Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi



Abstract
:1. Introduction
2. Cuticular Infection Route
3. Alternative Infection Routes for Entomopathogenic Fungi
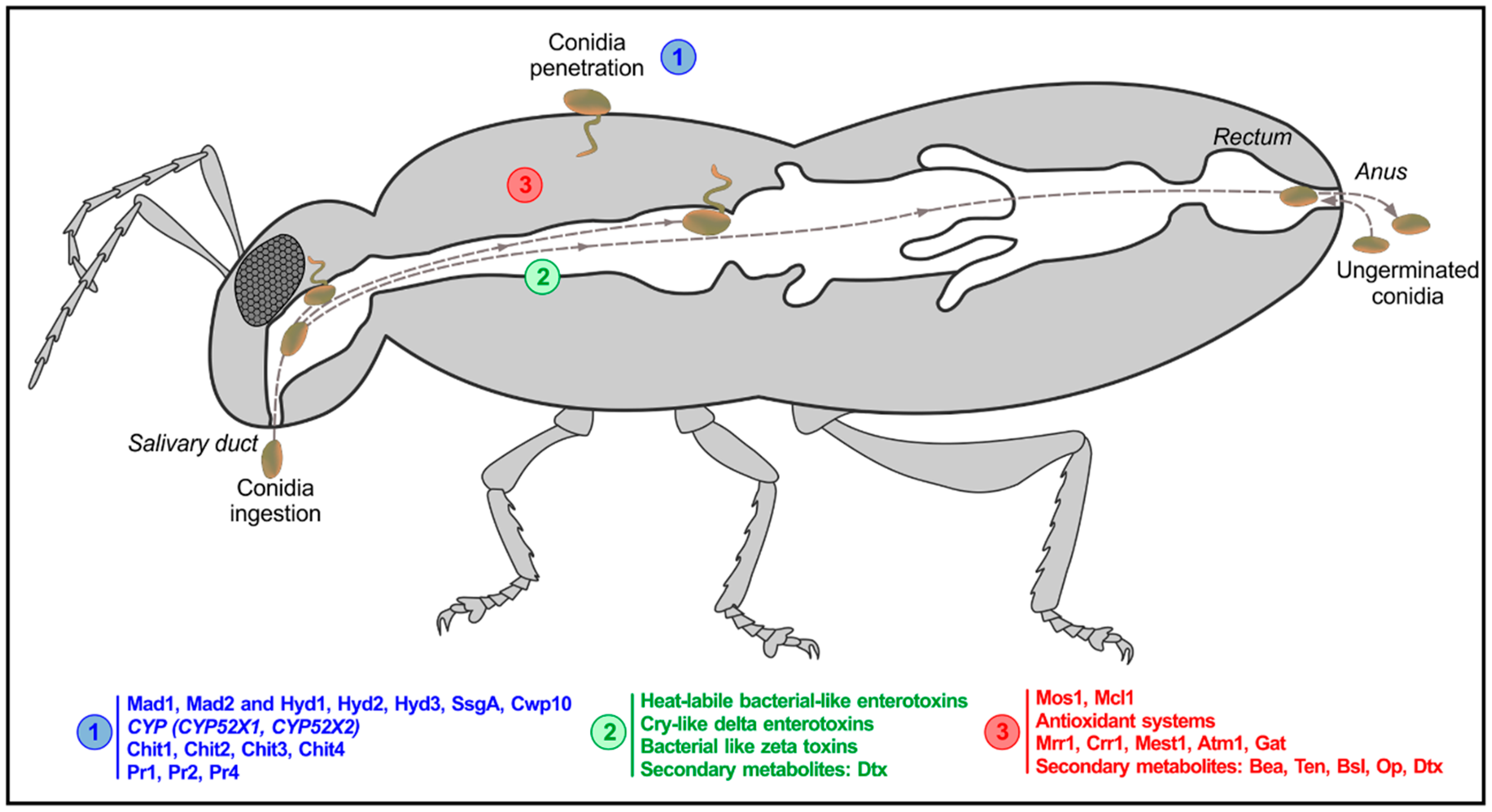
3.1. Oral Infection Route in Terrestrial Insects
3.2. Oral Infection Routes in Aquatic Insects: The Particular Case of Mosquito Larvae
4. What Is Known at the Molecular Level? Candidates and Shared Pathways
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Araújo, J.P.M.; Hughes, D.P. Diversity of entomopathogen fungi: Which groups conquered the insec body? In Advances in Genetics, 1st ed.; Lovett, B., St. Leger, R., Eds.; Elsevier: Cambridge, MA, USA, 2016; Volume 94, pp. 1–39. [Google Scholar]
- Blackwell, M.; Vega, F.E. Introduction: Seven Wonders of the Insect–Fungus World. In Insect-Fungal Associations: Ecology and Evolution, 1st ed.; Blackwell, M., Vega, F.E., Eds.; Oxford University Press: New York, NY, USA, 2005; pp. XIII–XVII. [Google Scholar]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovett, B.; St. Leger, R.J. The Insect Pathogens. Microbiol. Spectr. 2017, 5, 1–19. [Google Scholar]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Schabel, S. Oral infection of Hylobius pales by Metarhizium anisopliae. J. Invertebr. Pathol. 1976, 383, 377–383. [Google Scholar] [CrossRef]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; St. Leger, R.J.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef]
- Gao, Q.; Jin, K.; Ying, S.; Zhang, Y.; Xiao, G.; Shang, Y.; Duan, Z.; Hu, X.; Xie, X.; Zhou, G.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 5, 122. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xiao, G.; Zheng, P.; Shang, Y.; Su, Y.; Zhang, X.; Liu, X.; Zhan, S. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafaluk-Mohr, C.; Wagner, S.; Joop, G. Cryptic changes in immune response and fitness in Tribolium castaneum as a consequence of coevolution with Beauveria bassiana. J. Invertebr. Pathol. 2018, 152, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Boucias, D.; Pedland, J. Attachment of mycopathogens to cuticle. The initial events of mycoses in arthropod hosts. In The Fungal Spore and Disease Initiation in Plant and Animals, 1st ed.; Cole, G., Hoch, H., Eds.; Springer: Boston, MA, USA, 1991; pp. 101–127. [Google Scholar]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic Fungi: New Insights into Host-Pathogen Interactions. In Advances in Genetics, 1st ed.; Lovett, B., St. Leger, R., Eds.; Elsevier: Cambridge, MA, USA, 2016; Volume 94, pp. 307–364. [Google Scholar]
- Pedrini, N.; Crespo, R.; Juárez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. - C Toxicol. Pharmacol. 2007, 146, 124–137. [Google Scholar] [CrossRef]
- Pedrini, N.; Zhang, S.; Juárez, M.; Keyhani, N. Molecular characterization and expression analysis of a suite of cytochrome P450 enzymes implicated in insect hydrocarbon degradation in the entomopathogenic fungus Beauveria bassiana. Microbiology 2010, 2549–2557. [Google Scholar] [CrossRef]
- Zhang, S.; Widemann, E.; Bernard, G.; Lesot, A.; Pinot, F.; Pedrini, N.; Keyhani, N.O. CYP52X1, representing new cytochrome P450 subfamily, displays fatty acid hydroxylase activity and contributes to virulence and growth on insect cuticular substrates in entomopathogenic fungus Beauveria bassiana. J. Biol. Chem. 2012, 287, 13477–13486. [Google Scholar] [CrossRef]
- Lin, L.; Fang, W.; Liao, X.; Wang, F.; Wei, D.; St. Leger, R.J. The MrCYP52 cytochrome P450 monoxygenase gene of Metarhizium robertsii is important for utilizing insect epicuticular hydrocarbons. PLoS ONE 2011, 6, e28984. [Google Scholar] [CrossRef]
- Ferron, P. Fungal control. In Comprehensive Insect Physiology, Biochemistry and Pharmacology, 1st ed.; Kerkut, G., Gilbert, L., Eds.; Academic Press: New York, NY, USA, 1985; pp. 313–346. [Google Scholar]
- Boucias, D.G. Entomopathogenic Fungi: “Perfect” Phyla. In Principles of Insect Pathology, 1st ed.; Boucias, D.G., Pendland, J.C., Eds.; Springer: New York, NY, USA, 1998; pp. 287–319. [Google Scholar]
- Nahar, P.; Ghormade, V.; Deshpande, M.V. The extracellular constitutive production of chitin deacetylase in Metarhizium anisopliae: Possible edge to entomopathogenic fungi in the biological control of insect pests. J. Invertebr. Pathol. 2004, 85, 80–88. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef]
- Lyakhovich, V.V.; Vavilin, V.A.; Zenkov, N.K.; Menshchikova, E.B. Active Defense under Oxidative Stress. The Antioxidant Responsive Element. Biochemistry 2006, 71, 962–974. [Google Scholar] [CrossRef]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: An evolutionary perspective. Trends Plant Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef]
- Singh, D.; Kaur, G. Production, HPLC analysis, and in situ apoptotic activities of Swainsonine. Biotechnol. Program. 2014, 30, 1196–1205. [Google Scholar] [CrossRef]
- Biswas, T.; Joop, G.; Rafaluk-Mohr, C. Cross-resistance: A consequence of bi-partite host-parasite coevolution. Insects 2018, 9, 28. [Google Scholar] [CrossRef]
- Batta, Y.A. Efficacy of two species of entomopathogenic fungi against the stored-grain pest, Sitophilus granarius L. (Curculionidae: Coleoptera), via oral ingestion. Egypt. J. Biol. Pest Control 2018, 1–8. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.A.; Quintela, E.D.; Mascarin, G.M.; Pedrini, N.; Liao, L.M.; Ferri, P.H. Unveiling chemical defense in the rice stalk stink bug against the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2015, 127, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.C.; Hartzer, K.; Toutges, M.; Oppert, B. Evaluation of quantitative PCR reference genes for gene expression studies in Tribolium castaneum after fungal challenge. J. Microbiol. Methods 2010, 80, 219–221. [Google Scholar] [CrossRef]
- Pedrini, N.; Villaverde, M.; Fuse, C.; Dal Bello, G.; Juárez, M. Beauveria bassiana infection alters colony development and defensive secretions of the beetles Tribolium castaneum and Ulomoides dermestoides (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2010, 103, 1094–1099. [Google Scholar] [CrossRef]
- Keilin, D. On a new saccharomycete Monosporella unicuspidata gen.n.nom, n.sp., parasitic in the body cavity of a dipterous larva (Dasyhelea obscura Winnertz). Parasitology 1920, 12, 83–91. [Google Scholar] [CrossRef]
- Veen, K. Oral infection of second-instar nymphs of Schistocerca gregaria by Metarhizium anisopliae. J. Invertebr. Pathol. 1966, 8, 254–256. [Google Scholar] [CrossRef]
- Peirson, H. The life history and control of the pales weevil (Hylobius pales). Harvard For. Bull. 1921, 3, 33. [Google Scholar]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2013, 7, 562–578. [Google Scholar] [CrossRef]
- Choi, M.K.; Le, M.T.; Nguyen, D.T.; Choi, H.; Kim, W.; Kim, J.H.; Chun, J.; Hyeon, J.; Seo, K.; Park, C. Genome-level identification, gene expression, and comparative analysis of porcine ß-defensin genes. BMC Genet. 2012, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Nontini, G.; Mathlein, R. Groenmykos foerorsakad av Metarhizium anisopliae (Metsch.) Sorok. I. Groenmykosen som biologiskt insekt- bekaempningsmedel. Medd. Vaextskyddanst. 1944, 43, 1–58. [Google Scholar]
- Gabriel, B. Fungus infections of insects via the alimentary tract. J. Insect Pathol. 1959, 1, 19–330. [Google Scholar]
- Radha, K.; Nirula, K.K.; Menok, K.P.V. The green muscardine disease of Oryctes rhinoceros L. II. The causal organism. Rev. Appl. Entomol. 1956, 45, 336–337. [Google Scholar]
- Dillon, R.J.; Charnley, A.K. Invasion of the pathogenic fungus Metarhizium anisopliae through the guts of germfree desert locusts, Schistocerca gregaria. Mycopathologia 1986, 96, 59–66. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Inhibition of Metarhizium anisopliae by the gut bacterial flora of the desert locust, Schistocerca gregaria: Evidence for an antifungal toxin. J. Invertebr. Pathol. 1986, 47, 350–360. [Google Scholar] [CrossRef]
- Allee, L.L.; Goettel, M.S.; Gol’berg, A.; Whitney, H.S.; Roberts, D.W. Infection by Beauveria bassiana of Leptinotarsa decemlineata larvae as a consequence of fecal contamination of the integument following per os inoculation. Mycopathologia 1990, 111, 17–24. [Google Scholar] [CrossRef]
- Yanagita, T. Studies on oral infection of larvae of the silkworm, Bombyx mori, with Beauveria bassiana. J. Seric Sci. Japan 1987, 56, 279–284. [Google Scholar]
- Lacey, C.M.; Lacey, L.A.; Roberts, D.R. Route of invasion and histopathology of Metarhizium anisopliae in Culex quinquefasciatus. J. Invertebr. Pathol. 1988, 52, 108–118. [Google Scholar] [CrossRef]
- Goettel, M.S. Viability of Tolypocladium cylindrosporum (Hyphomycetes) conidia following ingestion and excretion by larval Aedes aegypti. J. Invertebr. Pathol. 1988, 51, 275–277. [Google Scholar] [CrossRef]
- Miranpuri, G.S.; Khachatourians, G.G. Infection sites of theentomopathogenic fungus Beauveria bassiana in the larvae of the mosquito Aedes aegypti. Entomol. Exp. Appl. 1991, 59, 19–27. [Google Scholar] [CrossRef]
- Butt, T.M.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.G.; Taylor, J.W.D.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Garrido-Jurado, I.; et al. Metarhizium anisopliae pathogenesis of mosquito larvae: A verdict of accidental death. PLoS ONE 2013, 8, e81686. [Google Scholar] [CrossRef]
- Sweeney, A.W. Infection of mosquito larvae by Culicinomyces sp. through anal papillae. J. Invertebr. Pathol. 1979, 33, 249–251. [Google Scholar] [CrossRef]
- Sweeney, A.W. The mode of infection of the insect pathogenic fungus Culicinomyces in larvae of the mosquito Culex fatigans. Aust. J. Zool. 1975, 23, 1–8. [Google Scholar] [CrossRef]
- Rodrigues, J.; Campos, V.C.; Humber, R.A.; Luz, C. Efficacy of Culicinomyces spp. against Aedes aegypti eggs, larvae and adults. J. Invertebr. Pathol. 2018, 157, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Luz, C.; Humber, R.A. New insights into the in vitro development and virulence of Culicinomyces spp. as fungal pathogens of Aedes aegypti. J. Invertebr. Pathol. 2017, 146, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. The role of epigenetics in host–parasite coevolution: Lessons from the model host insects Galleria mellonella and Tribolium castaneum. Zoology 2016, 119, 273–280. [Google Scholar] [CrossRef]
- Qin, Y.; Ying, S.H.; Chen, Y.; Shen, Z.C.; Feng, M.G. Integration of insecticidal protein Vip3aa1 into Beauveria bassiana enhances fungal virulence to Spodoptera litura larvae by cuticle and per os infection. Appl. Environ. Microbiol. 2010, 76, 4611–4618. [Google Scholar] [CrossRef]
- Farenhorst, M.; Knols, B.G.J. Fungal entomopathogens for the control of adult mosquitoes: A look at the issues. Proc. Neth. Entomol. Soc. Meet. 2007, 18, 51–59. [Google Scholar]


{kind=link}
| Gene Family | Description | Number of Genes | |
|---|---|---|---|
| Beauveria bassiana | Metarhizium robertsii | ||
| Heat labile bacterial-like toxins | Bacterial heat labile enterotoxin IIB, A chain (enzymatic) and IIA A | 13 | 6 |
| Cry-like delta enterotoxins | Bacterial delta endotoxin, N-terminal | 8 | 0 |
| Zeta toxins, bacterial-like | Bacterial toxin | 3 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. J. Fungi 2019, 5, 33. https://doi.org/10.3390/jof5020033
Mannino MC, Huarte-Bonnet C, Davyt-Colo B, Pedrini N. Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. Journal of Fungi. 2019; 5(2):33. https://doi.org/10.3390/jof5020033
Chicago/Turabian StyleMannino, M. Constanza, Carla Huarte-Bonnet, Belén Davyt-Colo, and Nicolás Pedrini. 2019. "Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi" Journal of Fungi 5, no. 2: 33. https://doi.org/10.3390/jof5020033
APA StyleMannino, M. C., Huarte-Bonnet, C., Davyt-Colo, B., & Pedrini, N. (2019). Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. Journal of Fungi, 5(2), 33. https://doi.org/10.3390/jof5020033