
Biocontrol Potential of Trichoderma Ghanense and Trichoderma Citrinoviride toward Pythium aphanidermatum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials Utilized
2.2. Collection Site and Isolation of the Fungal Pathogen
2.3. Plant Sampling and Isolation of Rhizospheric and Endophytic Fungi
2.4. Characterization of Fungal Isolates
DNA Extraction, Amplification, and Sequencing
2.5. Efficacy of Rhizospheric and Endophytic Fungi as Biocontrol Agents against P. aphanidermatum
2.5.1. In Vitro Evaluation of the Fungal Isolates’ Antifungal Activity towards Fungal Pathogens
2.5.2. Effect of Trichoderma Isolates on Extracellular Conductivity of P. aphanidermatum
2.6. Biocontrol Potential of Trichoderma Isolates against P. aphanidermatum on Cucumber in Pots
Fungal Inoculum Preparation, Experimental Design, and Plant Growth Condition
2.7. Statistical Analysis
3. Results
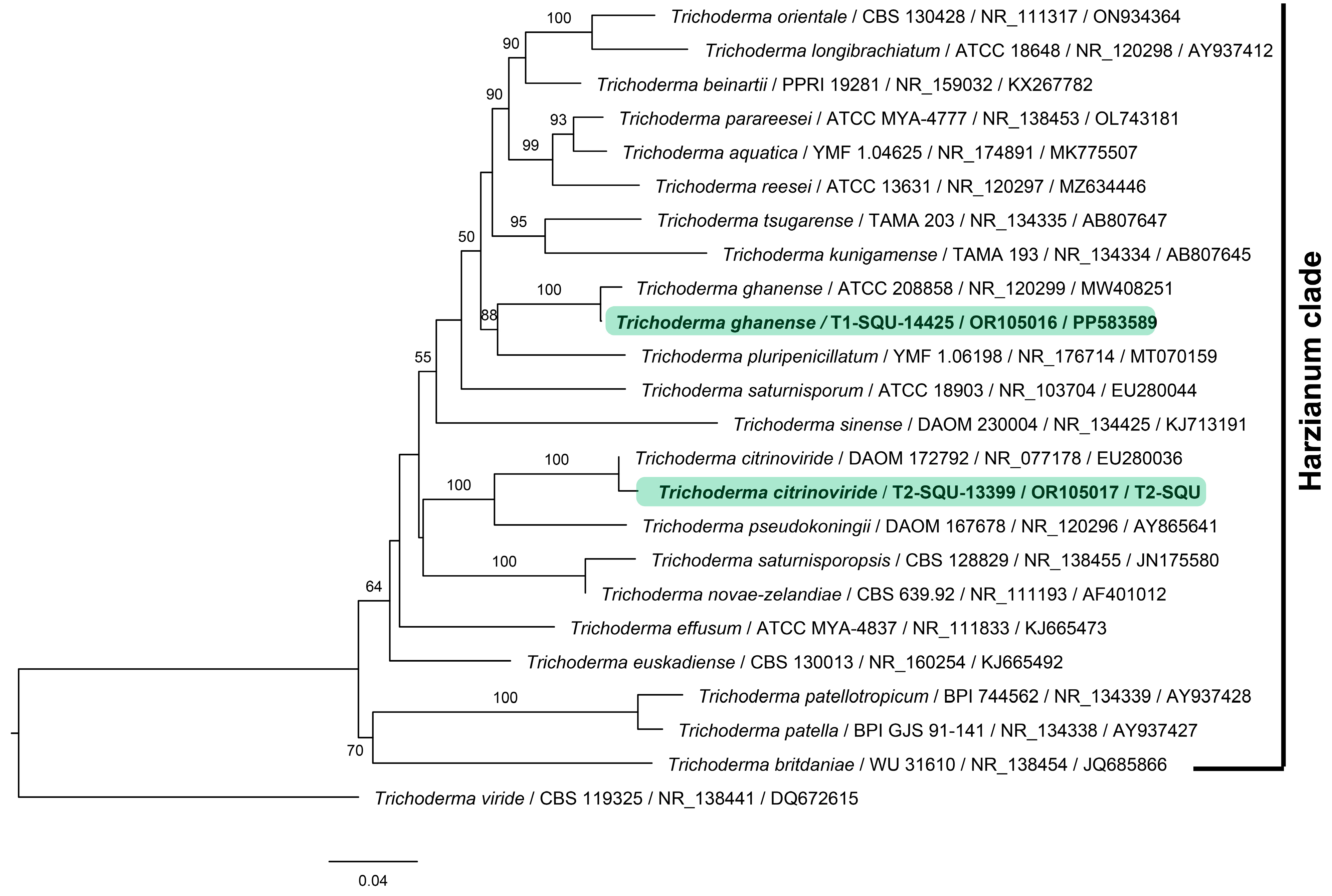
3.1. Identification of the Antagonistic Fungi
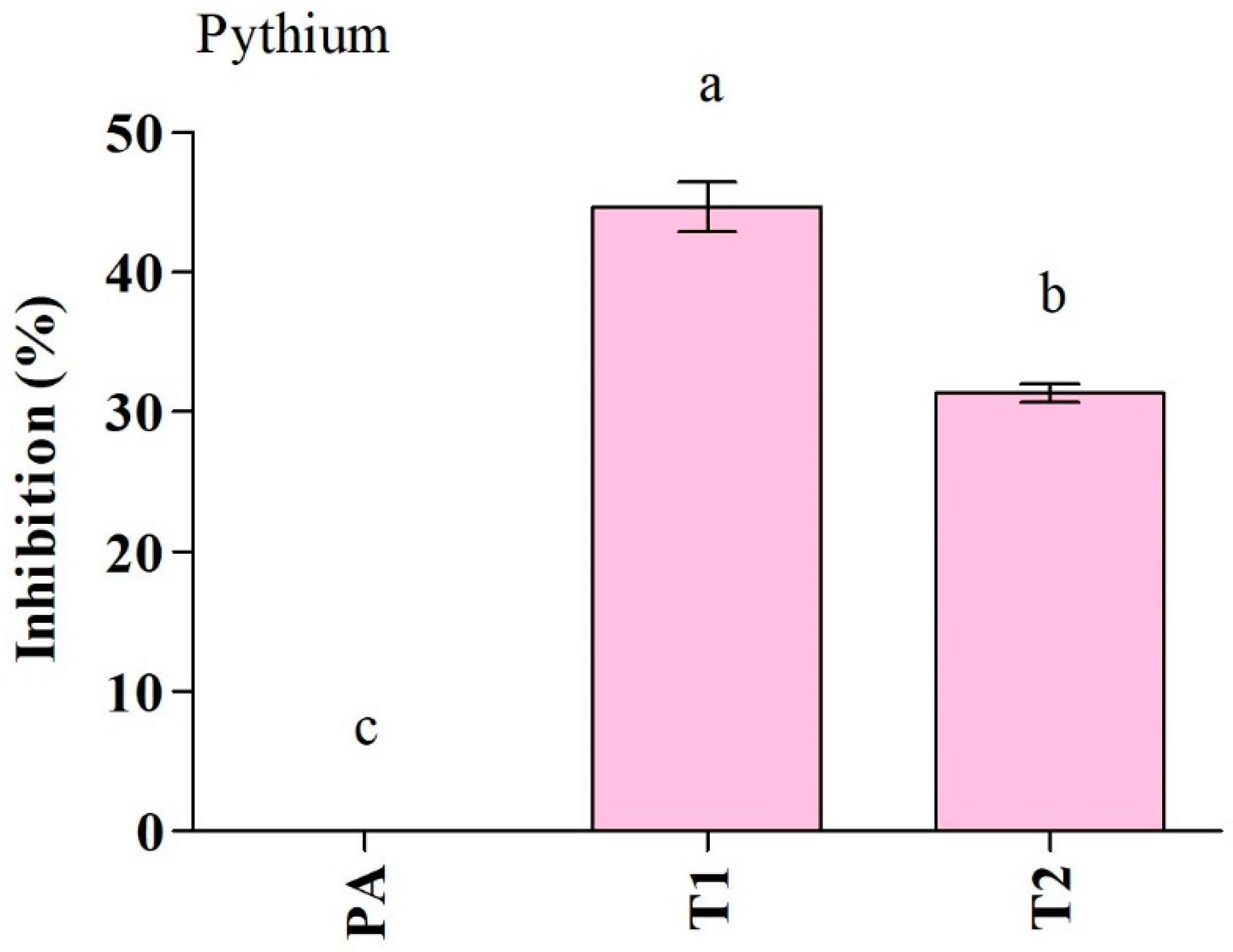
3.2. Antifungal Activity of the Trichoderma Isolates against Fungal Pathogen (P. aphanidermatum)
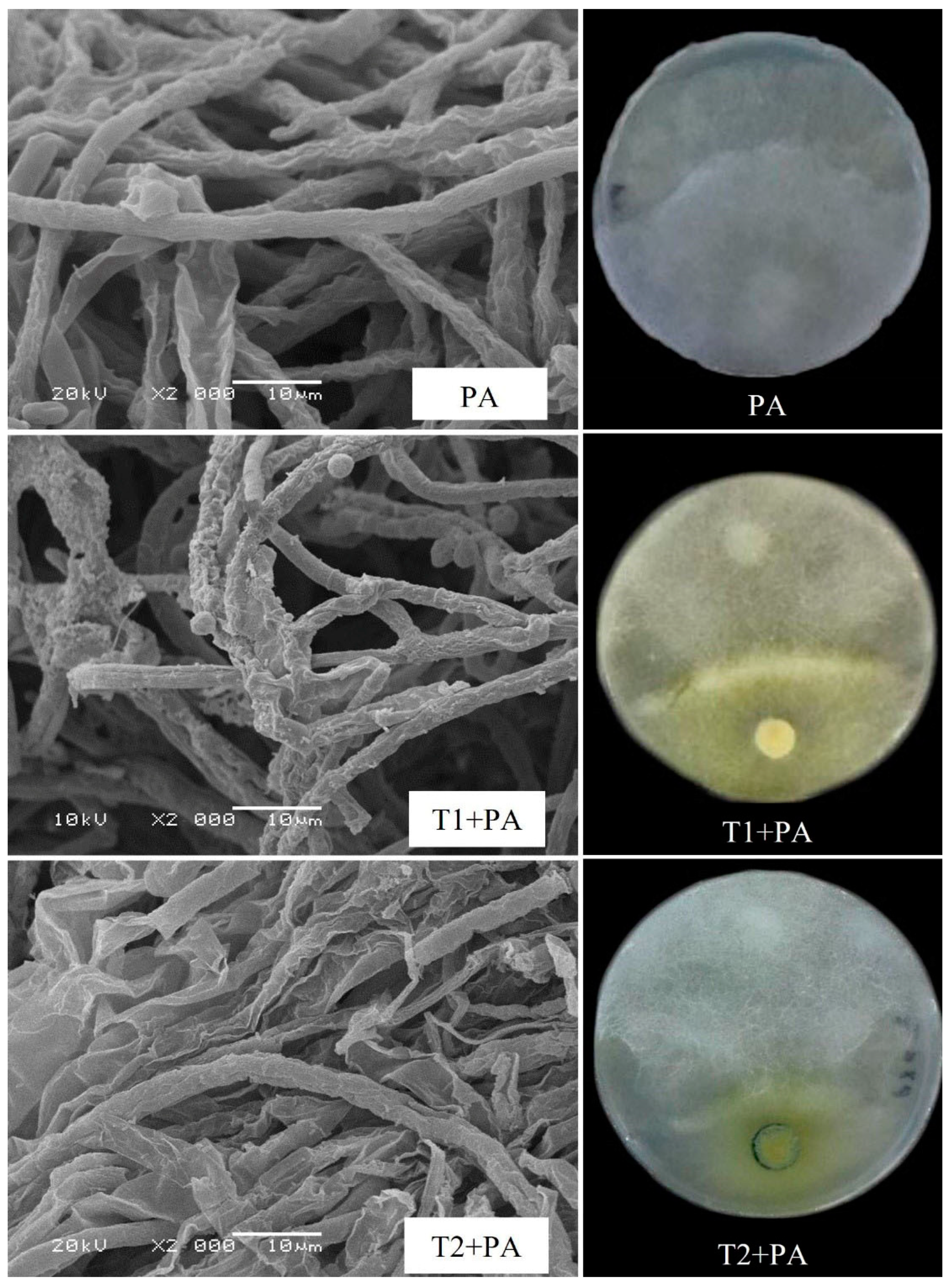
3.2.1. Dual Culture and Scanning Electron Microscope Assays
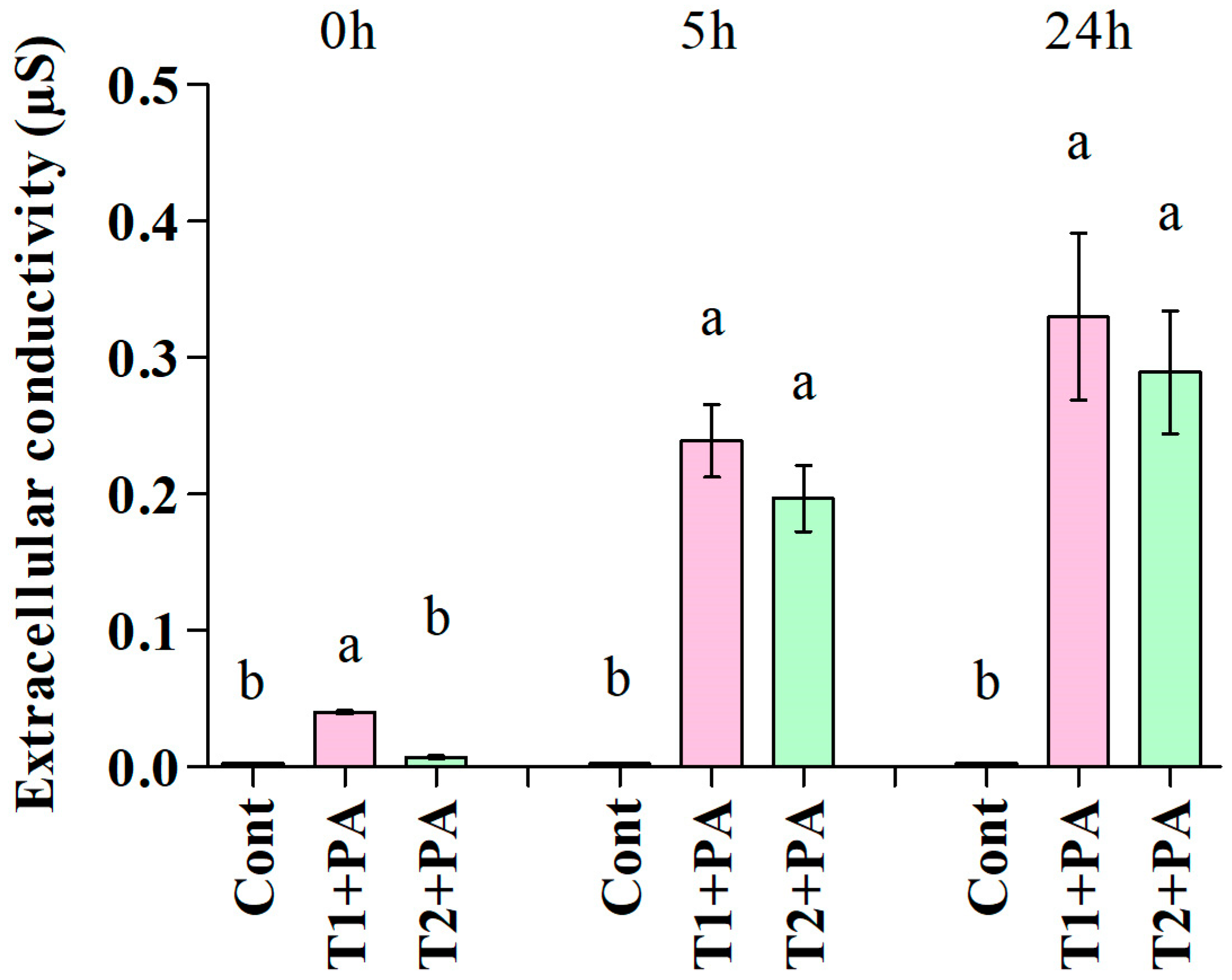
3.2.2. Extracellular Conductivity
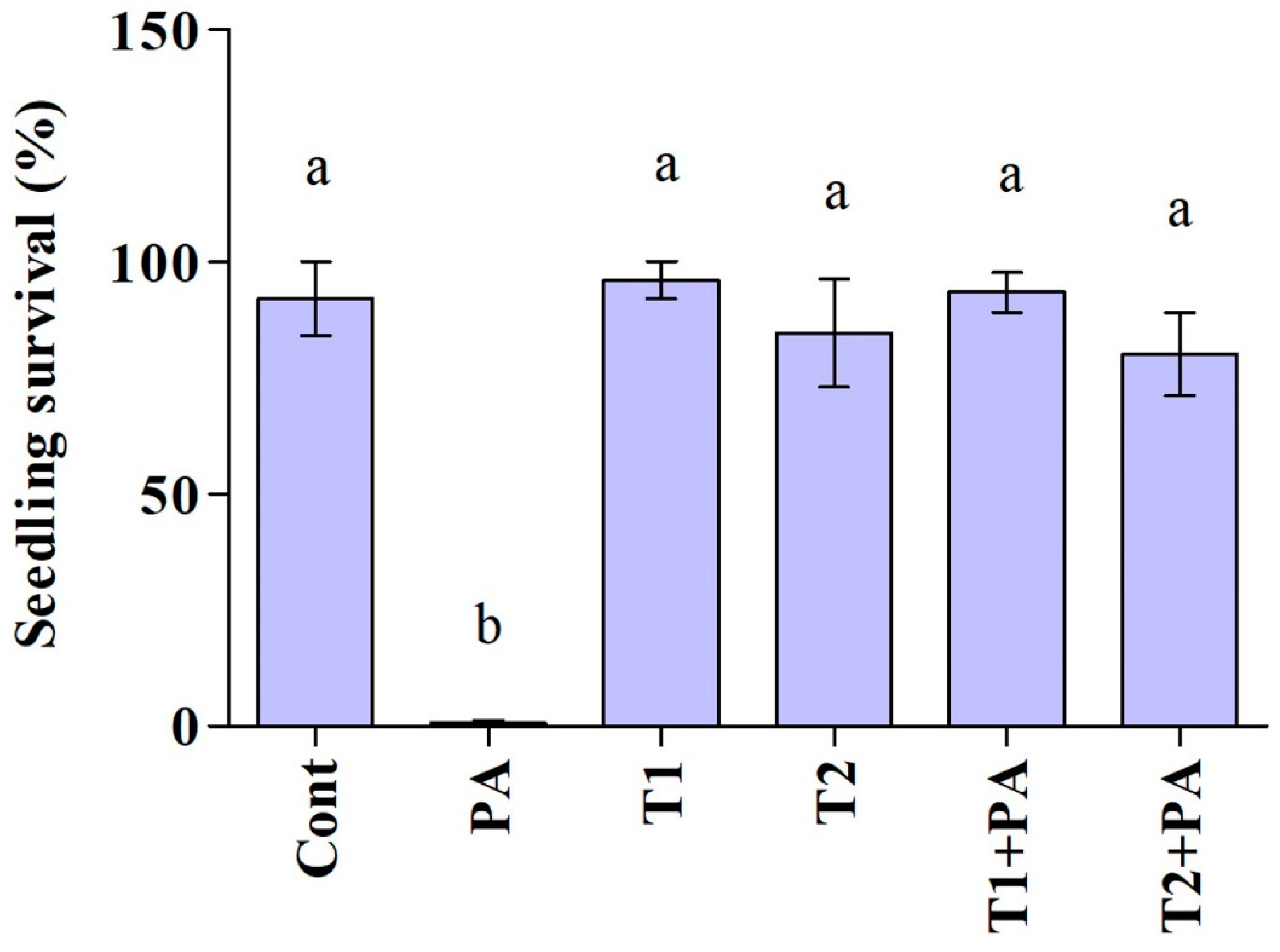
3.3. Effect of Trichoderma Isolates on Cucumber Growth and Damping-Off Disease
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jia, H.; Wang, H. Introductory Chapter: Studies on Cucumber. In Cucumber Economic Values and Its Cultivation and Breeding; Wang, H., Ed.; IntechOpen: Rijeka, Croatia, 2021; Chapter 1. [Google Scholar]
- Qian, C.; Ren, N.; Wang, J.; Xu, Q.; Chen, X.; Qi, X. Effects of exogenous application of CPPU, NAA and GA4+ 7 on parthenocarpy and fruit quality in cucumber (Cucumis sativus L.). Food Chem. 2018, 243, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; Shi, Y.; Han, L.; Li, P.; Zha, Z.; Liu, C.; Liu, X.; Huang, P.; Liu, Y.; Tang, Q. Identification of seven undescribed cucurbitacins in Cucumis sativus (cucumber) and their cytotoxic activity. Phytochemistry 2022, 197, 113123. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Zhang, L.; Zhou, Y.; Fang, Q. Cucurbitacin B alleviates cerebral ischemia/reperfusion injury by inhibiting NLRP3 inflammasome-mediated inflammation and reducing oxidative stress. Biosci. Biotechnol. Biochem. 2022, 86, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Involvement of auxin in growth and stress response of cucumber. Veg. Res. 2022, 2, 13. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Nema, N.K.; Maity, N.; Sarkar, B.K. Phytochemical and therapeutic potential of cucumber. Fitoterapia 2013, 84, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Mandey, J.; Wolayan, F.; Pontoh, C.; Kowel, Y. Effect of orally administrated of cucumber (Cucumis sativus L.) seed juice on the performance and carcass parameters of broiler chickens. IOP Conf. Ser. Earth Environ. Sci. 2020, 492, 012025. [Google Scholar] [CrossRef]
- Ni, L.; Punja, Z.K. Management of fungal diseases on cucumber (Cucumis sativus L.) and tomato (Solanum lycopersicum L.) crops in greenhouses using Bacillus subtilis. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Islam, M.T., Rahman, M.M., Pandey, P., Boehme, M.H., Haesaert, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–28. [Google Scholar]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Redda, E.T.; Ma, J.; Mei, J.; Li, M.; Wu, B.; Jiang, X. Antagonistic potential of different isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. Eur. J. Exp. Biol. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Peng, Y.; Li, S.J.; Yan, J.; Tang, Y.; Cheng, J.P.; Gao, A.J.; Yao, X.; Ruan, J.J.; Xu, B.L. Research Progress on Phytopathogenic Fungi and Their Role as Biocontrol Agents. Front. Microbiol. 2021, 12, 670135. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L.; Toxicology, C. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Kuang, Y.; Splivallo, R.; Chatterjee, P.; Karlovsky, P. Interactions among filamentous fungi Aspergillus niger, Fusarium verticillioides and Clonostachys rosea: Fungal biomass, diversity of secreted metabolites and fumonisin production. BMC Microbiol. 2016, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Khan, S.U.; Khan, W.U.; Saleh, T.A.; Khan, M.H.U.; Ullah, S.; Ali, A.; Ikram, M. Antagonist effects of strains of Bacillus spp. against Rhizoctonia solani for their protection against several plant diseases: Alternatives to chemical pesticides. C. R. Biol. 2019, 342, 124–135. [Google Scholar] [CrossRef]
- Ma, M.; Taylor, P.W.; Chen, D.; Vaghefi, N.; He, J.-Z. Major Soilborne Pathogens of Field Processing Tomatoes and Management Strategies. Microorganism 2023, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Babadoost, M. Oomycete Diseases of Cucurbits: History, Significance, and Management. In Horticultural Reviews; Jules, J., Ed.; 2016; Volume 44, pp. 279–314. [Google Scholar]
- Rosslenbroich, H.-J.; Stuebler, D. Botrytis cinerea—History of chemical control and novel fungicides for its management. Crop Prot. 2000, 19, 557–561. [Google Scholar] [CrossRef]
- Leroux, P. Chemical control of Botrytis cinerea and its resistance to chemical fungicides. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Meszka, B.; Broniarek-Niemiec, A.; Bielenin, A. The status of dodine resistance of Venturia inaequalis populations in Poland. Phytopathol. Pol. 2008, 47, 57–61. [Google Scholar]
- Matson, M.E.; Small, I.M.; Fry, W.E.; Judelson, H.S. Metalaxyl resistance in Phytophthora infestans: Assessing role of RPA190 gene and diversity within clonal lineages. Phytopathology 2015, 105, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A.J.P. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Muthukumar, A.; Eswaran, A.; Sanjeevkumas, K. Exploitation of Trichoderma species on the growth of Pythium aphanidermatum in chilli. Braz. J. Microbiol. 2011, 42, 1598–1607. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases—What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial interactions within multiple-strain biological control agents impact soil-borne plant disease. Front. Microbiol. 2020, 11, 585404. [Google Scholar] [CrossRef]
- Subedi, P.; Gattoni, K.; Liu, W.; Lawrence, K.S.; Park, S.-W. Current utility of plant growth-promoting rhizobacteria as biological control agents towards plant-parasitic nematodes. Plants 2020, 9, 1167. [Google Scholar] [CrossRef]
- Savita, A.; Giri, B.; Prasad, R.; Wu, Q.; Varma, A. Fungi as biological control agents. In Biofertilizers for Sustainable Agriculture and Environment; Springer: Cham, Switzerland, 2019; Volume 55. [Google Scholar]
- Singh, A.; Srivastava, M.; Kumar, V.; Sharma, A.; Pandey, S.; Shahid, M. Exploration and interaction of Trichoderma species and their metabolites by confrontation assay against Pythium aphanidermatum. Int. J. Sci. Res. 2014, 3, 44–48. [Google Scholar]
- El_Komy, M.H.; Saleh, A.A.; Eranthodi, A.; Molan, Y.Y. Characterization of novel Trichoderma asperellum isolates to select effective biocontrol agents against tomato Fusarium wilt. Plant Pathol. J. 2015, 31, 50. [Google Scholar] [CrossRef]
- Galarza, L.; Akagi, Y.; Takao, K.; Kim, C.S.; Maekawa, N.; Itai, A.; Peralta, E.; Santos, E.; Kodama, M. Characterization of Trichoderma species isolated in Ecuador and their antagonistic activities against phytopathogenic fungi from Ecuador and Japan. J. Gen. Plant Pathol. 2015, 81, 201–210. [Google Scholar] [CrossRef]
- Woo, S.L.; Lorito, M. Exploiting the interactions between fungal antagonists, pathogens and the plant for biocontrol. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Springer: Berlin/Heidelberg, Germany, 2007; pp. 107–130. [Google Scholar]
- Hoyos-Carvajal, L.; Duque, G.; Orduz, S. Antagonismo in vitro de Trichoderma spp. sobre aislamientos de Sclerotinia spp. y Rhizoctonia spp. Rev. Colomb. Cienc. Hortícolas 2008, 2, 76–86. [Google Scholar] [CrossRef]
- Hjeljord, L.G.; Tronsmo, A. Effect of germination initiation on competitive capacity of Trichoderma atroviride P1 conidia. J. Phytopathol. 2003, 93, 1593–1598. [Google Scholar] [CrossRef] [PubMed]
- Le, H.; Black, L.; Sikora, R. Evaluation of Trichoderma spp. for biocontrol of tomato sudden caused by Pythium aphanidermatum following flooding in tropical hot season. Commun. Agric. Appl. Biol. Sci. 2003, 68, 463–474. [Google Scholar] [PubMed]
- Watanabe, S.; Kumakura, K.; Kato, H.; Iyozumi, H.; Togawa, M.; Nagayama, K. Identification of Trichoderma SKT-1, a biological control agent against seedborne pathogens of rice. J. Gen. Plant Pathol. 2005, 71, 351–356. [Google Scholar] [CrossRef]
- Karthikeyan, M.; Radhika, K.; Mathiyazhagan, S.; Bhaskaran, R.; Samiyappan, R.; Velazhahan, R. Induction of phenolics and defense-related enzymes in coconut (Cocos nucifera L.) roots treated with biocontrol agents. Braz. J. Plant Physiol. 2006, 18, 367–377. [Google Scholar] [CrossRef]
- Sharma, K.; Mishra, A.K.; Misra, R.S. Morphological, biochemical and molecular characterization of Trichoderma harzianum isolates for their efficacy as biocontrol agents. J. Phytopathol. 2009, 157, 51–56. [Google Scholar] [CrossRef]
- Gveroska, B.; Ziberoski, J. Trichoderma harzianum as a biocontrol agent against Alternaria alternata on tobacco. ATI-Appl. Technol. Innov. 2012, 7, 67–76. [Google Scholar] [CrossRef]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a Biocontrol Agent of Fungal Plant Pathogens. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 497–506. [Google Scholar]
- Swehla, A.; Pandey, A.K.; Nair, R.M. Bioactivity of Trichoderma harzianum isolates against the fungal root rot pathogens with special reference to Macrophomina phaseolina causing dry root rot of mungbean. Indian Phytopathol. 2020, 73, 787–792. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, W.-Y. Trichoderma brevicrassum strain TC967 with capacities of diminishing cucumber disease caused by Rhizoctonia solani and promoting plant growth. Biol. Control 2020, 142, 104151. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Santos, M.; Moreno-Gavíra, A.; Marín-Rodulfo, T.; Gea, F.J.; Diánez, F. Biological control of fungal diseases by Trichoderma aggressivum f. europaeum and its compatibility with fungicides. J. Fungi 2021, 7, 598. [Google Scholar] [CrossRef]
- Ferjani, H.; Bencheikh, Z. Diversity analysis of spontaneous populations of Moraea sisyrunchium (Iridacea) in different bioclimates of Tunisia. J. Anim. Plant Sci. 2022, 52, 9495–9502. [Google Scholar]
- Quattrocchi, U. CRC World Dictionary of Grasses: Common Names, Scientific Names, Eponyms, Synonyms, and Etymology-3 Volume set; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Al-Daghari, D.S.S.; Al-Sadi, A.M.; Al-Mahmooli, I.H.; Janke, R.; Velazhahan, R. Biological Control Efficacy of Indigenous Antagonistic Bacteria Isolated from the Rhizosphere of Cabbage Grown in Biofumigated Soil against Pythium aphanidermatum Damping-Off of Cucumber. Agriculture 2023, 13, 626. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and characterization of fungal endophytes isolated from medicinal plant Ephedra pachyclada as plant growth-promoting. J. Biomol. 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Sarkar, A.; Dutta, S. Characterisation of plant growth promoting traits and stress tolerant properties of Aspergillus terreus (MCC 1819) isolated from the rhizosphere of Aloe barbadensis Mill. and its application. J. Pharmacogn. Phytochem. 2019, 8, 2303–2310. [Google Scholar]
- Cai, F.; Dou, K.; Wang, P.; Chenthamara, K.; Chen, J.; Druzhinina, I.S. The Current State of Trichoderma Taxonomy and Species Identification. In Advances in Trichoderma Biology for Agricultural Applications; Amaresan, N., Sankaranarayanan, A., Dwivedi, M.K., Druzhinina, I.S., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 3–35. [Google Scholar]
- Mazrou, Y.S.; Makhlouf, A.H.; Elseehy, M.M.; Awad, M.F.; Hassan, M.M. Antagonistic activity and molecular characterization of biological control agent Trichoderma harzianum from Saudi Arabia. Egypt. J. Biol. Pest Control 2020, 30, 4. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Masoodi, R.S.; Al-Ismaili, M.; Al-Mahmooli, I.H. Population structure and development of resistance to hymexazol among Fusarium solani populations from date palm, citrus and cucumber. J. Phytopathol. 2015, 163, 947–955. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38—Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lakhani, H.N.; Vakharia, D.N.; Hassan, M.M.; Eissa, R.A. Fingerprinting and molecular comparison among two parental strains of Trichoderma spp. and their corresponding fusants produced by protoplast fusion. Biotechnol. Biotechnol. Equip. 2016, 30, 1065–1074. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary 623 Biology, University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 January 2022).
- Ji, S.H.; Gururani, M.A.; Chun, S.-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Kunova, A.; Bonaldi, M.; Saracchi, M.; Pizzatti, C.; Chen, X.; Cortesi, P. Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 2016, 16, 272. [Google Scholar] [CrossRef]
- Heckman, C.; Kanagasundaram, S.; Cayer, M.; Paige, J. Preparation of Cultured Cells for Scanning Electron Microscope. Protoc. Exch. 2007. [Google Scholar] [CrossRef]
- Manhas, R.K.; Kaur, T. Biocontrol potential of Streptomyces hydrogenans strain DH16 toward Alternaria brassicicola to control damping off and black leaf spot of Raphanus sativus. Front. Plant Sci. 2016, 7, 1869. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Abo-Elyousr, K.A.M.; Mousa, M.A.A.; Saad, M.M. Use of Trichoderma culture filtrates as a sustainable approach to mitigate early blight disease of tomato and their influence on plant biomarkers and antioxidants production. Front. Plant Sci. 2023, 14, 1192818. [Google Scholar] [CrossRef] [PubMed]
- Benito, E.P.; Ten Have, A.; van’t Klooster, J.W.; van Kan, J.A. Fungal and plant gene expression during synchronized infection of tomato leaves by Botrytis cinerea. Eur. J. Plant Pathol. 1998, 104, 207–220. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-P.; Pan, Z.-C.; Yang, L.-N.; Burdon, J.J.; Friberg, H.; Sui, Q.-j.; Zhan, J. Optimizing plant disease management in agricultural ecosystems through rational in-crop diversification. Front. Plant Sci. 2021, 12, 767209. [Google Scholar] [CrossRef]
- Pathak, V.M.; Verma, V.K.; Rawat, B.S.; Kaur, B.; Babu, N.; Sharma, A.; Dewali, S.; Yadav, M.; Kumari, R.; Singh, S. Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: A comprehensive review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef] [PubMed]
- He, D.-C.; He, M.-H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Mahmoud, A.F.; Abdalla, O.A.; Protection, P. Biological control of fungi associated with damping-off and root rot disease of cucumber (Cucumis sativus L.). Arch. Phytopathol. 2021, 54, 870–885. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 2023, 14, 1160551. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Ahmad, J.S.; Baker, R. Integration of Pythium nunn and Trichoderma harzianum isolate T-95 for the biological control of Pythium damping-off of cucumber. Plant Soil 1990, 121, 243–250. [Google Scholar] [CrossRef]
- Huang, X.; Cui, H.; Yang, L.; Lan, T.; Zhang, J.; Cai, Z. The microbial changes during the biological control of cucumber damping-off disease using biocontrol agents and reductive soil disinfestation. Biocontrol 2017, 62, 97–109. [Google Scholar] [CrossRef]
- Park, Y.-H.; Mishra, R.C.; Yoon, S.; Kim, H.; Park, C.; Seo, S.-T.; Bae, H. Endophytic Trichoderma citrinoviride isolated from mountain-cultivated ginseng (Panax ginseng) has great potential as a biocontrol agent against ginseng pathogens. J. Ginseng Res. 2019, 43, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Khadka, R.B.; Rotondo, F.; Miller, S.A. The pursuit in Nepal of native Trichoderma spp. for plant disease biocontrol. PhytoFrontiers 2022, 2, 242–256. [Google Scholar] [CrossRef]
- Risoli, S.; Cotrozzi, L.; Sarrocco, S.; Nuzzaci, M.; Pellegrini, E.; Vitti, A. Trichoderma-induced resistance to Botrytis cinerea in Solanum species: A Meta-Analysis. Plants 2022, 11, 180. [Google Scholar] [CrossRef]
- Aswani, R.; Thomas, R.; Radhakrishnan, E. Induction of plant defense response by endophytic microorganisms. In Biocontrol Mechanisms of Endophytic Microorganisms; Elsevier: Amsterdam, The Netherlands, 2022; pp. 89–115. [Google Scholar]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Zhang, F.; Ge, H.; Zhang, F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yan, L.; Zhang, M.; Chang, X.; Zhu, D.; Wei, D.; Naeem, M.; Song, C.; Wu, X.; Liu, T. Changes in the density and composition of rhizosphere pathogenic Fusarium and beneficial Trichoderma contributing to reduced root rot of intercropped soybean. Pathogens 2022, 11, 478. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1934578X1200701133. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Treatment |
|---|---|
| Cont | treated with sterile distilled water |
| T1 | treated with Trichoderma ghanense |
| T2 | treated with Trichoderma citrinoviride |
| PA | treated with Pythium aphanidermatum |
| T1 + PA | treated with Trichoderma ghanense + Pythium aphanidermatum |
| T2 + PA | treated with Trichoderma citrinoviride + Pythium aphanidermatum |
| Sample Code | Fungal Species | Isolated Host | Source of Isolation | GenBank Accession No. | Location | Antagonistic Activity | |
|---|---|---|---|---|---|---|---|
| ITS | EF1α | Pythium aphanidermatum | |||||
| T1 | Trichoderma ghanense | Dactyloctenium robecchii | stem | OR105016 | PP583589 | Al-Seeb | + |
| T2 | Trichoderma citrinoviride | Moraea sisyrinchium | soil | OR105017 | T2-SQU * | Al-Seeb | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shuaibi, B.K.; Kazerooni, E.A.; Al-Maqbali, D.; Al-Kharousi, M.; Al-Yahya’ei, M.N.; Hussain, S.; Velazhahan, R.; Al-Sadi, A.M. Biocontrol Potential of Trichoderma Ghanense and Trichoderma Citrinoviride toward Pythium aphanidermatum. J. Fungi 2024, 10, 284. https://doi.org/10.3390/jof10040284
Al-Shuaibi BK, Kazerooni EA, Al-Maqbali D, Al-Kharousi M, Al-Yahya’ei MN, Hussain S, Velazhahan R, Al-Sadi AM. Biocontrol Potential of Trichoderma Ghanense and Trichoderma Citrinoviride toward Pythium aphanidermatum. Journal of Fungi. 2024; 10(4):284. https://doi.org/10.3390/jof10040284
Chicago/Turabian StyleAl-Shuaibi, Badriya Khalfan, Elham Ahmed Kazerooni, Dua’a Al-Maqbali, Moza Al-Kharousi, Mohamed N. Al-Yahya’ei, Shah Hussain, Rethinasamy Velazhahan, and Abdullah Mohammed Al-Sadi. 2024. "Biocontrol Potential of Trichoderma Ghanense and Trichoderma Citrinoviride toward Pythium aphanidermatum" Journal of Fungi 10, no. 4: 284. https://doi.org/10.3390/jof10040284
APA StyleAl-Shuaibi, B. K., Kazerooni, E. A., Al-Maqbali, D., Al-Kharousi, M., Al-Yahya’ei, M. N., Hussain, S., Velazhahan, R., & Al-Sadi, A. M. (2024). Biocontrol Potential of Trichoderma Ghanense and Trichoderma Citrinoviride toward Pythium aphanidermatum. Journal of Fungi, 10(4), 284. https://doi.org/10.3390/jof10040284