Sci. Adv. 2017, 3, e1700898. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, Z.W.; Strickland, A.B.; Shi, M. Cryptococcus neoformans Infection in the Central Nervous System: The Battle between Host and Pathogen. J. Fungi 2022, 8, 1069. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nosanchuk, J.D. Recognition of fungal priority pathogens: What next? PLoS Negl. Trop. Dis. 2023, 17, e0011136. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Denning, D.W. The WHO fungal priority pathogens list as a game-changer. Nat. Rev. Microbiol. 2023, 21, 211–212. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Cook, E.; Xess, I.; Hasan, F.; Fries, D.; Fries, B.C. Isolation and Characterization of Senescent Cryptococcus neoformans and Implications for Phenotypic Switching and Pathogenesis in Chronic Cryptococcosis. Eukaryot. Cell 2009, 8, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Pechuan, X.; Goldman, D.L.; Edelman, B.; Bergman, A.; Fries, B.C. Old Cryptococcus neoformans Cells Contribute to Virulence in Chronic Cryptococcosis. mBio 2013, 4, e00455-13. [Google Scholar] [CrossRef] [PubMed]
- Orner, E.P.; Zhang, P.; Jo, M.C.; Bhattacharya, S.; Qin, L.; Fries, B.C. High-Throughput Yeast Aging Analysis for Cryptococcus (HYAAC) microfluidic device streamlines aging studies in Cryptococcus neoformans. Commun. Biol. 2019, 2, 256. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.K.A.; Bhattacharya, S.; Oliveira, N.K.; Savitt, A.G.; Zamith-Miranda, D.; Nosanchuk, J.D.; Fries, B.C. Replicative Aging Remodels the Cell Wall and Is Associated with Increased Intracellular Trafficking in Human Pathogenic Yeasts. mBio 2021, 13, e0019022. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Casadevall, A. Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.S.; Coelho, C.; De Leon-Rodriguez, C.M.; Rossi, D.C.P.; Camacho, E.; Jung, E.H.; Kulkarni, M.; Casadevall, A. Cryptococcus neoformans urease affects the outcome of intracellular pathogenesis by modulating phagolysosomal pH. PLoS Pathog. 2018, 14, e1007144. [Google Scholar] [CrossRef] [PubMed]
- De Leon-Rodriguez, C.M.; Fu, M.S.; Çorbali, M.O.; Cordero, R.J.B.; Casadevall, A. The Capsule of Cryptococcus neoformans Modulates Phagosomal pH through Its Acid-Base Properties. mSphere 2018, 3, e00437-18. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, S.; Li, Z.; Fu, M.S.; Ding, M.; Fulton, S.R.; Yoder, J.M.; Tran, V.; Nielsen, K. The Cyclin Cln1 Controls Polyploid Titan Cell Formation following a Stress-Induced G(2) Arrest in Cryptococcus. mBio 2021, 12, e0250921. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Burgos, E.J.; Stuckey, P.V.; Santiago-Tirado, F.H. Real-time visualization of phagosomal pH manipulation by Cryptococcus neoformans in an immune signal-dependent way. Front. Cell Infect. Microbiol. 2022, 12, 967486. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Diago-Navarro, E.; Wang, X.; Fenster, M.; Fries, B.C. Characterization of the virulence of Cryptococcus neoformans strains in an insect model. Virulence 2015, 6, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.D.; Horstmeier, C.D.; Land, G.A.; Foxworth, J.H. Rapid urea broth test for yeasts. J. Clin. Microbiol. 1978, 7, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.J.; Akyel, A.; Morasko, V.J.; Gerlach, R.; Phillips, A.J. Temperature-dependent inactivation and catalysis rates of plant-based ureases for engineered biomineralization. Eng. Rep. 2021, 3, e12299. [Google Scholar] [CrossRef]
- Kmetzsch, L.; Joffe, L.S.; Staats, C.C.; de Oliveira, D.L.; Fonseca, F.L.; Cordero, R.J.; Casadevall, A.; Nimrichter, L.; Schrank, A.; Vainstein, M.H.; et al. Role for Golgi reassembly and stacking protein (GRASP) in polysaccharide secretion and fungal virulence. Mol. Microbiol. 2011, 81, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Oliveira, D.L.; Joffe, L.S.; Hu, G.; Gazos-Lopes, F.; Fonseca, F.L.; Almeida, I.C.; Frases, S.; Kronstad, J.W.; Rodrigues, M.L. Role of the Apt1 protein in polysaccharide secretion by Cryptococcus neoformans. Eukaryot. Cell 2014, 13, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Cleare, W.; Feldmesser, M.; Glatman-Freedman, A.; Goldman, D.L.; Kozel, T.R.; Lendvai, N.; Mukherjee, J.; Pirofski, L.A.; Rivera, J.; et al. Characterization of a murine monoclonal antibody to Cryptococcus neoformans polysaccharide that is a candidate for human therapeutic studies. Antimicrob. Agents Chemother. 1998, 42, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Ramos, C.L.; Frases, S.; Godinho, R.M.d.C.; Fonseca, F.L.; Rodrigues, M.L. Lack of chitin synthase genes impacts capsular architecture and cellular physiology in Cryptococcus neoformans. Cell Surf. 2018, 2, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.T.; Braverman, A.L.; Selmecki, A. Flow Cytometry Analysis of Fungal Ploidy. Curr. Protoc. Microbiol. 2018, 50, e58. [Google Scholar] [CrossRef] [PubMed]
- Samantaray, S.; Correia, J.N.; Garelnabi, M.; Voelz, K.; May, R.C.; Hall, R.A. Novel cell-based in vitro screen to identify small-molecule inhibitors against intracellular replication of Cryptococcus neoformans in macrophages. Int. J. Antimicrob. Agents 2016, 48, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Jamil, K.; Polyak, M.J.; Feehan, D.D.; Surmanowicz, P.; Stack, D.; Li, S.S.; Ogbomo, H.; Olszewski, M.; Ganguly, A.; Mody, C.H. Phagosomal F-Actin Retention by Cryptococcus gattii Induces Dendritic Cell Immunoparalysis. mBio 2020, 11, e01821-20. [Google Scholar] [CrossRef]
- Yoneda, A.; Doering, T.L. A eukaryotic capsular polysaccharide is synthesized intracellularly and secreted via exocytosis. Mol. Biol. Cell 2006, 17, 5131–5140. [Google Scholar] [CrossRef] [PubMed]
- García-Rodas, R.; Cordero, R.J.; Trevijano-Contador, N.; Janbon, G.; Moyrand, F.; Casadevall, A.; Zaragoza, O. Capsule growth in Cryptococcus neoformans is coordinated with cell cycle progression. mBio 2014, 5, e00945-14. [Google Scholar] [CrossRef]
- Stukes, S.; Casadevall, A. Visualizing non-lytic exocytosis of Cryptococcus neoformans from macrophages using digital light microscopy. J. Vis. Exp. 2014, 92, e52084. [Google Scholar] [CrossRef]
- Dragotakes, Q.; Jacobs, E.; Ramirez, L.S.; Yoon, O.I.; Perez-Stable, C.; Eden, H.; Pagnotta, J.; Vij, R.; Bergman, A.; D’Alessio, F.; et al. Bet-hedging antimicrobial strategies in macrophage phagosome acidification drive the dynamics of Cryptococcus neoformans intracellular escape mechanisms. PLoS Pathog. 2022, 18, e1010697. [Google Scholar] [CrossRef] [PubMed]
- Dragotakes, Q.; Stouffer, K.M.; Fu, M.S.; Sella, Y.; Youn, C.; Yoon, O.I.; De Leon-Rodriguez, C.M.; Freij, J.B.; Bergman, A.; Casadevall, A. Macrophages use a bet-hedging strategy for antimicrobial activity in phagolysosomal acidification. J. Clin. Investig. 2020, 130, 3805–3819. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.J.; Eastman, A.J.; Qiu, Y.; Gregorka, B.; Kozel, T.R.; Osterholzer, J.J.; Curtis, J.L.; Swanson, J.A.; Olszewski, M.A. Cryptococcus neoformans-induced macrophage lysosome damage crucially contributes to fungal virulence. J. Immunol. 2015, 194, 2219–2231. [Google Scholar] [CrossRef] [PubMed]
- Gaylord, E.A.; Choy, H.L.; Doering, T.L. Dangerous Liaisons: Interactions of Cryptococcus neoformans with Host Phagocytes. Pathogens 2020, 9, 891. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, K.; Love, J.C.; Ploegh, H.L.; Vyas, J.M. Recruitment of CD63 to Cryptococcus neoformans phagosomes requires acidification. Proc. Natl. Acad. Sci. USA 2006, 103, 15945–15950. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.P.; Casadevall, A. Reciprocal modulation of ammonia and melanin production has implications for cryptococcal virulence. Nat. Commun. 2023, 14, 849. [Google Scholar] [CrossRef] [PubMed]
- Orner, E.P.; Bhattacharya, S.; Kalenja, K.; Hayden, D.; Del Poeta, M.; Fries, B.C. Cell Wall-Associated Virulence Factors Contribute to Increased Resilience of Old Cryptococcus neoformans Cells. Front. Microbiol. 2019, 10, 2513. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Jain, N.; Fries, B.C. Modulation of Replicative Lifespan in Cryptococcus neoformans: Implications for Virulence. Front. Microbiol. 2017, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Alonso-Crisóstomo, L.; Székely, T., Jr.; Diago-Navarro, E.; Orner, E.P.; Smith, K.; Munshi, M.A.; Del Poeta, M.; Balázsi, G.; Fries, B.C. Generational distribution of a Candida glabrata population: Resilient old cells prevail, while younger cells dominate in the vulnerable host. PLoS Pathog. 2017, 13, e1006355. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.M.; Levitz, S.M. Chloroquine antagonizes the proinflammatory cytokine response to opportunistic fungi by alkalizing the fungal phagolysosome. J. Infect. Dis. 2001, 183, 935–942. [Google Scholar] [CrossRef]
- Hommel, B.; Sturny-Leclère, A.; Volant, S.; Veluppillai, N.; Duchateau, M.; Yu, C.-H.; Hourdel, V.; Varet, H.; Matondo, M.; Perfect, J.R.; et al. Cryptococcus neoformans resists to drastic conditions by switching to viable but non-culturable cell phenotype. PLoS Pathog. 2019, 15, e1007945. [Google Scholar] [CrossRef]
- Tucker, S.C.; Casadevall, A. Replication of Cryptococcus neoformans in macrophages is accompanied by phagosomal permeabilization and accumulation of vesicles containing polysaccharide in the cytoplasm. Proc. Natl. Acad. Sci. USA 2002, 99, 3165–3170. [Google Scholar] [CrossRef]
- Fu, J.; Morris, I.R.; Wickes, B.L. The Production of Monokaryotic Hyphae by Cryptococcus neoformans Can Be Induced by High Temperature Arrest of the Cell Cycle and Is Independent of Same-Sex Mating. PLoS Pathog. 2013, 9, e1003335. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.D.; Hashemi, J.; Bibi, M.; Pulver, R.; Bavli, D.; Nahmias, Y.; Wellington, M.; Sapiro, G.; Berman, J. A tetraploid intermediate precedes aneuploid formation in yeasts exposed to fluconazole. PLoS Biol. 2014, 12, e1001815. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Acuña, M.; Pacifici, N.; Lewis, J.S. Vomocytosis: Too Much Booze, Base, or Calcium? mBio 2019, 10, e02526-19. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Golgi Morphology | Young | Old |
|---|---|---|
| Peripherical | 62.85% | 60% |
| Central | 37.15% | 40% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.K.A.; Min, S.; Yoo, K.; Fries, B.C. Host–Pathogen Interactions and Correlated Factors That Are Affected in Replicative-Aged Cryptococcus neoformans. J. Fungi 2024, 10, 279. https://doi.org/10.3390/jof10040279
Silva VKA, Min S, Yoo K, Fries BC. Host–Pathogen Interactions and Correlated Factors That Are Affected in Replicative-Aged Cryptococcus neoformans. Journal of Fungi. 2024; 10(4):279. https://doi.org/10.3390/jof10040279
Chicago/Turabian StyleSilva, Vanessa K. A., Sungyun Min, Kyungyoon Yoo, and Bettina C. Fries. 2024. "Host–Pathogen Interactions and Correlated Factors That Are Affected in Replicative-Aged Cryptococcus neoformans" Journal of Fungi 10, no. 4: 279. https://doi.org/10.3390/jof10040279
APA StyleSilva, V. K. A., Min, S., Yoo, K., & Fries, B. C. (2024). Host–Pathogen Interactions and Correlated Factors That Are Affected in Replicative-Aged Cryptococcus neoformans. Journal of Fungi, 10(4), 279. https://doi.org/10.3390/jof10040279