
The Combination of Enzymes and Conidia of Entomopathogenic Fungi against Aphis gossypii Nymphs and Spodoptera frugiperda Larvae





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Enzyme Assay
2.3. Thermostability
2.4. Virulence Assay
2.4.1. Aphis gossypii
2.4.2. Spodoptera frugiperda
2.5. Statistical Analysis
3. Results
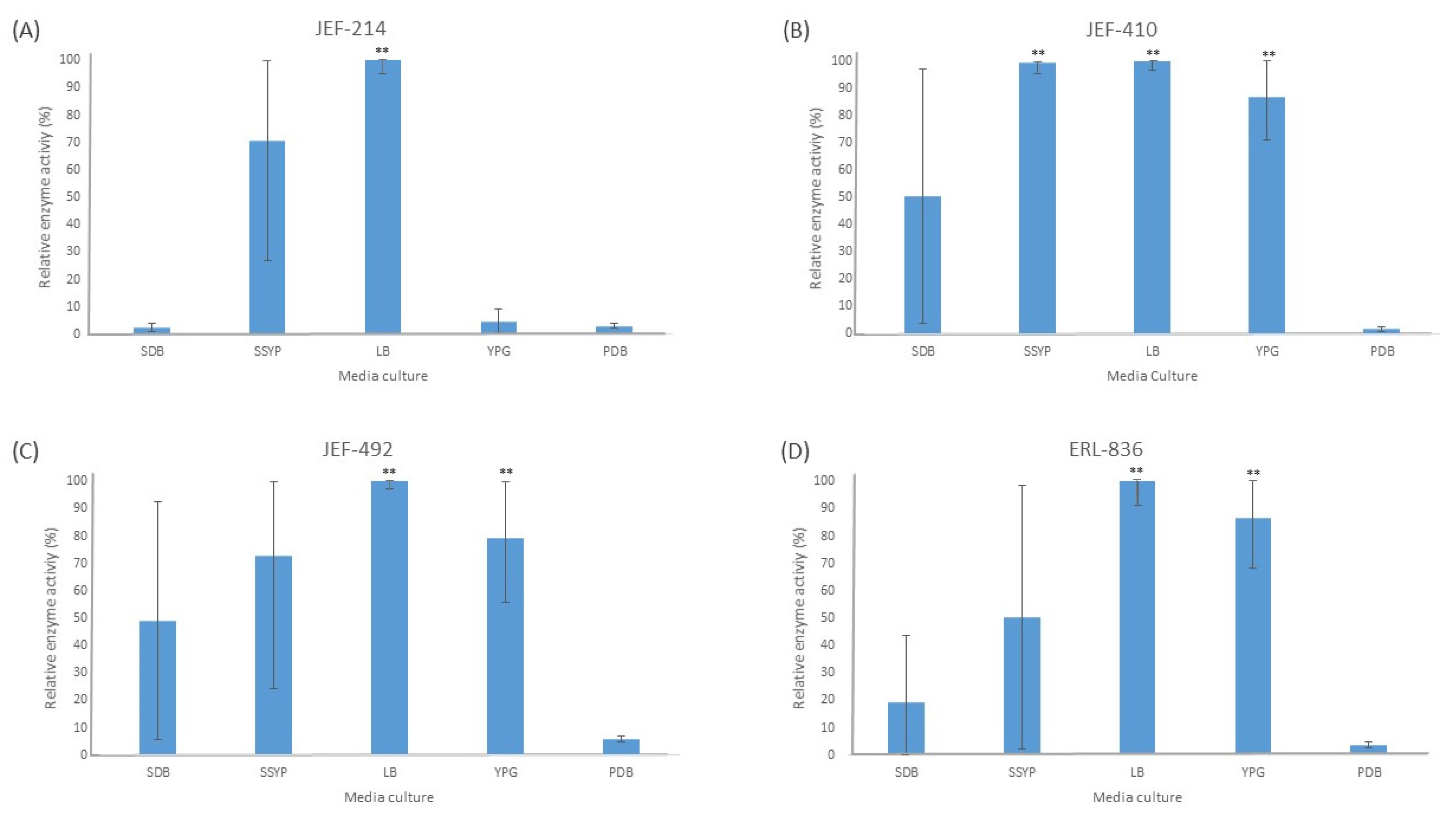
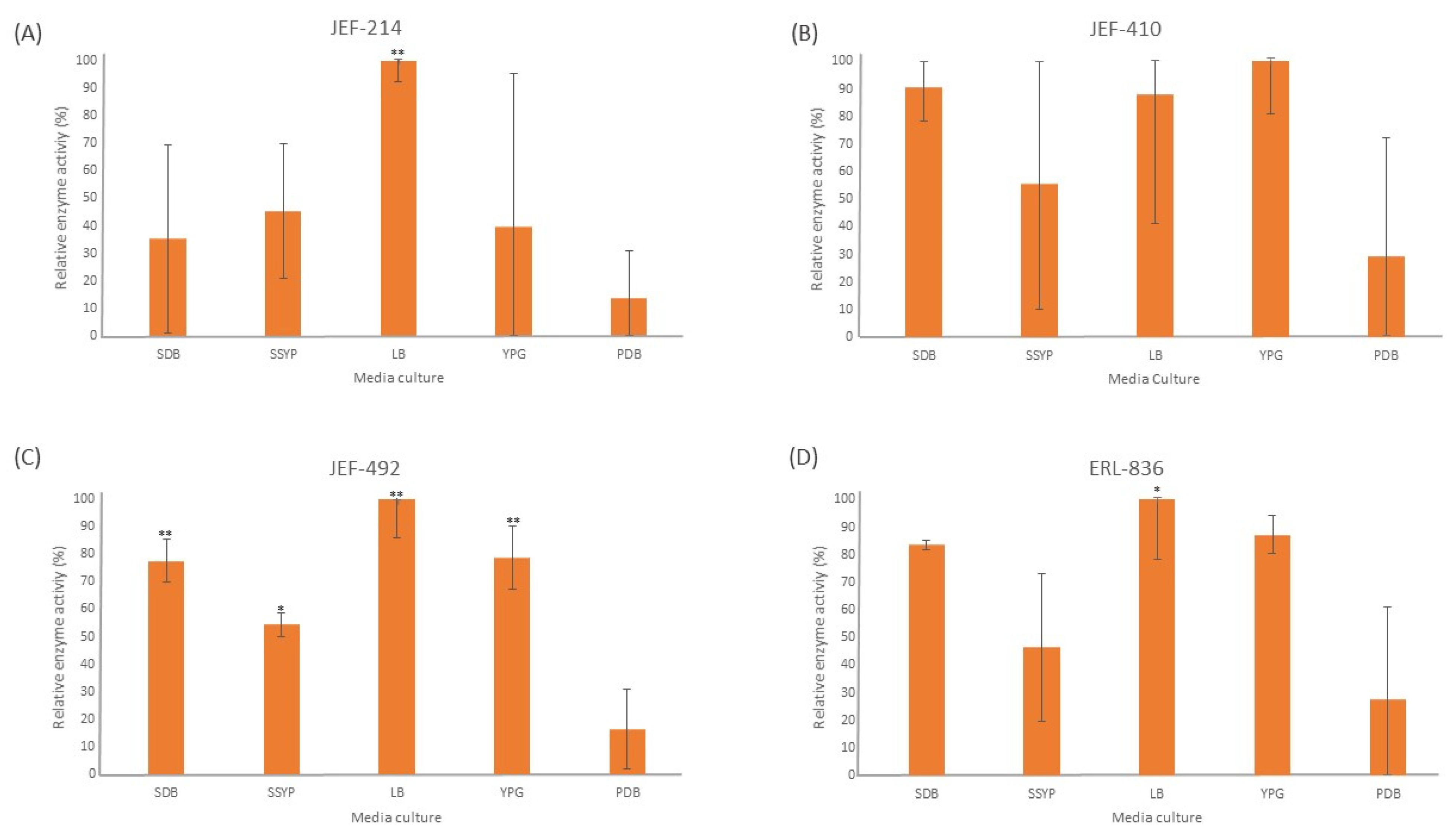
3.1. Enzyme Activity
3.1.1. Protease
3.1.2. Lipase
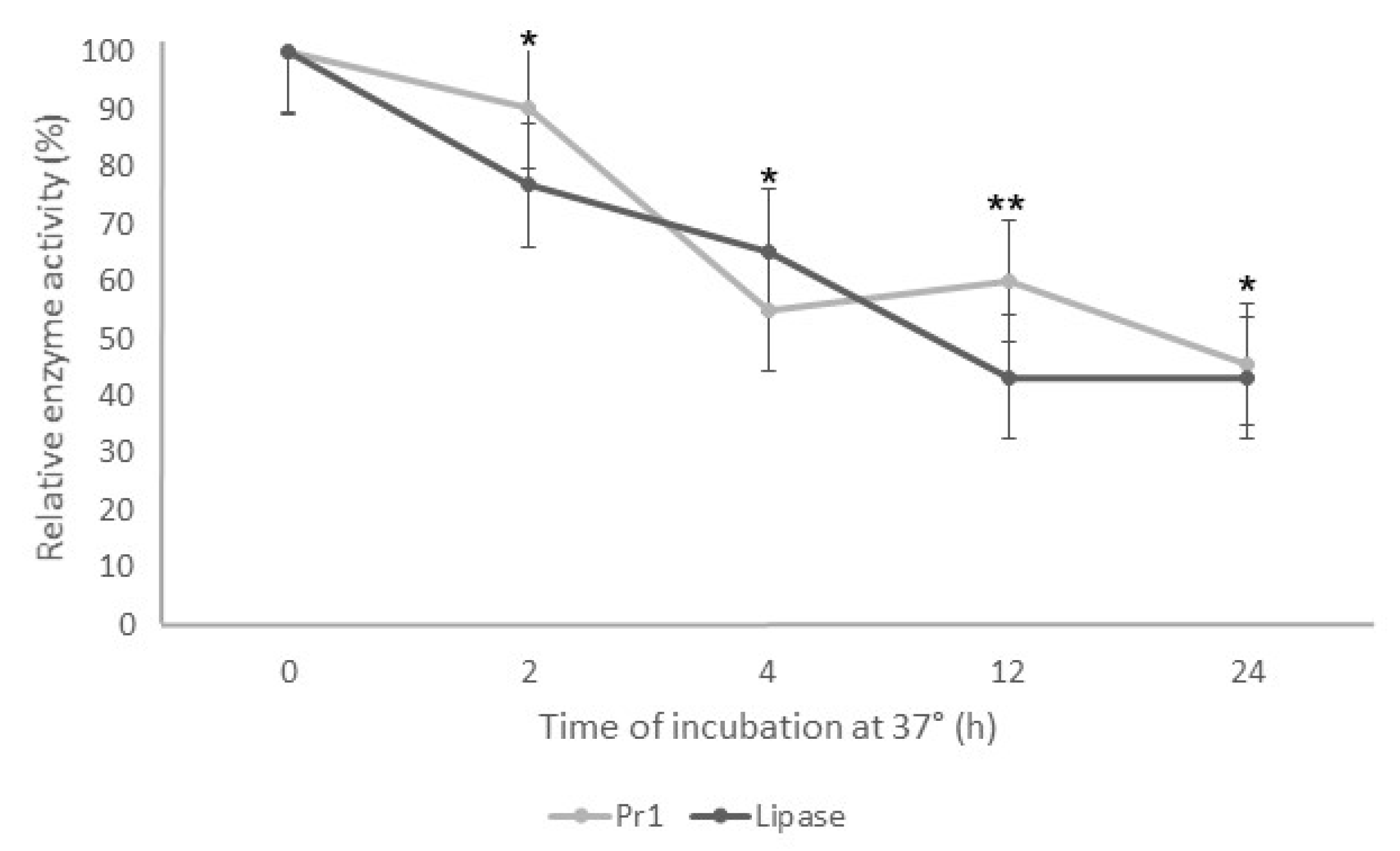
3.2. Thermostability of Supernatant
3.3. Virulence
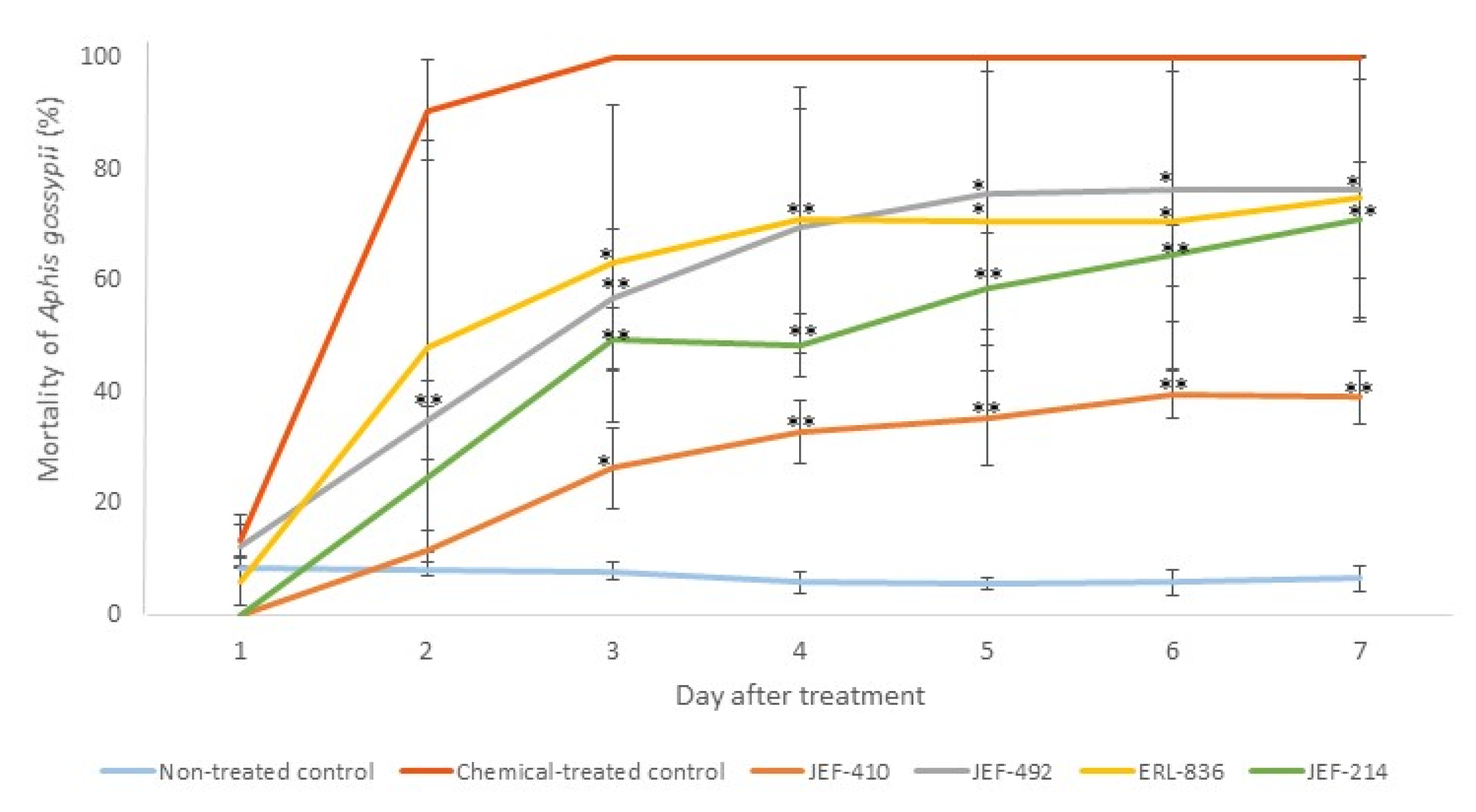
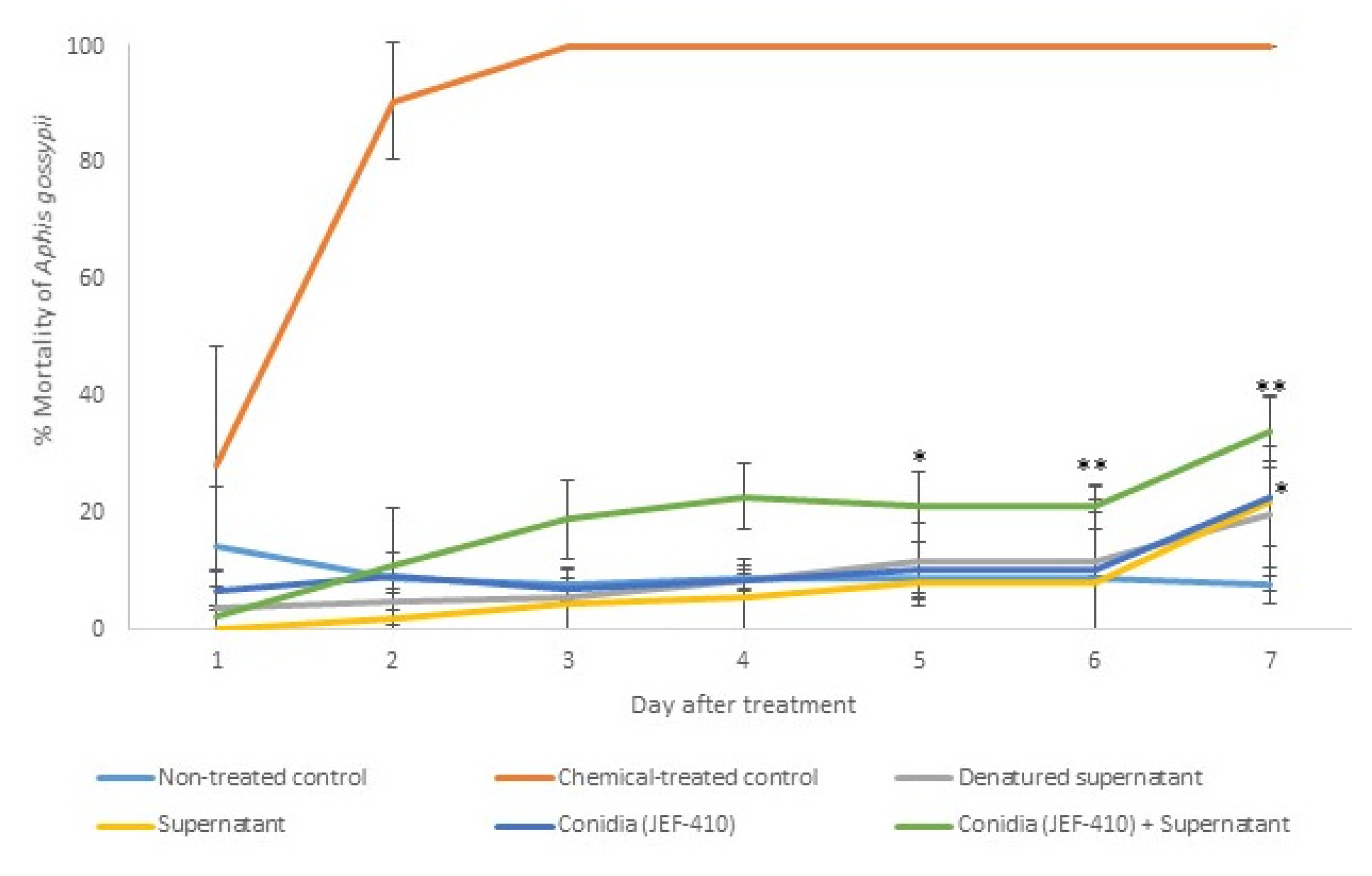
3.3.1. Aphis gossypii
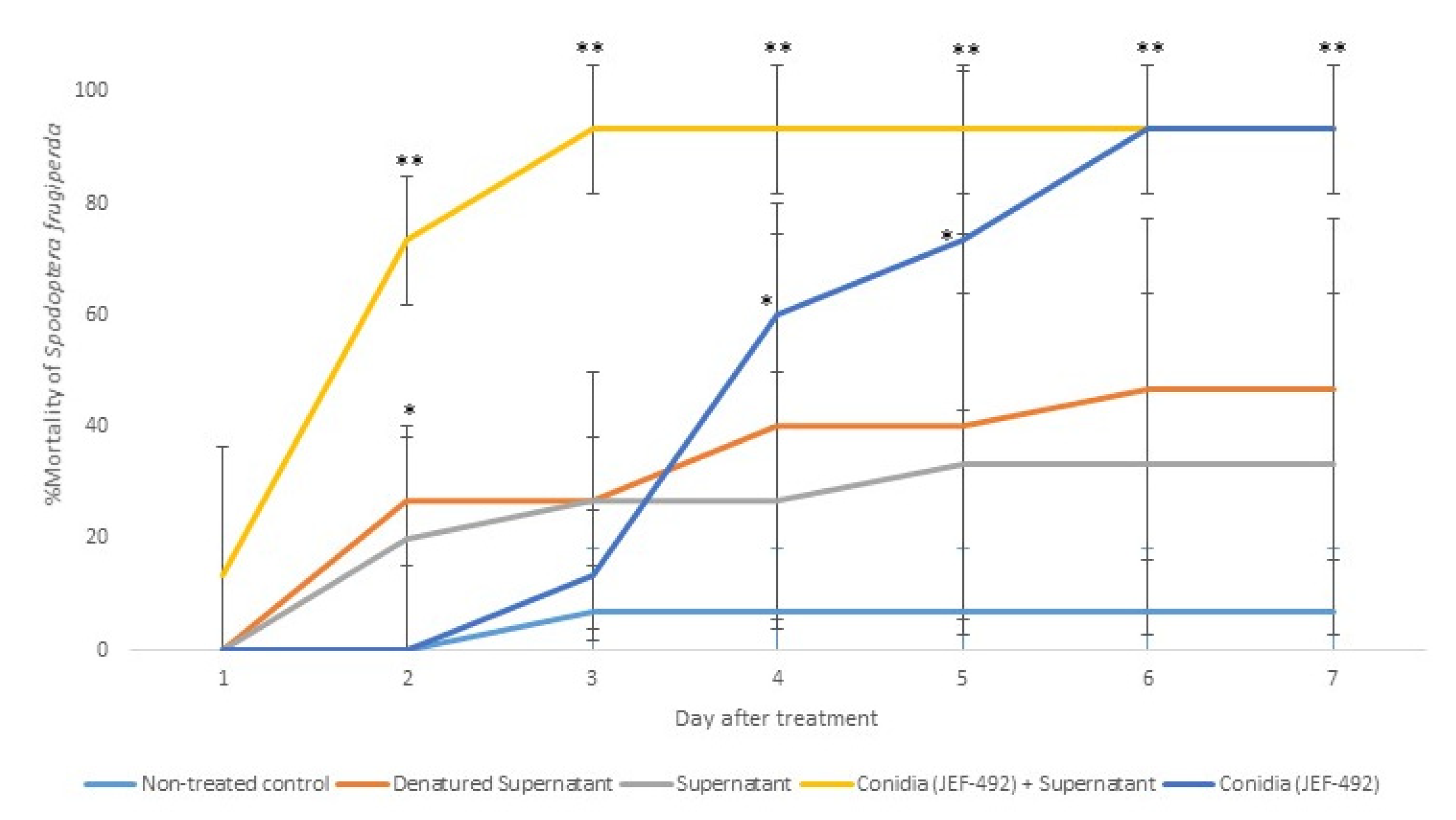
3.3.2. Spodoptera frugiperda
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Mseddi, J.; Ben Farhat-Touzri, D.; Azzouz, H. Selection and characterization of thermotolerant Beauveria bassiana isolates and with insecticidal activity against the cotton-melon aphid Aphis gossypii (Glover) (Hemiptera: Aphididae). Pest Manag. Sci. 2022, 78, 2183–2195. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Sánchez, F.A.; Rivera, G.; Bocanegra-García, V.; Martínez-Padrón, H.Y.; Berrones-Morales, M.; Niño-García, N.; Herrera-Mayorga, V. Advances in Control Strategies against Spodoptera frugiperda. A Review. Molecules 2021, 26, 5587. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Lopes, R.B.; Delalibera Junior, I.; Fernandes, E.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrate and microorganisms: Plenty of new opportunities. BioControl 2017, 63, 39–59. [Google Scholar] [CrossRef]
- Alves, S.B.; Almeida, J.E.M.; Moino, A., Jr.; Alves, L.F.A. Técnicas de laboratório 637-771. In Controle Microbiano de Insetos; Alves, S.B., Ed.; Fealq: Piracicaba, Brazil, 1998; p. 1163. [Google Scholar]
- Ferreira, J.M.; Soares, F.E.F. Entomopathogenic fungi hydrolytic enzymes: A new approach to biocontrol? J. Nat. Pestic. Res. 2023, 3, 100020. [Google Scholar] [CrossRef]
- Pedrini, N. The Entomopathogenic Fungus Beauveria bassiana Shows Its Toxic Side within Insects: Expression of Genes Encoding Secondary Metabolites during Pathogenesis. J. Fungi 2022, 8, 488. [Google Scholar] [CrossRef]
- Mondal, S.; Baksi, S.; Koris, A.; Vatai, G. Journey of enzymes in entomopathogeni fungi. Pac. Sci. 2016, 18, 85–99. [Google Scholar] [CrossRef]
- Schrank, A.; Vainstein, M.H. Metarhizium anisopliae enzymes and toxins. Toxicon 2010, 56, 1267–1274. [Google Scholar] [CrossRef]
- Rosa-Garcia, N.M.; López-Barrera, G.L.; Mireles-Martínez, M.; Villegas-Mendoza, J.M. Metarhizium anisopliae Pr1 Isoform Activation during Pathogenesis in Diaphorina citri. Southwest. Entomol. 2018, 43, 199–207. [Google Scholar] [CrossRef]
- Khun, K.K.; Wilson, B.A.L.; Stevens, M.M.; Huwer, R.K.; Ash, G.J. Integration of entomopathogenic fungi into IPM programs: Studies involving Weevils (Coleoptera: Curculionoidea) affecting horticultural crops. Insects 2020, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.H.A.; Mohammed, S.H.; Eltaly, R.I.; Moustafa, M.A.M.; Fónagy, A.; Farag, S.M. Co-application of entomopathogenic fungi with chemical insecticides against culex pipiens. J. Invertebr. Pathol. 2023, 198, 107916. [Google Scholar] [CrossRef] [PubMed]
- Vidhate, R.; Singh, J.; Ghormade, V.; Chavan, S.B.; Patil, A.; Deshpande, M.V. Use of hydrolytic enzymes of Myrothecium verrucaria and conidia of Metarhizium anisopliae, singly and sequentially to control pest and pathogens in grapes and their compatibility with pesticides used in the field. Biopestic. Int. 2015, 11, 48–60. [Google Scholar]
- Zhang, P.; Zhang, X.; Zhao, Y.; Ren, Y.; Mu, W.; Liu, F. Efficacy of granular applications of clothianidin and nitenpyram against Aphis gossypii (Glover) and Apolygus lucorum (Meyer-Dür) in cotton fields in China. Crop Prot. 2015, 78, 27–34. [Google Scholar] [CrossRef]
- Ma, K.S.; Tang, Q.L.; Liang, P.Z.; Li, J.H.; Gao, X.W. A sublethal concentration of afidopyropen suppresses the population growth of the cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). J. Integr. Agric. 2022, 21, 2055–2064. [Google Scholar] [CrossRef]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Prot. 2021, 145, 105641. [Google Scholar] [CrossRef]
- FAO. The Global Action for Fall Armyworm Control: Action Framework 2020–2022. Working Together to Tame the Global Threat; Food and Agriculture Organization of United Nations: Rome, Italy, 2020.
- Kim, J.C.; Lee, M.R.; Kim, S.; Lee, S.J.; Park, S.E.; Nai, Y.S.; Lee, G.S.; Shin, T.Y.; Kim, J.S. Tenebrio molitor-mediated entomopathogenic fungal library construction for pest management. J. Asia-Pac. Entomol. 2018, 21, 196–204. [Google Scholar] [CrossRef]
- Campos, R.A.; Arruda, W.; Boldo, J.T.; Silva, M.V.; Barros, N.M.; Azevedo, J.L.; Schrank, A.; Vainstein, M.H. Boophilus microplus infection by Beauveria amorpha and Beauveria bassiana: SEM analysis and regulation of subtilisin-like proteases and chitinases. Curr. Microbiol. 2005, 50, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Tsujita, T.; Ninomiya, H.; Okuda, H. P-nitrophenyl butyrate hydrolyzing activity of hormone-sensitive lipase from bovine adipose tissue. J. Lipid. Res. 1989, 30, 997–1004. [Google Scholar] [CrossRef]
- Im, Y.; Park, S.-E.; Lee, S.Y.; Kim, J.-C.; Kim, J.S. Early-Stage Defense Mechanism of Cotton Aphid Aphis gossypii against infection with the insect-killing fungus Beauveria bassiana JEF-544. Front. Immunol. 2022, 13, 907088. [Google Scholar] [CrossRef]
- Jeon, I.; Kim, J.S. Soil treatment with Beauveria and Metarhizium to control fall armyworm, Spodoptera frugiperda, during the soil-dwelling stage. J. Asia-Pac. Entomol. 2024, 27, 102193. [Google Scholar] [CrossRef]
- Jung, Y.; Yang, H.; Lee, I.-Y.; Yong, T.-S.; Lee, S. Core/Sheath-Structured Composite Nanofibers Containing Cinnamon Oil: Their Antibacterial and Antifungal Properties and Acaricidal Effect against House Dust Mites. Polymers 2020, 12, 243. [Google Scholar] [CrossRef] [PubMed]
- Ayres, M.; Ayres Júnior, M.; Ayres, D.L.; Santos, A.A. Bioestat—Aplicações Estatísticas nas Áreas das Ciências Bio-Médicas; Ong Mamirauá: Belém, PA, Brazil, 2007. [Google Scholar]
- Montesinos-Matías, R.; Ordaz-Hernández, A.; Angel-Cuapio, A.; Colin-Bonifacio, Y.; Garcia-Garcia, R.E.; Ángel-Sahagún, C.A.; Arredondo-Bernal, H.C. Principal component analysis of the biological characteristics of entomopathogenic fungi in nutrient-limited and cuticle-based media. J. Basic Microbiol. 2021, 61, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.A.; Wang, C.S.; Butt, T.M. Nutrition influences growth and virulence of the insect-pathogenic fungus Metarhizium anisopliae. FEMS Microbiol. Rev. 2005, 251, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Andreis, F.C.; Schrank, A.; Thompson, C.E. Molecular evolution of Pr1 proteases depicts ongoing diversification in Metarhizium spp. Mol. Genet. Genom. 2019, 294, 901–917. [Google Scholar] [CrossRef] [PubMed]
- Ayudya, D.R.; Herlinda, S.; Suwandi, S. Insecticidal activity of culture filtrates from liquid medium of Beauveria bassiana isolates from South Sumatra (Indonesia) wetland soil against larvae of Spodoptera litura. Biodivers. J. 2019, 20, 8. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Carrasco-Díaz, J.-A.; Santiago-Álvarez, C. Insecticidal and antifeedant activities of proteins secreted by entomopathogenic fungi against Spodoptera littoralis (Lep., Noctuidae). J. Appl. Entomol. 2006, 130, 442–452. [Google Scholar] [CrossRef]
- Nirmal, N.P.; Laxman, R.S. Enhanced thermostability of a fungal alkaline protease by different additives. Enzyme Res. 2014, 2014, 109303. [Google Scholar] [CrossRef]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Rougier, M. Effect of temperature on vegetative growth of Beauveria bassiana isolates from different origins. Mycologia 1997, 89, 389–392. [Google Scholar] [CrossRef]
- Fargues, J.; Maniania, N.K.; Delmas, J.C.; Smits, N. Influence de la température sur la croissance in vitro d’hyphomycètes entomopathogènes. Agronomie 1992, 12, 557–564. [Google Scholar] [CrossRef]
- Bhosale, S.H.; Rao, M.B.; Deshpande, V.V.; Srinivasan, M.C. Thermostability of high-activity alkaline protease from Conidiobolus coronatus (NCL 86.8.20). Enzyme Microb. Technol. 1995, 17, 136–139. [Google Scholar] [CrossRef]
- Lin, K.P.; Feng, G.J.; Pu, F.L.; Hou, X.D.; Cao, S.L. Enhancing the Thermostability of Papain by Immobilizing on Deep Eutectic Solvents-Treated Chitosan with Optimal Microporous Structure and Catalytic Microenvironment. Front. Bioeng. Biotechnol. 2020, 8, 576266. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, J.M.; Fernandes, É.K.K.; Kim, J.S.; Soares, F.E.F. The Combination of Enzymes and Conidia of Entomopathogenic Fungi against Aphis gossypii Nymphs and Spodoptera frugiperda Larvae. J. Fungi 2024, 10, 292. https://doi.org/10.3390/jof10040292
Ferreira JM, Fernandes ÉKK, Kim JS, Soares FEF. The Combination of Enzymes and Conidia of Entomopathogenic Fungi against Aphis gossypii Nymphs and Spodoptera frugiperda Larvae. Journal of Fungi. 2024; 10(4):292. https://doi.org/10.3390/jof10040292
Chicago/Turabian StyleFerreira, Juliana M., Éverton K. K. Fernandes, Jae Su Kim, and Filippe Elias F. Soares. 2024. "The Combination of Enzymes and Conidia of Entomopathogenic Fungi against Aphis gossypii Nymphs and Spodoptera frugiperda Larvae" Journal of Fungi 10, no. 4: 292. https://doi.org/10.3390/jof10040292
APA StyleFerreira, J. M., Fernandes, É. K. K., Kim, J. S., & Soares, F. E. F. (2024). The Combination of Enzymes and Conidia of Entomopathogenic Fungi against Aphis gossypii Nymphs and Spodoptera frugiperda Larvae. Journal of Fungi, 10(4), 292. https://doi.org/10.3390/jof10040292