Molecular Identification, Antifungal Susceptibility, and Geographic Origin of Clinical Strains of Sporothrix schenckii Complex in Mexico

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
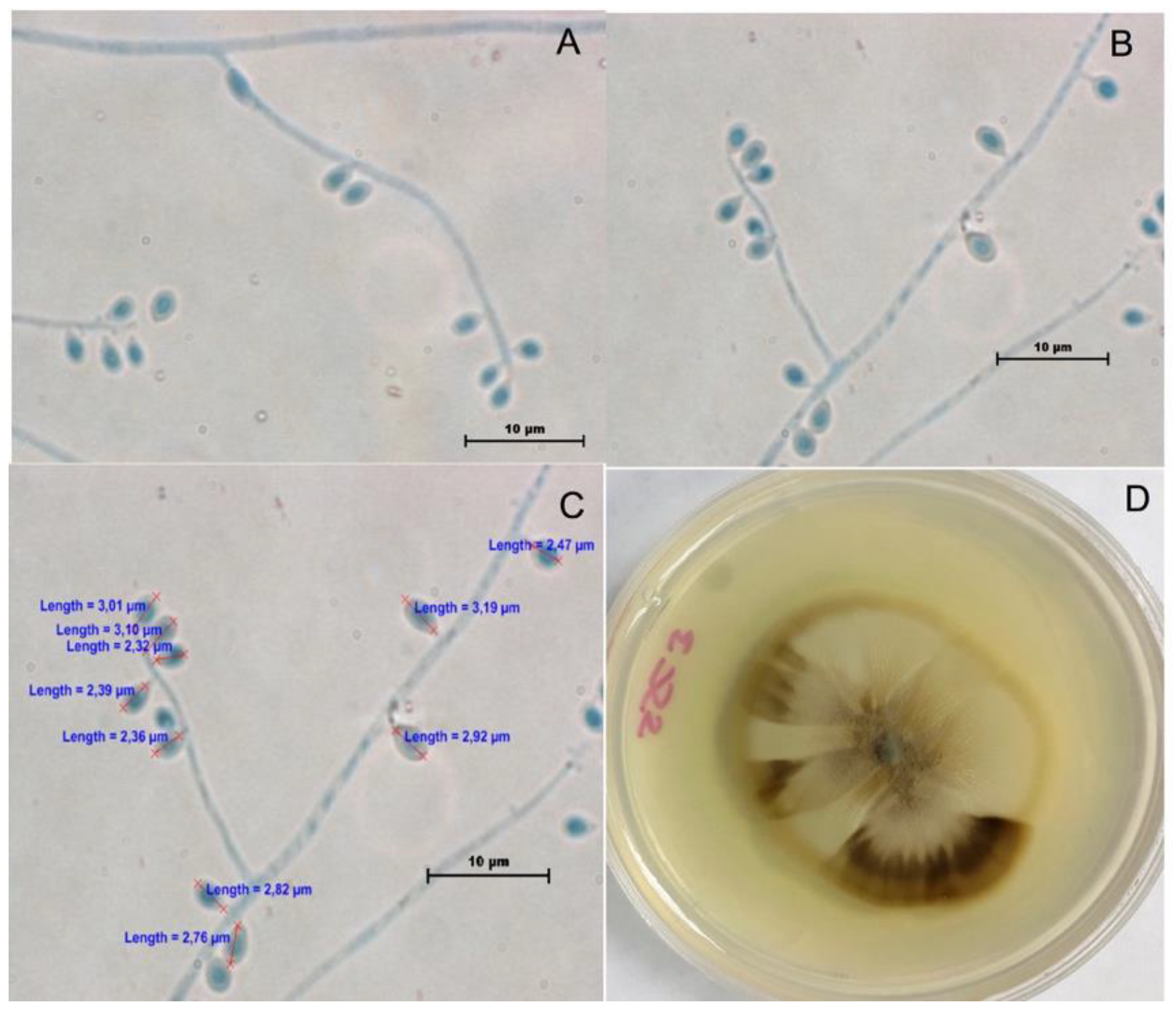
2.1. Phenotypic Characterization
2.2. Antifungal Susceptibility
2.3. Molecular Identification
3. Results
3.1. Phenotypic Characterization
3.2. Antifungal Susceptibility
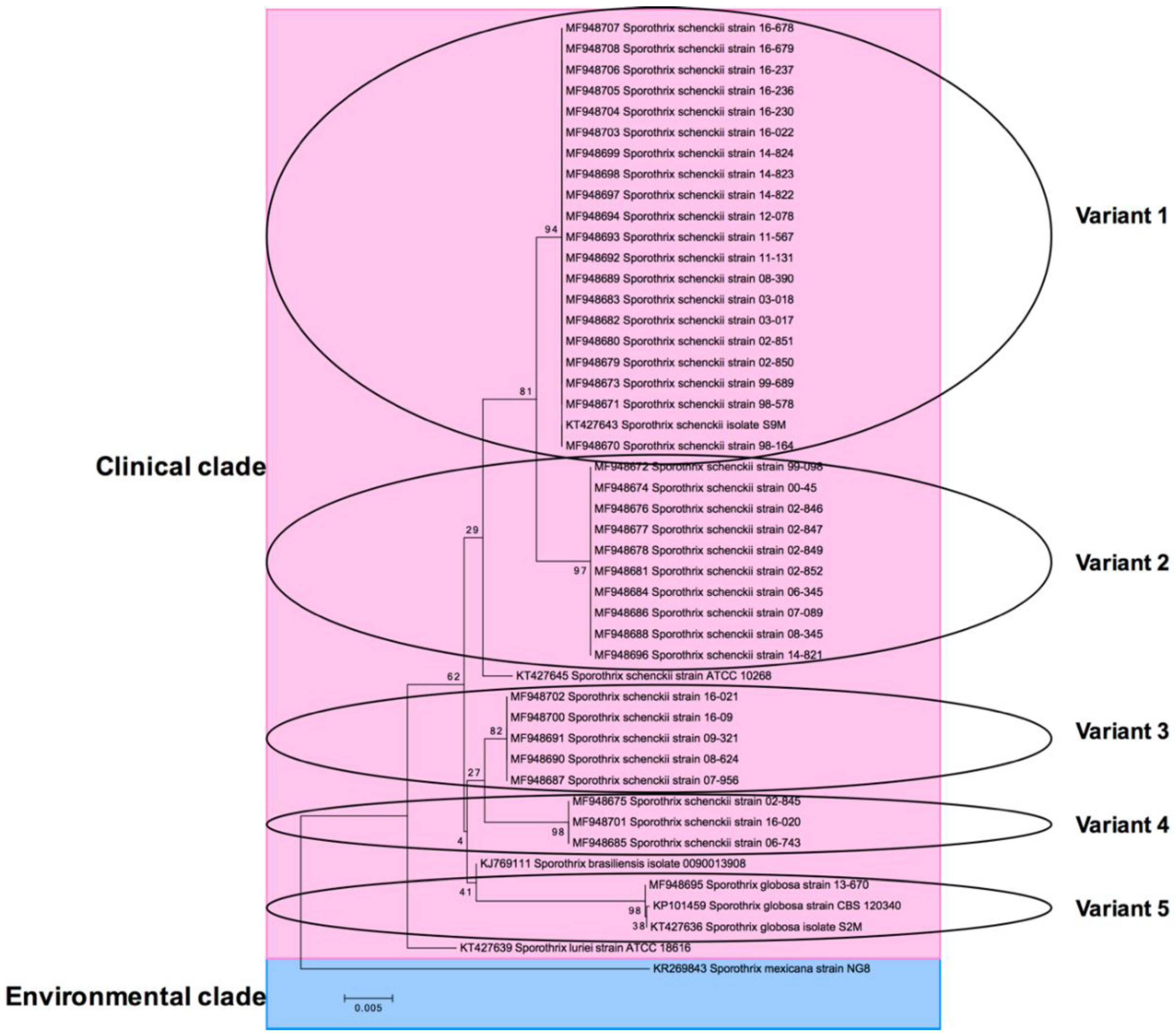
3.3. Molecular Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oliveira, M.M.; Almeida-Paes, R.; Gutierrez-Galhardo, M.C.; Zancope-Oliveira, R.M. Molecular identification of the Sporothrix schenckii complex. Rev. Iberoam. Micol. 2014, 31, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Melo Teixeira, M.; de Hoog, G.S.; Pacheco Schubach, T.M.; Pereira, S.A.; Fernandes, G.F.; Lopes Bezerra, L.M.; Felipede Camargo, M.S.; de Camargo, Z.P. Phylogenetic analysis reveals a high prevalence of Sporothrix brasiliensis in feline sporotrichosis outbreaks. PLoS Negl. Trop. Dis. 2013, 7, e2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Wan, Z.; Zhang, Z.; Li, F.; Li, R.; Liu, X. Phenotypic and molecular identification of Sporothrix isolates of clinical origin in Northeast China. Mycopathologia 2013, 176, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Yikelamu, A.; Tanaka, R.; Igawa, K.; Yokozeki, H.; Yaguchi, T. Studies in Phylogeny, Development of Rapid IdentificationMethods, Antifungal Susceptibility, and Growth Rates of Clinical Strains of Sporothrix schenckii Complex in Japan. Med. Mycol. J. 2016, 57, E47–E57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hagen, F.; Stielow, B.; Rodrigues, A.M.; Samerpitak, K.; Zhou, X.; Feng, P.; Yang, L.; Chen, M.; Deng, S.; et al. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia 2015, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Del Rocio Reyes-Montes, M.; Perez-Mejia, A.; Navarro-Barranco, H.; Souza, V.; Zuniga, G.; Toriello, C. Phenotyping and genotyping of Sporothrix schenckii isolates according to geographic origin and clinical form of Sporotrichosis. J. Clin. Microbiol. 2002, 40, 3004–3011. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Gamboa, L.; Martinez-Hernandez, F.; Maravilla, P.; Flisser, A. A population genetics analysis in clinical isolates of Sporothrix schenckii based on calmodulin and calcium/calmodulin-dependent kinase partial gene sequences. Mycoses 2018. [Google Scholar] [CrossRef] [PubMed]
- Ottonelli Stopiglia, C.D.; Magagnin, C.M.; Castrillon, M.R.; Mendes, S.D.; Heidrich, D.; Valente, P.; Scroferneker, M.L. Antifungal susceptibilities and identification of species of the Sporothrix schenckii complex isolated in Brazil. Med. Mycol. 2014, 52, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Marimon, R.; Gene, J.; Cano, J.; Trilles, L.; Dos Santos Lazera, M.; Guarro, J. Molecular phylogeny of Sporothrix schenckii. J. Clin. Microbiol. 2006, 44, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, A.; Tirado-Sanchez, A.; Paredes-Solis, V.; Cepeda-Valdes, R.; Gonzalez, G.M.; Trevino-Rangel, R.J.; Fierro-Arias, L. Cutaneous disseminated sporotrichosis: Clinical experience of 24 cases. J. Eur. Acad. Dermatol. Venereol. 2018, 32, e77–e79. [Google Scholar] [CrossRef] [PubMed]
- Marimon, R.; Cano, J.; Gene, J.; Sutton, D.A.; Kawasaki, M.; Guarro, J. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical interest. J. Clin. Microbiol. 2007, 45, 3198–3206. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; McGinnis, M.R.; Pincus, D.H.; Goldson, P.R.; Kerkering, T.M. Evaluation of the API 20C yeast identification system for the differentiation of some dematiaceous fungi. J. Clin. Microbiol. 1989, 27, 2565–2569. [Google Scholar] [PubMed]
- John, H.R.; Barbara, D.A.; David, A.; Beth, A.S.; Steven, D.B.; Vishnu, C. M38-A2: Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, Approved Standard, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2002; Volume 28. [Google Scholar]
- Gonzalez, G.M.; Rojas, O.C.; Bocanegra-Garcia, V.; Gonzalez, J.G.; Garza-Gonzalez, E. Molecular diversity of Cladophialophora carrionii in patients with chromoblastomycosis in Venezuela. Med. Mycol. 2013, 51, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A Multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Madrid, H.; Cano, J.; Gene, J.; Bonifaz, A.; Toriello, C.; Guarro, J. Sporothrix globosa, a pathogenic fungus with widespread geographical distribution. Rev. Iberoam. Micol. 2009, 26, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cui, Y.; Zhen, Y.; Yao, L.; Shi, Y.; Song, Y.; Chen, R.; Li, S. Genetic variation of Sporothrix globosa isolates from diverse geographic and clinical origins in China. Emerg. Microbes. Infect. 2017, 6, e88. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; Leon-Navarro, I.; Rodriguez-Brito, S.; Mendoza, M.; Nino-Vega, G.A. Molecular epidemiology of human sporotrichosis in Venezuela reveals high frequency of Sporothrix globosa. BMC Infect. Dis. 2015, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Kohler, L.M.; Hamdan, J.S.; Ferrari, T.C. Successful treatment of a disseminated Sporothrix schenckii infection and in vitro analysis for antifungal susceptibility testing. Diagn. Microbiol. Infect. Dis. 2007, 58, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Zaini, F.; Kordbacheh, P.; Safara, M.; Heidari, M. Sporothrix schenckii complex in Iran: Molecular identification and antifungal susceptibility. Med. Mycol. 2016, 54, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, S.; Isla, G.; Szusz, W.; Vivot, W.; Hevia, A.; Davel, G.; Canteros, C.E. Molecular identification and susceptibility profile of Sporothrix schenckii sensu lato isolated in Argentina. Mycoses 2018. [Google Scholar] [CrossRef] [PubMed]
- Kohler, L.M.; Soares, B.M.; de Assis Santos, D.; Da Silva Barros, M.E.; Hamdan, J.S. In vitro susceptibility of isolates of Sporothrix schenckii to amphotericin B, itraconazole, and terbinafine: Comparison of yeast and mycelial forms. Can. J. Microbiol. 2006, 52, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Gremiao, I.D.; Menezes, R.C.; Schubach, T.M.; Figueiredo, A.B.; Cavalcanti, M.C.; Pereira, S.A. Feline sporotrichosis: Epidemiological and clinical aspects. Med. Mycol. 2015, 53, 15–21. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strain | Species (Molecular Identification) | Lesion Type | Origin from Mexico | GenBank Accession No. CAL Gen | Variant in Phylogenetic Tree |
|---|---|---|---|---|---|
| 98-164 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948670 | 1 |
| 98-578 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948671 | 1 |
| 99-098 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948672 | 2 |
| 99-689 | Sporothrix schenckii | Fixed cutaneous | Mexico City | MF948673 | 1 |
| 00-45 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948674 | 2 |
| 02-845 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948675 | 4 |
| 02-846 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948676 | 2 |
| 02-847 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948677 | 2 |
| 02-849 | Sporothrix schenckii | Fixed cutaneous | Mexico City | MF948678 | 2 |
| 02-850 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948679 | 1 |
| 02-851 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948680 | 1 |
| 02-852 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948681 | 2 |
| 03-017 | Sporothrix schenckii | Lymphocutaneous | Veracruz | MF948682 | 1 |
| 03-018 | Sporothrix schenckii | Lymphocutaneous | Veracruz | MF948683 | 1 |
| 06-345 | Sporothrix schenckii | Lymphocutaneous | San Luis Potosi | MF948684 | 2 |
| 06-743 | Sporothrix schenckii | Lymphocutaneous | San Luis Potosi | MF948685 | 4 |
| 07-089 | Sporothrix schenckii | Lymphocutaneous | Coahuila | MF948686 | 2 |
| 07-956 | Sporothrix schenckii | Lymphocutaneous | San Luis Potosi | MF948687 | 3 |
| 08-345 | Sporothrix schenckii | Lymphocutaneous | Coahuila | MF948688 | 2 |
| 08-390 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948689 | 1 |
| 08-624 | Sporothrix schenckii | Lymphocutaneous | Nuevo Leon | MF948690 | 3 |
| 09-321 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948691 | 3 |
| 11-131 | Sporothrix schenckii | Lymphocutaneous | Puebla | MF948692 | 1 |
| 11-567 | Sporothrix schenckii | Lymphocutaneous | Nuevo Leon | MF948693 | 1 |
| 12-078 | Sporothrix schenckii | Lymphocutaneous | Oaxaca | MF948694 | 1 |
| 13-670 | Sporothrix globosa | Lymphocutaneous | Nuevo Leon | MF948695 | 5 |
| 14-821 | Sporothrix schenckii | Lymphocutaneous | Puebla | MF948696 | 2 |
| 14-822 | Sporothrix schenckii | Lymphocutaneous | Puebla | MF948697 | 1 |
| 14-823 | Sporothrix schenckii | Lymphocutaneous | Puebla | MF948698 | 1 |
| 14-824 | Sporothrix schenckii | Lymphocutaneous | Oaxaca | MF948699 | 1 |
| 16-09 | Sporothrix schenckii | Lymphocutaneous | Oaxaca | MF948700 | 3 |
| 16-020 | Sporothrix schenckii | Lymphocutaneous | Oaxaca | MF948701 | 4 |
| 16-021 | Sporothrix schenckii | Disseminated | Oaxaca | MF948702 | 3 |
| 16-022 | Sporothrix schenckii | Lymphocutaneous | Oaxaca | MF948703 | 1 |
| 16-230 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948704 | 1 |
| 16-236 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948705 | 1 |
| 16-237 | Sporothrix schenckii | Lymphocutaneous | Mexico City | MF948706 | 1 |
| 16-678 | Sporothrix schenckii | Lymphocutaneous | Jalisco | MF948707 | 1 |
| 16-679 | Sporothrix schenckii | Lymphocutaneous | Jalisco | MF948708 | 1 |
| Antifungal | GM | Range (µg/mL) | MIC50 (µg/mL) | MIC90 (µg/mL) |
|---|---|---|---|---|
| Amphotericin B AMB | 2.554 | 0.5–4 | 4 | 4 |
| Itraconazole ITC | 4 | 0.125–2 | 4 | 8 |
| Voriconazole VRC | 2.946 | 0.5–8 | 2 | 4 |
| Posaconazole PSC | 1.769 | 0.5–8 | 1 | 8 |
| Fluconazole FLC | >64 | 64–>64 | 64 | >64 |
| Terbinafine TRB | 0.374 | 0.06–1 | 0.5 | 1 |
| Caspofungin CSF | 1.959 | 0.5–8 | 2 | 4 |
| Anidulafungin ANF | 2.083 | 0.25–8 | 2 | 4 |
| Micafungin MCF | 2.402 | 0.5–8 | 2 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas, O.C.; Bonifaz, A.; Campos, C.; Treviño-Rangel, R.D.J.; González-Álvarez, R.; González, G.M. Molecular Identification, Antifungal Susceptibility, and Geographic Origin of Clinical Strains of Sporothrix schenckii Complex in Mexico. J. Fungi 2018, 4, 86. https://doi.org/10.3390/jof4030086
Rojas OC, Bonifaz A, Campos C, Treviño-Rangel RDJ, González-Álvarez R, González GM. Molecular Identification, Antifungal Susceptibility, and Geographic Origin of Clinical Strains of Sporothrix schenckii Complex in Mexico. Journal of Fungi. 2018; 4(3):86. https://doi.org/10.3390/jof4030086
Chicago/Turabian StyleRojas, Olga C., Alexandro Bonifaz, Christian Campos, Rogelio De J. Treviño-Rangel, Rafael González-Álvarez, and Gloria M. González. 2018. "Molecular Identification, Antifungal Susceptibility, and Geographic Origin of Clinical Strains of Sporothrix schenckii Complex in Mexico" Journal of Fungi 4, no. 3: 86. https://doi.org/10.3390/jof4030086
APA StyleRojas, O. C., Bonifaz, A., Campos, C., Treviño-Rangel, R. D. J., González-Álvarez, R., & González, G. M. (2018). Molecular Identification, Antifungal Susceptibility, and Geographic Origin of Clinical Strains of Sporothrix schenckii Complex in Mexico. Journal of Fungi, 4(3), 86. https://doi.org/10.3390/jof4030086