
Transcriptome Analysis of Picea crassifolia in Response to Rust Infestation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Preparation and Microscopic Observation
2.3. RNA Isolation
2.4. Data Processing
2.5. RT-qPCR Validation of Differential Gene Expression
3. Results and Analysis
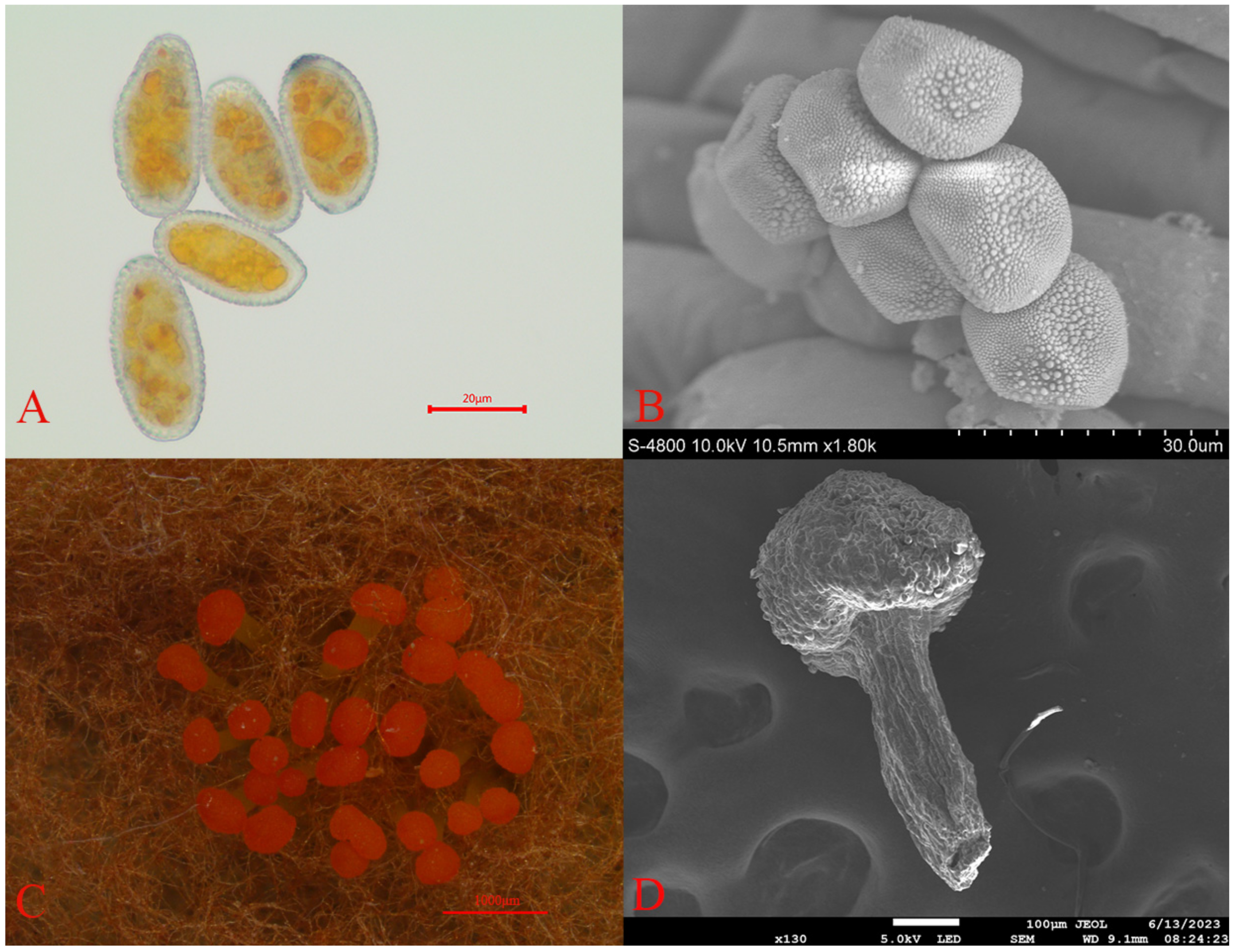
3.1. Pathogen Identification
3.2. Differences in Stomatal Morphology of Spruce Needles
3.3. Morphological Differences in the Wax Layer of One-Year-Old and Two-Year-Old Spruce Needles
3.4. Sequencing Data and Quality Assessment
3.5. Screening of Differentially Expressed Genes
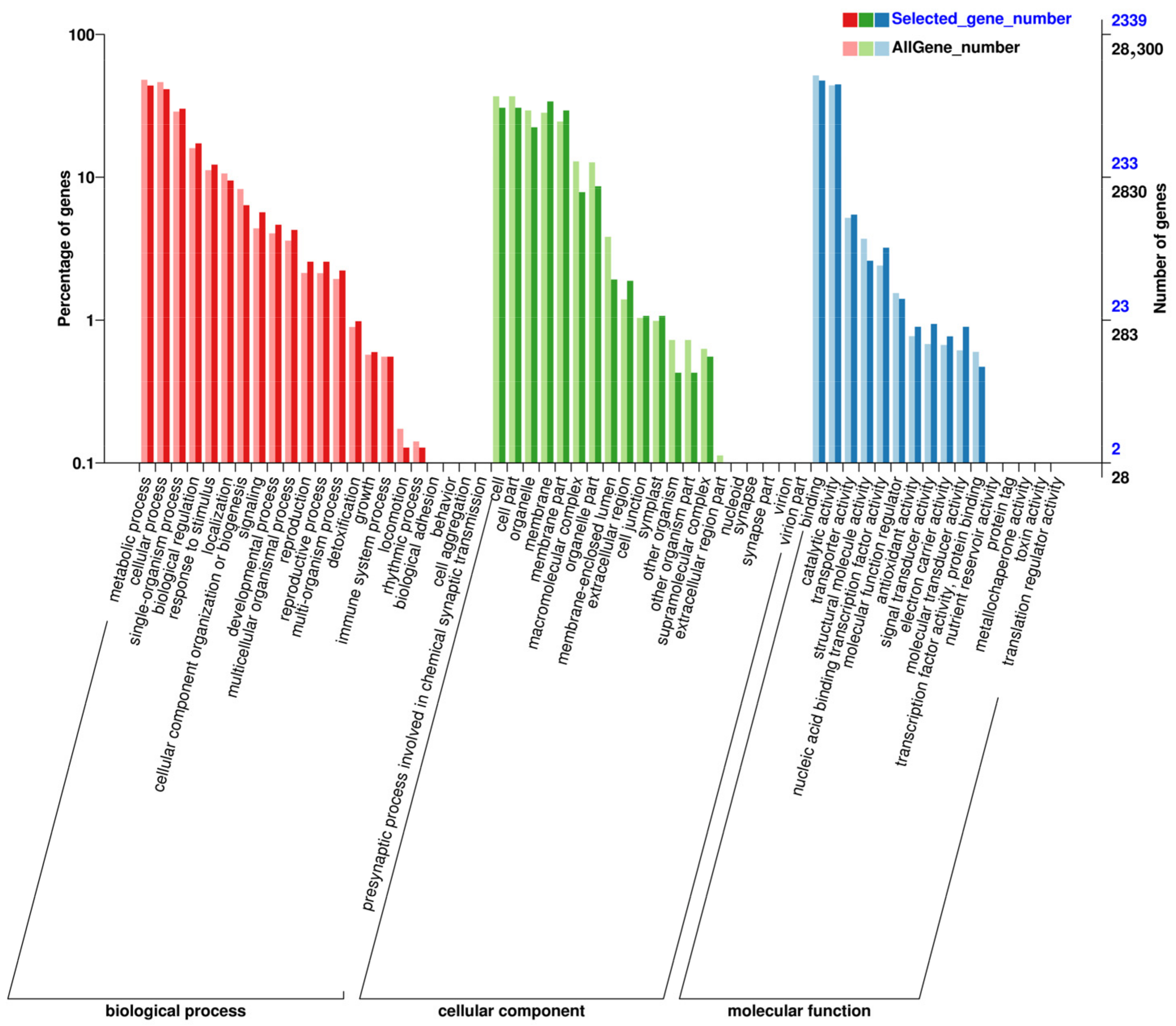
3.6. Analysis of Differentially Expressed Gene Functions Using GO Annotation
3.7. KEGG Pathway Analysis of Differentially Expressed Genes
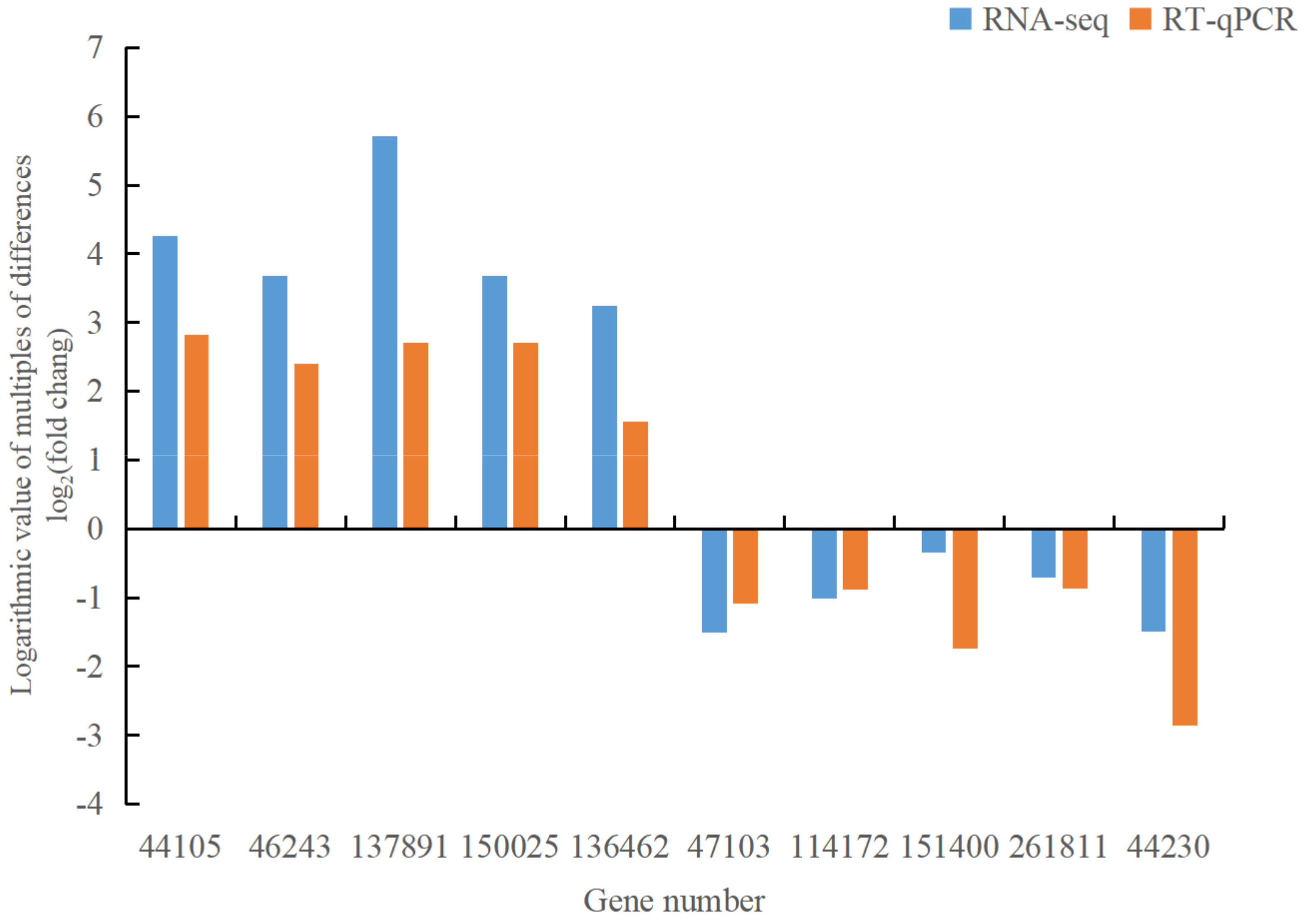
3.8. Validation of Differentially Expressed Genes by RT-qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, X.L. Phylogenetic Analysis of Fallen-Needle Disease and Its Pathogen in Artificial Spruce Forests in Western Sichuan Province, China. Ph.D. Thesis, Sichuan Agricultural University, Chengdu, China, 2016. (In Chinese). [Google Scholar]
- Deng, Q.Q.; Wang, X.; Yan, X.J. Progress in the application of transcriptome and metabolome in defense response to forest fungal diseases. World For. Res. 2022, 35, 27–32. (In Chinese) [Google Scholar]
- Tan, J.L.; Ma, M.G.; Che, T.; Bai, Y.J. Research on the canopy interception characteristics of Picea crassifolia based on different depression. Prog. Earth Sci. 2009, 24, 825–833. (In Chinese) [Google Scholar]
- Zhao, D.C. The occurrence rule and prevention agent screening of Picea crassifolia rust in Qilian Mountain Nature Reserve. Mod. Gard. 2024, 47, 49–51. (In Chinese) [Google Scholar]
- Huo, H.C.; Yang, F.; Zhang, R.K.; Xu, H.X.; Cao, X.W. Progress in spruce cone rust disease. Gansu For. Sci. Technol. 2022, 47, 27–33. (In Chinese) [Google Scholar]
- Ma, M.R. Application of pollution-free technology in the control of spruce rust. South. Agric. 2021, 15, 72–73. (In Chinese) [Google Scholar]
- Zhao, H.H. Pathogen rule and control of leaf rust in Picea crassifolia. Peasant. Staff. Memb. 2021, 11, 163–164. (In Chinese) [Google Scholar]
- Cai, C.L. Law and preliminary prevention measures of spruce leaf rust in Taohe Forest Area. Gansu For. 2017, 2, 41–42. (In Chinese) [Google Scholar]
- Niu, H. Screening and Functional Characterization of Genes Regulating Abnormal Stomatal Development in Wheat. Master’s Thesis, Tianjin Agricultural College, Tianjin, China, 2023. (In Chinese). [Google Scholar]
- Shang, Y.F. Research on the Mechanism of Abnormal Stomatal Formation and Development in Wheat. Master’s Thesis, Tianjin Agricultural College, Tianjin, China, 2021. (In Chinese). [Google Scholar]
- Liu, Y.Q.; Liu, L.N.; Chen, Y.; Li, C.Y.; Yang, J. Analysis of plant pathogenic fungal effector proteins and plant interaction dynamics. Mol. Plant Breed. 2018, 16, 6678–6687. (In Chinese) [Google Scholar]
- Que, Y.X.; Song, X.X.; Xu, L.P.; Chen, R.K. Progress in the interaction mechanism between plants and pathogenic fungi. Biotechnol. Commun. 2009, 20, 282–285. (In Chinese) [Google Scholar]
- Ni, N.N.; Yang, X.N.; Zhao, J.H.; Li, Z.Y.; Qiao, L.H.; Li, G.W.; Wang, D.W.; Wang, H. Initial mapping of stripe rust resistance genes in Uraltu wheat based on BSA-seq. J. Henan Agric. Univ. 2024, 58, 44–51. (In Chinese) [Google Scholar]
- Zhao, J. Physiological Response Mechanism of Pear Fruit Callus to Rot Fungal Infection. Ph.D. Thesis, Northwest A & F University, Xianyang, China, 2012. (In Chinese). [Google Scholar]
- Huang, Y.C.; Ren, D.L.; He, B.; Zhao, Y.M.; Gong, X.J.; Chen, J.X.; Liu, L.Y. Research progress of transcriptomics and metabolomics in plant abiotic stress. Jiangsu Agric. Sci. 2023, 51, 1–7. (In Chinese) [Google Scholar]
- Ye, X.L.; Li, X.G.; Chen, S.H.; Wang, Q. Studies on the effects of surfactants on the stomata and wax layer of soybean leaves. Soybean Sci. 2000, 8, 49–56. (In Chinese) [Google Scholar]
- Li, N. Mining and Validation of Key Genes for Wax Synthesis in Jujube Fruit. Ph.D. Thesis, Shanxi Agricultural University, Taiyuan, China, 2021. (In Chinese). [Google Scholar]
- Qin, S.W.; Yan, W.W.; Li, C.L.; Wu, H.Z.; He, Z.G.; Qiu, X.; Zhao, L.F. Functional analysis of plant signaling pathway-related genes in heteromorphic leaf genesis of poplar based on triple sequencing technology. Mol. Plant Breed. 2023, 21, 6688–6697. (In Chinese) [Google Scholar]
- Fan, W.Q.; Liu, X.X.; Ma, X.L.; Yang, H.; Zhu, J.Z.; Tang, J.N.; Yue, S.J.; Zheng, R. Mining of male sterility-related genes in Lycium barbarum using transcriptomic data. Chin. Herb. Med. 2023, 54, 2226–2234. (In Chinese) [Google Scholar]
- Li, Y.G.; Li, Y.; Liu, W.W.; Yang, M.X.; Wen, J.Z. Expression analysis of differentially displayed genes in the early stage of nonaffinity intercropping between soybean and Mycosphaerella spp. J. Plant Prot. 2011, 38, 413–418. (In Chinese) [Google Scholar]
- Yin, X.M.; Lian, Z.H.; Liu, Y.X.; Wang, L.X.; Chen, D.; He, Y.D. Correlation between banana leaf structure and the occurrence of brown-margined gray spot disease in different genotypes. Trop. Agric. Sci. 2022, 42, 27–32. (In Chinese) [Google Scholar]
- Xu, Y.Y.; Luo, G.M.; Luo, Y.J.; Liu, P.P.; Wang, E.Y.; Chen, Y.T.; Zhou, L. Scanning electron microscopic observation on the infestation of gardenia leaves by Phytophthora septoria. J. Electron Microsc. 2021, 40, 441–445. (In Chinese) [Google Scholar]
- Xuan, Z.; Che, H.Y.; Cao, X.R.; He, Y.H.; Luo, D.Q. Analysis of common differentially expressed genes for leaf yellowing in betel nut based on transcriptome sequencing. J. Trop. Crops 2021, 42, 2834–2840. (In Chinese) [Google Scholar]
- Fang, T.J.; Hou, L.; Bai, L.C. Research on the pathogenesis of root rot of Lycium barbarum in Qaidam region based on transcriptome sequencing. Arid. Zone Resour. Environ. 2022, 36, 133–140. (In Chinese) [Google Scholar]
- Li, M.X. Transcriptome Analysis of Walnut in Response to Xanthomonas Infection and Disease Resistance. Master’s Thesis, Northwest A & F University, Xianyang, China, 2023. (In Chinese). [Google Scholar]
- Wang, T.T.; Chen, G.; Bao, Y.L.; Jin, D.C. Transcriptome analysis of soybean root system in response to early iron deficiency stress. J. Nanjing Agric. Univ. 2022, 45, 224–234. (In Chinese) [Google Scholar]
- Que, Y.X.; Song, Q.Q.; Xu, L.P.; Chen, R.K. Progress in the study of plant-pathogen fungus interactions. Biotechnol. Lett. 2009, 20, 282–285. (In Chinese) [Google Scholar]
- Li, C.L.; Niu, L.J.; Hu, L.L.; Liao, W.B.; Chen, Y. Effects of calcium ions on nitric oxide-induced adventitious rootogenesis in cucumber under drought conditions. J. Appl. Ecol. 2017, 28, 3619–3626. (In Chinese) [Google Scholar]
- Li, S. Mechanism of RDR1 Gene in Plant Defense against Virus-like Viruses. Ph.D. Thesis, Southwest University, Chongqing, China, 2022. (In Chinese). [Google Scholar]
- Sang, S.L. Regulatory Effects of Extrtoplasmic Hydrogen Peroxide and Aquaporin PIP1; 4 on the Induction of Hpa 1 Protein in Arabidopsis. Ph.D. Thesis, Agricultural University of Nanjing, Nanjing, China, 2011. (In Chinese). [Google Scholar]
- Jia, F.L. Analysis of Differential Expression of Genes Responding to Hydrogen Peroxide Stress in Poplar Leaf Blight Pathogens. Master’s Thesis, Northeast Forestry University, Harbin, China, 2022. (In Chinese). [Google Scholar]
- Liu, N.; Qiao, M.; Sun, J.W.; Chen, Y.; Hou, C.Y.; Han, S.F.; Wang, D.M. Analysis of transcriptome data from leaf rust-infested wheat leaves. J. Plant Genet. Resour. 2019, 20, 991–1000. (In Chinese) [Google Scholar]
- Feng, Y.T.; Li, J.Y.; Tang, L.; Niu, X.L.; Chen, Y.H.; Luo, K. Identification of cassava GH3 gene family and its expression analysis in response to pathogen infestation. Mol. Plant Breed. 2022, 20, 1755–1765. (In Chinese) [Google Scholar]
- Zhao, W.; Zhao, L.; Zhang, J.P.; Qi, Y.N.; Wang, L.M.; Xie, Y.P.; Li, W.J.; Dang, Z.; Yuan, M.L.; Zhang, Y.P. Joint transcriptomic and metabolomic analysis of the response mechanism of Huperzia serrata roots to salt stress. Grass Sci. 2022, 39, 1151–1164. (In Chinese) [Google Scholar]
- Li, Z.; Yang, Y.Y.; Dai, L.Y.; Zhang, F.F.; A, X.X.; Dong, C.; Wang, B.; Tang, C.F. Progress in the research and utilization of rice leaf blight resistance genes and related factors. Chin. Agron. Bull. 2022, 38, 91–99. (In Chinese) [Google Scholar]
- Liu, D.X.; Wang, Y.P.; Wu, J. Role of WRKY transcription factors in phytohormone-mediated disease resistance pathways. Mol. Plant Breed. 2023, 1–7. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20220512.0937.010.html (accessed on 21 April 2024). (In Chinese).
- He, D.; Li, P.; Zhao, S.; Zhao, T.T.; Tian, M.; Jang, H. Role of phytohormones in rice blast fungus-rice pathosystem. North. Rice 2023, 53, 55–58. (In Chinese) [Google Scholar]
- Chen, L.; Lin, Y.; Chen, P.F.; Wang, S.H.; Ding, Y.F. Proteomic analysis of rice phloem sap in response to iron deficiency. J. Bot. 2019, 54, 194–207. (In Chinese) [Google Scholar]
- Li, Y.; Zuo, K.J.; Tang, K.X. Overview of functional studies on plant GH3 gene family. Bot. Bull. 2008, 25, 507–515. (In Chinese) [Google Scholar]
- Zhang, C. Identification of the GH3 Family of Jasmonic Acid-Regulated Genes and Analysis of Disease Resistance and Damage in Potato. Ph.D. Thesis, Northwest Agriculture and Forestry University, Xianyang, China, 2021. (In Chinese). [Google Scholar]
- Hu, K.; Li, N. Carotenoids, the favored antioxidant family. Univ. Chem. 2010, 25 (Suppl. S1), 94–98. (In Chinese) [Google Scholar]
- He, F.; Ding, M.; Zheng, H.N.; Chen, Y.J.; Xue, X.X. Paeoniflorin inhibits IL-13-induced oxidative stress and autophagy [J/OL] in BEAS-2B cells via STAT 3. Chin. J. Hosp. Pharm. 2024, 1–8. Available online: http://kns.cnki.net/kcms/detail/42.1204.R.20240312.1006.006.html (accessed on 21 April 2024). (In Chinese).
- Chai, P.P. Identification of Aspergillus Flavus Resistance in Peanut Germplasm and Analysis of Resistance-Related PAL Gene Family in Henan Province. Master’s Thesis, Zhengzhou University, Zhengzhou, China, 2021. (In Chinese). [Google Scholar]
- Xu, Q. Research on the molecular mechanism of powdery mildew resistance in prickly pear. J. Huazhong Agric. Univ. 2011, 30, 124–127. (In Chinese) [Google Scholar]
- Zhang, Y. Study of the Mechanism of Enhancing Salt Tolerance and Cadmium Stress Caused by Melatonin Seed. Master’s Thesis, Zhejiang University, Hangzhou, China, 2022. (In Chinese). [Google Scholar]
- Zhao, S.P. Analysis of Genes Related to Systemic Infection and Induced Resistance in Cucumber Flow Gum Disease. Ph.D. Thesis, Agricultural University of Hebei, Baoding, China, 2021. (In Chinese). [Google Scholar]
- Wang, N. Transcriptome and Expression Profiling Study and Related Gene Expression Analysis of Cereal Grains Interacting with Rust Fungus. Master’s Thesis, Hebei Normal University, Shijiazhuang, China, 2015. (In Chinese). [Google Scholar]
- Xie, K.Z.; Qiu, H.Z.; Yue, Y.; Hu, X.Y.; Luo, A.H.; Liu, Y.Q.; Sun, X.H.; Tang, D.J. Effects of Fusarium spinosum infestation on photosynthetic efficiency and chlorophyll fluorescence parameters of potato. J. Plant Prot. 2022, 49, 927–937. (In Chinese) [Google Scholar]
- Zheng, J.Y. Screening of Potato High Resistance to Late Blight Resources and Mining of Disease Resistance Related Genes. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. (In Chinese). [Google Scholar]
- Wang, S.Y.; Sun, Z.G.; Tian, H. Effects of exogenous melatonin on photosynthesis and physiological indexes of alfalfa under salt stress. Heilongjiang Anim. Husb. Vet. Med. 2023, 5, 12–16+21. (In Chinese) [Google Scholar] [CrossRef]
- Liu, J.C.; Su, D.Y. Effects of high temperature on chloroplast ribosomes and chloroplast protein biosynthesis in wheat. J. Bot. 1985, 27, 63–67. (In Chinese) [Google Scholar]
- Dong, J.X.; Dong, H.T.; Li, D.B. Progress in the study of plant disease resistance genes. J. Plant Pathol. 2001, 31, 1–9. (In Chinese) [Google Scholar]
- Wang, L.F. Research on Physiological Mechanism and Ultrastructure of Rust Resistance in Sunflower. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2008. (In Chinese). [Google Scholar]
- Zhang, R.J. Effects of Different Pretreatment on Photosynthetic Apparatus of Rice under Water Stress. Master’s Thesis, Shenyang Normal University, Shenyang, China, 2012. (In Chinese). [Google Scholar]
- Si, S.W. Classification of Cucumber Resistance to Downy Mildew Types Based on Allergic Reactions. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2011. (In Chinese). [Google Scholar]
- Gu, Z.F.; Wang, W.Q.; Zhu, A.P.; Zhu, X.M.; He, H.L.; Pan, J.S.; Cai, R. Correlation of cucumber resistance to downy mildew with chlorophyll content and stomatal density. J. Shanghai Jiao Tong Univ. 2004, 4, 381–384. (In Chinese) [Google Scholar]
- Zhao, Z.L.; Liu, Q.N.; Su, L.P. Methods of in vitro larval characterization of resistance to faba bean erythroplasma. J. Yunnan Agric. Univ. 1995, 2, 120–122. (In Chinese) [Google Scholar]
- Chai, C.B. Evaluation of Pear Resistance to Black Star Disease and Research on Physiological Mechanisms. Master’s Thesis, Shanxi Agricultural University, Jinzhong, China, 2015. (In Chinese). [Google Scholar]
- Liu, Z.L. Study on Sugarcane-Sugarcane Whip-Blackfly Interactions and Localization of Sugarcane Black Sigatoka Quantitative Trait Loci. Ph.D. Thesis, Guangxi University, Nanning, China, 2023. (In Chinese). [Google Scholar]
- Ci, E.J.M.; Wang, H.P.; Gou, X.Y.; Tong, K.X.; Guo, R.C.; Wng, J.B.; Cao, G.X.; Bao, S.G.; Xie, X.D.; Chen, X.Q. Cloning and preliminary functional analysis of barley stomatal development-related gene HvSPCH. J. Wheat Crops 2022, 42, 1175–1181. (In Chinese) [Google Scholar]
- Chen, S.C. Preliminary Study on the Function of Rice Stomatal Development-Related Genes. Master’s Thesis, Henan University, Kaifeng, China, 2015. (In Chinese). [Google Scholar]
- Chen, Q.Y. Functional Study on the Regulation of Stomatal Development by STOMAGEN-Like Genes in Maize. Master’s Thesis, Guangxi University, Nanning, China, 2017. (In Chinese). [Google Scholar]
- Lv, X.C.; Zheng, H.; Zhang, W.Y.; Lu, X.L.; Fang, Y.X.; Chen, J.M.; Xue, D.W.; Zhang, X.Q. Research on physiology and wax gene expression of different varieties of barley under drought stress. J. Hangzhou Norm. Univ. 2019, 18, 135–140. (In Chinese) [Google Scholar]
- Zhou, X.Y. Cloning and Characterization of Rice Leaf Epidermal Wax Development and Wax-Related Transcription Factor Genes OsWTF1 and OsWTF2. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2007. (In Chinese). [Google Scholar]
- Tian, L.B. Construction of Genetic Map and QTL Localization of Powdery Mildew Resistance in Bitter Melon. Ph.D. Thesis, Shenyang Agricultural University, Shenyang, China, 2015. (In Chinese). [Google Scholar]
- Wang, Z. Premium Research on the Causes and Pathogenesis of Ginkgo Yellowing Disease. Master’s Thesis, Liaoning Normal University, Dalian, China, 2008. (In Chinese). [Google Scholar]
- Feng, L.Z. Research on the Resistance Mechanism of Eucalyptus Species Lines to Scorch Disease. Ph.D. Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2008. (In Chinese). [Google Scholar]
- Wu, R.J.; Wen, X.P.; Li, Z.C.; Mei, L.N.; Shen, L.N.; Wu, Z.Y. Cloning and expression analysis of sucrose phosphate synthase gene in Pinus massoniana. J. For. Environ. 2022, 42, 199–207. (In Chinese) [Google Scholar]
- Tong, T.; Fang, Y.X.; Zhang, X.Q.; Xue, D.W. Progress in the study of plant epidermal wax properties and related genes. J. Hangzhou Norm. Univ. (Nat. Sci. Ed.) 2019, 18, 610–618. (In Chinese) [Google Scholar]
- Li, L.; Zhao, M.X.; Wang, J.H.; Liu, S.Z.; Wang, G.Y. Progress of research on the synthesis, transportation and regulatory mechanism of plant epidermal waxes. J. China Agric. Univ. 2023, 28, 1–19. (In Chinese) [Google Scholar]
- Wang, T.Y. Molecular Mechanism Analysis of Arabidopsis Thaliana Histone Acetyltransferase GCN5 Regulating Seed Fatty Acid and Stem Epidermal Wax Synthesis. Ph.D. Thesis, China Agricultural University, Beijing, China, 2016. (In Chinese). [Google Scholar]
- Xue, B.; Tian, X.; Xu, C.L.; Yang, Y.M.; Wang, D.X.; Hu, Y.X.; Su, J.E. Bioinformatics analysis of tobacco caffeic acid 3-O-methyltransferase. Tianjin Agric. Sci. 2019, 25, 5–8. (In Chinese) [Google Scholar]
- Fang, T.J.; Bai, L.C. Analysis of the community structure and diversity of endophytic fungi in the needles of Picea crassifolia. Mol. Plant Breed. 2022, 20, 5198–5204. (In Chinese) [Google Scholar]
- Wen, T.T.; Wang, Y.; Li, Z.; Lin, C.; Guan, Y.J.; Hu, J. Effects of seed coat structure and composition on seed water permeability in pea. Seed 2016, 35, 19–25. (In Chinese) [Google Scholar]
- Zhao, H.Y.; Wei, J.H.; Song, Y.R. Progress in lignin biosynthesis and its genetic engineering. J. Plant Physiol. Mol. Biol. 2004, 30, 361–370. (In Chinese) [Google Scholar]
- Li, B.; Ni, Z.Y.; Wang, J.; Lv, M.; Fan, L. Progress of caffeic acid-O-methyltransferase gene (COMT), a key enzyme for lignin biosynthesis. Mol. Plant Breed. 2010, 8, 117–124. (In Chinese) [Google Scholar]
- Tong, Q. Expression Characterization and Functional Study of Phenylalanine Deaminase Gene in Xing’an Larch; Inner Mongolia University: Hohhot, China, 2023. (In Chinese) [Google Scholar]
- Li, Y. Transcriptional and Metabolic Analysis of Lignin Biosynthesis in Sidecarpin. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2022. (In Chinese). [Google Scholar]
- Liu, H.J.; Wang, X.Z.; Wang, L.F.; Xu, L.Y.; Li, C.L.; Cao, Y.F. Study on the changes of enzyme activities in resistant family lines of Masson pine after infection with pine wood nematode. J. Southwest For. Univ. (Nat. Sci.) 2018, 38, 107–111. (In Chinese) [Google Scholar]
- Li, J. Analysis of Lignin Fractions during Infestation of Kiwifruit Ulcer Disease. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2022. (In Chinese). [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Primer Name | Sequences (5′ to 3′) | Clip Size |
|---|---|---|---|
| 137891 | s137891-F | F: GAAATTAGCTCCCGTGTTTC | 176 |
| s137891-R | R: GCTCTCTACAGGAAAATCAC | ||
| 46243 | s046243-F | F: TATGTTCAACCCGTCAAATC | 183 |
| s046243-R | R: CAGTGGATGATCCAGTTGAG | ||
| 150025 | s150025-F | F: TAGAAGAAAGAGGACCAGGG | 186 |
| s150025-R | R: CTGCAGCTAGTACCTTTCCC | ||
| 44105 | s044105-F | F: CTTCTCTTCCGCTCATTAAC | 198 |
| s044105-R | R: CTGTGGATTGCTCTGATACC | ||
| 136462 | s136462-F | F: TGCTGCTGCTACGAGACTTC | 191 |
| s136462-R | R: TGTTACAGTTGGTCCCAACG | ||
| 261811 | x261811-F | F: ATGACAGTCTTAAACAGCGG | 191 |
| x261811-R | R: AGGACTGTTCGATTGATCTC | ||
| 114172 | x114172-F | F: TTAGTAACACAAAGAAGCCG | 179 |
| x114172-R | R: GAGGATGCCAAATTAGCTTC | ||
| 151400 | x151400-F | F: ATTTCGATTCTCTTCTGGTC | 197 |
| x151400-R | R: GGATGTTATCCATGGGATTC | ||
| 47103 | x047103-F | F: TAATACGCCCTTCCTCTATC | 182 |
| x047103-R | R: CAACGCGACAGATTAAAAAG | ||
| 44230 | x044230-F | F: CATTAACATACACAGCTCCC | 183 |
| x044230-R | R: TAGACCAGTTGTGGGTATGC | ||
| SKI2 | SKI2-F | F: CACGCCTTCACAGCAATCAG | 291 |
| SKI2-R | R: TCCTGTAGCCCTCTTCCATCA |
| Reaction Component | Concentration | Volume (µL) |
|---|---|---|
| ChamQ SYBR Color qPCR Master Mix | 2× | 10 |
| Primer F | 5 µM | 0.4 |
| Primer R | 5 µM | 0.4 |
| Template (DNA) | 2 | |
| ddH2O | 7.2 | |
| Total | 20 µL |
| Number | Instrument Name | Model | Manufacturer |
|---|---|---|---|
| 1 | PCR instrument | MG96+ | Hangzhou Langji Scientific Instrument Co., Ltd., Hangzhou, China |
| 2 | Double stable time electrophoresis instrument | dyy-6c | Beijing Junyi Oriental Electrophoresis Equipment Co., Ltd., Beijing, China |
| 3 | Hole centrifuge | H1650 | Hunan Changsha Xiangyi Centrifuge Instrument Co., Ltd., Changsha, China |
| 4 | Precision pipette | a suit of | Eppendorf Co., Hamburg, Germany |
| 5 | Vortex mixer | MixMax | Hefei Ebenson Scientific Instrument Co., Ltd., Hefei, China |
| 6 | Gel imager | JY04S-3C | Beijing Junyi Oriental Electrophoresis Equipment Co., Ltd., Beijing, China |
| 7 | Fluorescence quantitative PCR instrument | 9600plus | Hangzhou Bori Technology Co., Ltd., Hangzhou, China |
| 8 | Ultra-trace visible light ultraviolet spectrophotometer | ND5000 | Beijing Baitech Biotechnology Co., Ltd., Beijing, China |
| Number | Name of Reagent or Consumable Material | Model/Type | Supplier |
|---|---|---|---|
| 1 | 2X ChamQ SYBR, COLOR qPCR Master Mix | Q411-02/03 | Nanjing Nuoweizan Biotechnology Co., Ltd., Nanjing, China |
| 2 | PCR primer | PAGE purification | Shenggong Bioengineering Co., Ltd., Shanghai, China |
| 3 | Eight even tube | PCR-0208-C | Aisijin Biotechnology (Hangzhou) Co., Ltd., Hangzhou, China |
| 4 | DL2000 | DL2501 | Shanghai Jierui Biological Engineering Co., Ltd., Shanghai, China |
| 5 | EX Taq enzyme | RR001 | Baori Doctor Material Technology (Beijing) Co., Ltd. (TaKaRa), Beijing, China |
| 6 | dNTP | BK6501A | Baori Doctor Material Technology (Beijing) Co., Ltd. (TaKaRa), Beijing, China |
| 7 | 10 × EX Taq Buffer | AB5401A | Baori Doctor Material Technology (Beijing) Co., Ltd. (TaKaRa), Beijing, China |
| 8 | Primer | Shenggong Bioengineering (Shanghai) Co., Ltd., Shanghai, China | |
| 9 | HiScript Q RT SuperMix for qPCR (+gDNA wiper) | R123-01 | Nanjing Nuoweizan Biotechnology Co., Ltd., Nanjing, China |
| 10 | Soil RNA Kit | R6825-02 | Omega Bio-Tek, Norcross, GA, USA |
| 11 | OminiPlant RNA, Kit(DNase I) | CW2598S | Beijing Kangwei Century Biotechnology Co., Ltd., Beijing, China |
| 12 | E.Z.N.A.® HP Tota RNA, Kit | R6812-01 | Omega Bio-Tek, Norcross, GA, USA |
| Brochure | Read Number | Base Number | GC (%) | %≥Q30 |
|---|---|---|---|---|
| N1 | 27,757,413 | 8,310,361,054 | 44.91% | 94.48% |
| N2 | 28,723,680 | 8,599,171,640 | 43.83% | 94.23% |
| N3 | 26,402,330 | 7,901,675,174 | 44.58% | 94.72% |
| L1 | 27,757,413 | 8,310,361,054 | 44.91% | 93.26% |
| L2 | 28,723,680 | 8,599,171,640 | 43.83% | 93.38% |
| L3 | 26,402,330 | 7,901,675,174 | 44.58% | 93.01% |
| B1 | 20,530,290 | 6,146,385,256 | 47.86% | 94.46% |
| B2 | 21,433,171 | 6,417,397,574 | 47.80% | 93.80% |
| B3 | 21,080,933 | 6,311,108,490 | 48.67% | 93.90% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Bai, L. Transcriptome Analysis of Picea crassifolia in Response to Rust Infestation. J. Fungi 2024, 10, 313. https://doi.org/10.3390/jof10050313
Li H, Bai L. Transcriptome Analysis of Picea crassifolia in Response to Rust Infestation. Journal of Fungi. 2024; 10(5):313. https://doi.org/10.3390/jof10050313
Chicago/Turabian StyleLi, Hailan, and Luchao Bai. 2024. "Transcriptome Analysis of Picea crassifolia in Response to Rust Infestation" Journal of Fungi 10, no. 5: 313. https://doi.org/10.3390/jof10050313
APA StyleLi, H., & Bai, L. (2024). Transcriptome Analysis of Picea crassifolia in Response to Rust Infestation. Journal of Fungi, 10(5), 313. https://doi.org/10.3390/jof10050313