Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains Used in This Study
2.2. Antifungal Susceptibility Testing
2.3. Definition of MIC Phenomena
2.4. Galleria mellonella In Vivo Treatment Studies
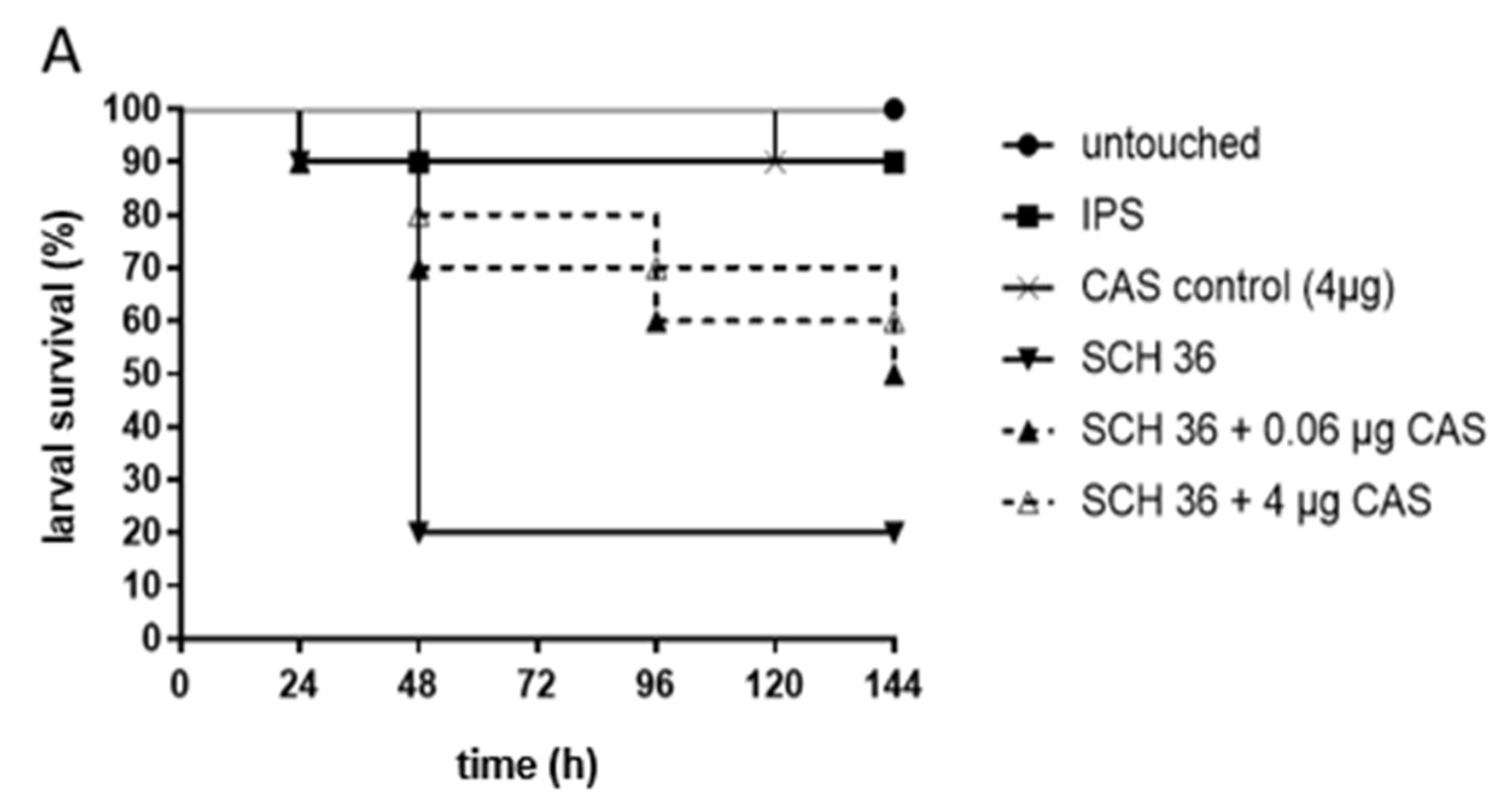
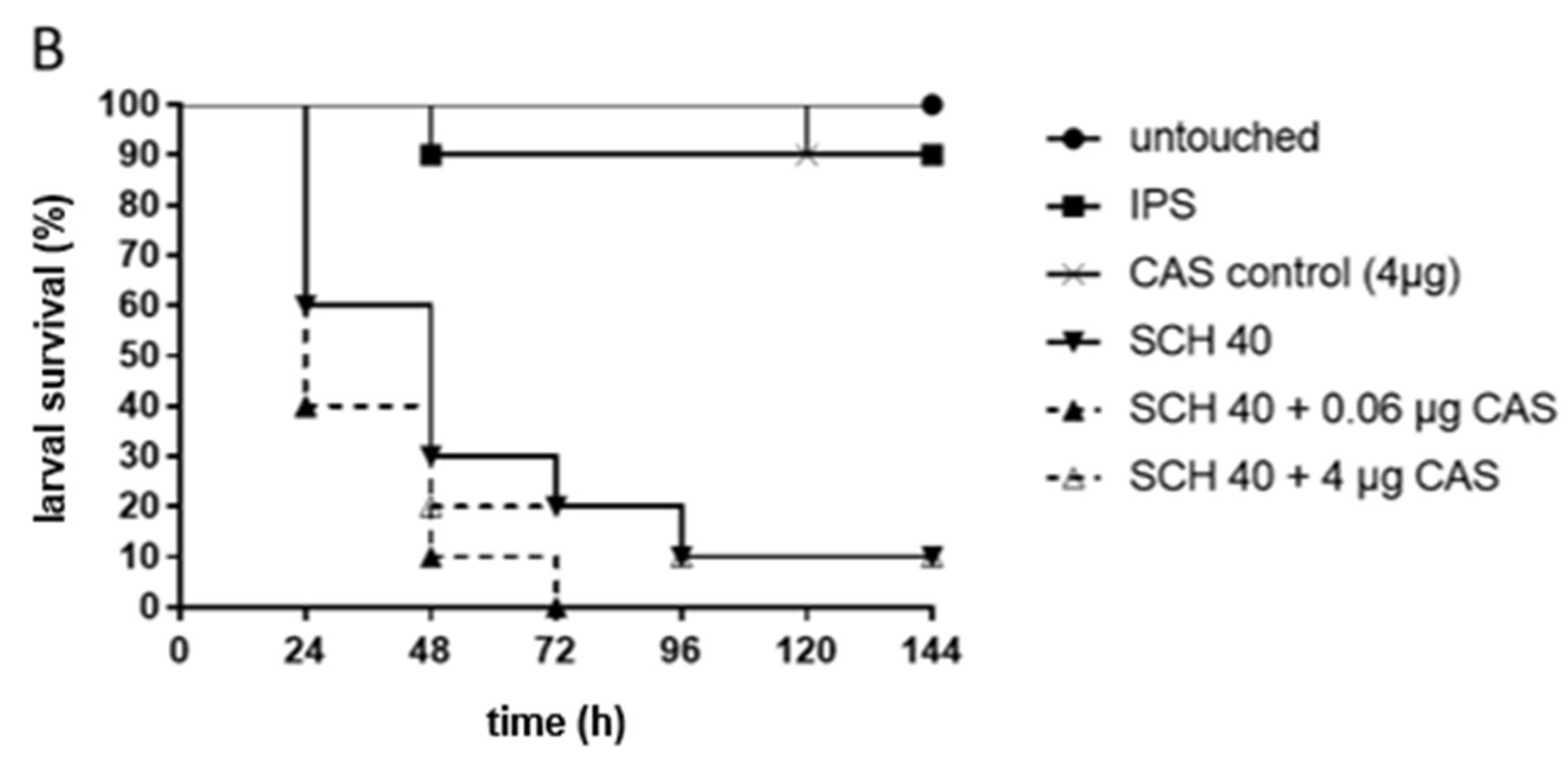
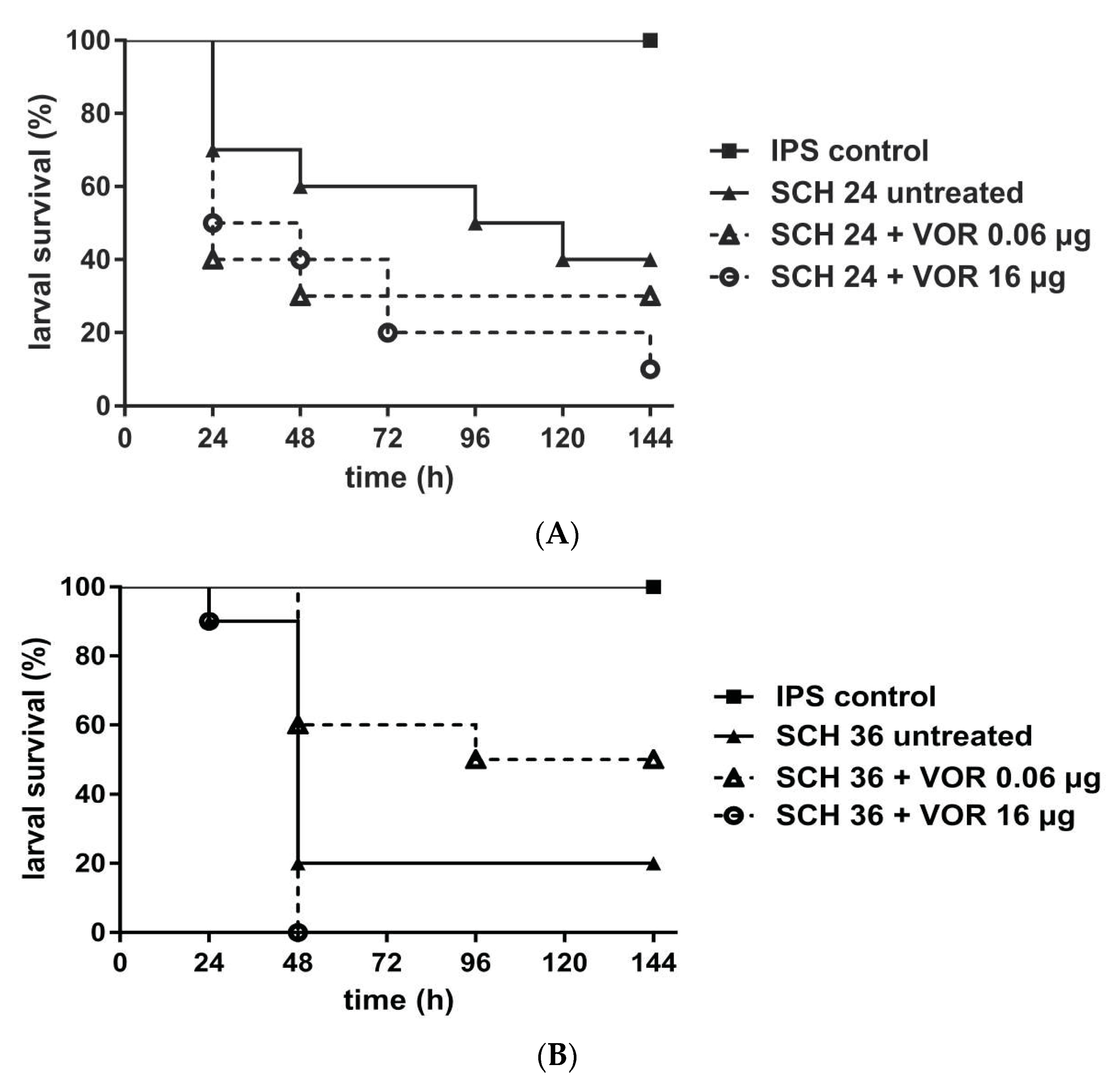
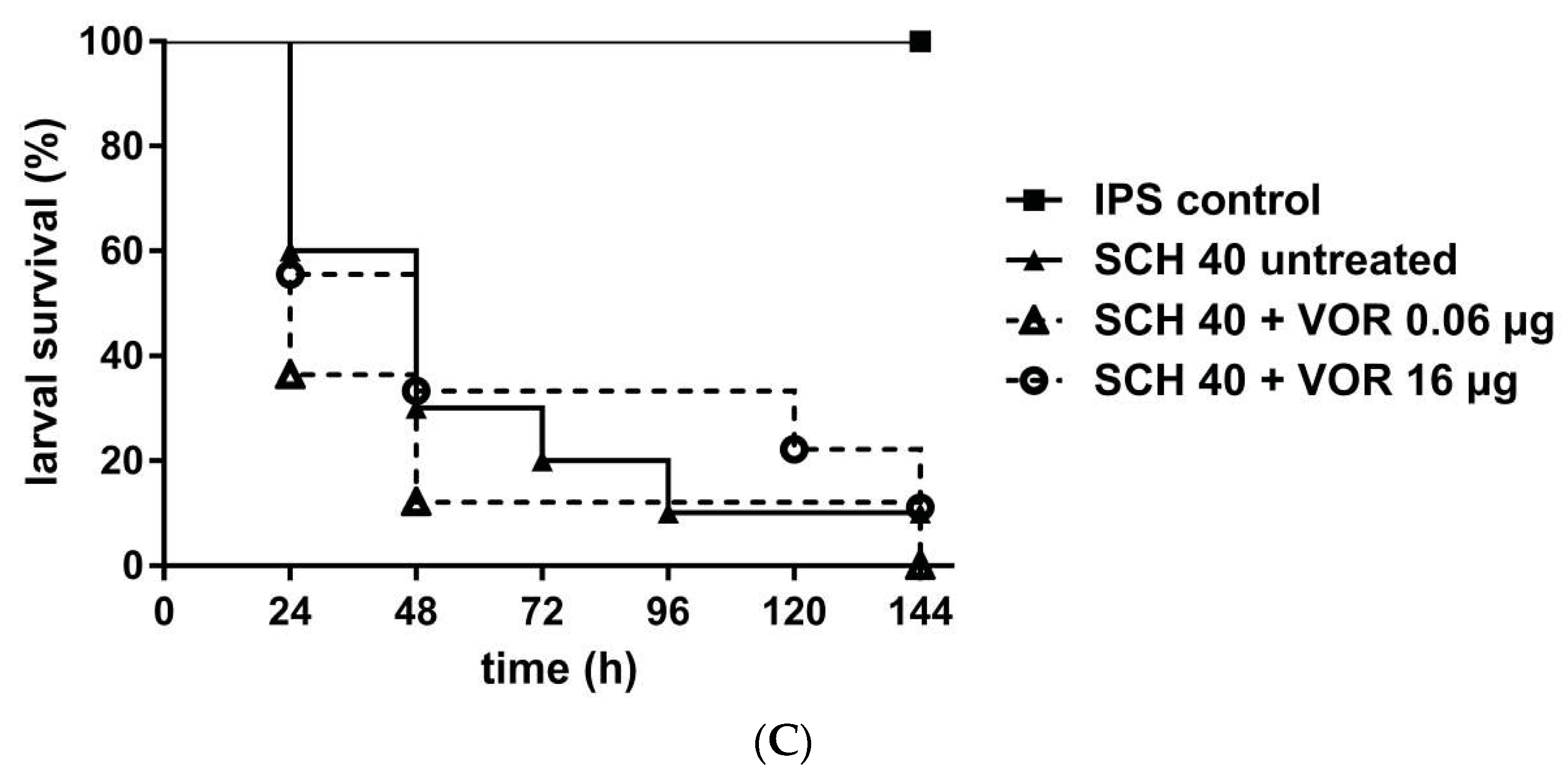
3. Results
4. Discussion
4.1. MIC-Phenomena
4.2. Galleria mellonella as a Model to Study MIC-Phenomena
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical practice guideline for the management of candidiasis: 2016 update by The Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Tudela, J.L.; Arendrup, M.C.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.W.; Donnelly, J.P.; Dromer, F.; et al. EUCAST definitive document EDEF 7.1: Method for the determination of broth dilution mics of antifungal agents for fermentative yeasts: Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST)∗. Clin. Microbiol. Infect. 2008, 14, 398–405. [Google Scholar]
- NCCLS. Reference method for broth dilution antifungal susceptibility testing of yeasts. In NCCLS document M27-A2, Bitte Folgendermaßen Zitieren, 2nd ed.; NCCLS: Wayne, PA, USA, 2002; ISBN 1-56238-469-4. [Google Scholar]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W.W. Breakpoints for antifungal agents: An update from eucast focussing on echinocandins against Candida spp. And triazoles against Aspergillus spp. Drug Resist. Updates 2013, 16, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A.J.; Cornely, O.A.; DonneNy, J.P.; Akova, M.; Arendrup, M.C.; Arikan-Akdagli, S.; Bassetti, M.; Bille, J.; Calandra, T.; Castagnola, E.; et al. ESCMID information in this manuscript was presented in part at ECCMID 2011. European Society for Clinical Microbiology and Infectious Diseases guideline for the diagnosis and management of Candida diseases 2012: Developing european guidelines in clinical microbiology and infectious diseases. Clin. Microbiol. Infect. 2012, 18, 1–8. [Google Scholar] [PubMed]
- Khlif, M.; Bogreau, H.; Michel-Nguyen, A.; Ayadi, A.; Ranque, S. Trailing or paradoxical growth of Candida albicans when exposed to caspofungin is not associated with microsatellite genotypes. Antimicrob. Agents Chemother. 2010, 54, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Schmalreck, A.F.; Kottmann, I.; Reiser, A.; Ruffer, U.; Scharr, E.; Vanca, E. An evaluation of seven methods of testing in vitro susceptibility of clinical yeast isolates to fluconazole. Mycoses 1995, 38, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Revankar, S.G.; Kirkpatrick, W.R.; McAtee, R.K.; Fothergill, A.W.; Redding, S.W.; Rinaldi, M.G.; Patterson, T.F. Interpretation of trailing endpoints in antifungal susceptibility testing by the national committee for clinical laboratory standards method. J. Clin. Microbiol. 1998, 36, 153–156. [Google Scholar] [PubMed]
- Lopez-Ribot, J.L.; McAtee, R.K.; Perea, S.; Kirkpatrick, W.R.; Rinaldi, M.G.; Patterson, T.F. Multiple resistant phenotypes of Candida albicans coexist during episodes of oropharyngeal candidiasis in human immunodeficiency virus-infected patients. Antimicrob. Agents Chemother. 1999, 43, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Rueda, C.; Puig-Asensio, M.; Guinea, J.; Almirante, B.; Cuenca-Estrella, M.; Zaragoza, O.; Padilla, B.; Muñoz, P.; Guinea, J.; Paño Pardo, J.R.; et al. Evaluation of the possible influence of trailing and paradoxical effects on the clinical outcome of patients with candidemia. Clin. Microbiol. Infect. 2017, 23, e41–e49. [Google Scholar] [CrossRef]
- Rueda, C.; Cuenca-Estrella, M.; Zaragoza, O. Paradoxical growth of Candida albicans in the presence of caspofungin is associated with multiple cell wall rearrangements and decreased virulence. Antimicrob. Agents Chemother. 2014, 58, 1071–1083. [Google Scholar] [CrossRef]
- Lee, M.-K.; Williams, L.E.; Warnock, D.W.; Arthington-Skaggs, B.A. Drug resistance genes and trailing growth in Candida albicans isolates. J. Antimicrob. Chemother. 2004, 53, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Gómez, A.; Sánchez-Carrillo, C.; Bouza, E.; Muñoz, P.; Escribano, P.; Guinea, J. Isavuconazole is highly active in vitro against Candida species isolates but shows trailing effect. Clin. Microbiol. Infect. 2018, 24, e1341–e1344. [Google Scholar] [CrossRef] [PubMed]
- Vanstraelen, K.; Lagrou, K.; Maertens, J.; Wauters, J.; Willems, L.; Spriet, I. The eagle-like effect of echinocandins: What’s in a name? Expert Rev. Anti-Infect. Ther. 2013, 11, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Mariné, M.; Pastor, F.J.; Sahand, I.H.; Pontón, J.; Quindós, G.; Guarro, J. Paradoxical growth of Candida dubliniensis does not preclude in vivo response to echinocandin therapy. Antimicrob. Agents Chemother. 2009, 53, 5297–5299. [Google Scholar] [CrossRef] [PubMed]
- Clemons, K.V.; Espiritu, M.; Parmar, R.; Stevens, D.A. Assessment of the paradoxical effect of caspofungin in therapy of candidiasis. Antimicrob. Agents Chemother. 2006, 50, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.A.; Ichinomiya, M.; Koshi, Y.; Horiuchi, H. Escape of candida from caspofungin inhibition at concentrations above the MIC (paradoxical effect) accomplished by increased cell wall chitin; evidence for β-1,6-glucan synthesis inhibition by caspofungin. Antimicrob. Agents Chemother. 2006, 50, 3160–3161. [Google Scholar] [CrossRef]
- Bizerra, F.C.; Melo, A.S.A.; Katchburian, E.; Freymüller, E.; Straus, A.H.; Takahashi, H.K.; Colombo, A.L. Changes in cell wall synthesis and ultrastructure during paradoxical growth effect of caspofungin on four different candida species. Antimicrob. Agents Chemother. 2011, 55, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P.; Kontoyiannis, D.P.; Prince, R.A.; Lewis, R.E. Attenuation of the activity of caspofungin at high concentrations against Candida albicans: Possible role of cell wall integrity and calcineurin pathways. Antimicrob. Agents Chemother. 2005, 49, 5146–5148. [Google Scholar] [CrossRef]
- Cowen, L.E.; Lindquist, S. Hsp90 potentiates the rapid evolution of new traits: Drug resistance in diverse fungi. Science 2005, 309, 2185–2189. [Google Scholar] [CrossRef]
- Munro, C.A.; Selvaggini, S.; De Bruijn, I.; Walker, L.; Lenardon, M.D.; Gerssen, B.; Milne, S.; Brown, A.J.P.; Gow, N.A.R. The pkc, hog and Ca2+ signalling pathways co-ordinately regulate chitin synthesis in Candida albicans. Mol. Microbiol. 2007, 63, 1399–1413. [Google Scholar] [CrossRef]
- Li, X.; Cai, Q.; Mei, H.; Zhou, X.; Shen, Y.; Li, D.; Liu, W. The RPD3/HDA1 family of histone deacetylases regulates azole resistance in Candida albicans. J. Antimicrob. Chemother. 2015, 70, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N. Attenuation of echinocandin activity at elevated concentrations: A review of the paradoxical effect. Curr. Opin. Infect. Dis. 2008, 20, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, S.; López-Ribot, J.L.; Najvar, L.K.; McCarthy, D.I.; Bocanegra, R.; Graybill, J.R. Caspofungin resistance in Candida albicans: Correlating clinical outcome with laboratory susceptibility testing of three isogenic isolates serially obtained from a patient with progressive Candida esophagitis. Antimicrob. Agents Chemother. 2004, 48, 1382–1383. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Rustad, T.R.; Rex, J.H.; White, T.C. The trailing end point phenotype in antifungal susceptibility testing is pH dependent. Antimicrob. Agents Chemother. 1999, 43, 1383–1386. [Google Scholar] [CrossRef] [PubMed]
- Bayegan, S.; Majoros, L.; Kardos, G.; Kemény-Beke, A.; Miszti, C.; Kovacs, R.; Gesztelyi, R. In vivo studies with a Candida tropicalis isolate exhibiting paradoxical growth in vitro in the presence of high concentration of caspofungin. J. Microbiol. 2010, 48, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Kavanagh, K. Caspofungin primes the immune response of the larvae of Galleria mellonella and induces a non-specific antimicrobial response. J. Med Microbiol. 2011, 60, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Sheehan, G. The use of Galleria mellonella larvae to identify novel antimicrobial agents against fungal species of medical interest. J. Fungi (Basel) 2018, 4, 113. [Google Scholar] [CrossRef]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Yang, H.-F.; Pan, A.-J.; Hu, L.-F.; Liu, Y.-Y.; Cheng, J.; Ye, Y.; Li, J.-B. Galleria mellonella as an in vivo model for assessing the efficacy of antimicrobial agents against Enterobacter cloacae infection. J. Microbiol. Immunol. Infect. 2017, 50, 55–61. [Google Scholar] [CrossRef]
- Jacobsen, I.D. Galleria mellonella as a model host to study virulence of candida. Virulence 2014, 5, 237–239. [Google Scholar] [CrossRef]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med Mycol. 2018, 57, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yang, X.; Yu, C.; Gong, Y.; Yuan, L.; Hao, L.; Sun, S. Linezolid in combination with azoles induced synergistic effects against Candida albicans and protected Galleria mellonella against experimental candidiasis. Front. Microbiol. 2019, 9, 3142. [Google Scholar] [CrossRef] [PubMed]
- Morio, F.; Lombardi, L.; Binder, U.; Loge, C.; Robert, E.; Graessle, D.; Bodin, M.; Lass-Flörl, C.; Butler, G.; Le Pape, P. Precise genome editing using a CRISPR-Cas9 method highlights the role of COERG11 amino acid substitutions in azole resistance in Candida orthopsilosis. J. Antimicrob. Chemother. 2019, 74, 2230–2238. [Google Scholar] [CrossRef]
- Astvad, K.M.T.; Meletiadis, J.; Whalley, S.; Arendrup, M.C. Fluconazole pharmacokinetics in Galleria mellonella larvae and performance evaluation of a bioassay compared to liquid chromatography-tandem mass spectrometry for hemolymph specimens. Antimicrob. Agents Chemother. 2017, 61, e00895-17. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and Amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Reeves, E.P. Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jukic, E.; Blatzer, M.; Binder, U.; Mayr, L.; Lass-Flörl, C.; Lackner, M. Impact of morphological sectors on antifungal susceptibility testing and virulence studies. Antimicrob. Agents Chemother. 2017, 61, e00755-17. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Fallon, J.P. Galleria mellonella larvae as models for studying fungal virulence. Fungal Biol. Rev. 2010, 24, 79–83. [Google Scholar] [CrossRef]
- Brennan, M.; Thomas, D.Y.; Whiteway, M.; Kavanagh, K. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med Microbiol. 2002, 34, 153–157. [Google Scholar] [CrossRef]
- Cotter, G.; Doyle, S.; Kavanagh, K. Development of an insect model for the in vivo pathogenicity testing of yeasts. FEMS Immunol. Med Microbiol. 2000, 27, 163–169. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martínez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol. 2013, 51, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.B.; O’Brien, E.; El Khoury, J.B.; Mylonakis, E. Methods for using Galleria mellonella as a model host to study fungal pathogenesis. Virulence 2010, 1, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.; Veli, N.; Coote, P.J. Evaluation of galleria mellonella larvae for measuring the efficacy and pharmacokinetics of antibiotic therapies against Pseudomonas aeruginosa infection. Int. J. Antimicrob. Agents 2014, 43, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Moreno, R.; El Khoury, J.B.; Idnurm, A.; Heitman, J.; Calderwood, S.B.; Ausubel, F.M.; Diener, A. Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 2005, 73, 3842–3850. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; de Lucas, M.P.; Mesa-Arango, A.C.; Fusco-Almeida, A.M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M.J.; Zaragoza, O. Antifungal efficacy during candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, D.; Patterson, T.F.; Rinaldi, M.G.; Revankar, S.G. Trailing end-point phenotype of Candida spp. In antifungal susceptibility testing to fluconazole is eliminated by altering incubation temperature. J. Med Microbiol. 2007, 56, 1003–1004. [Google Scholar] [CrossRef] [PubMed]
- Braga-Silva, L.; Mesquita, D.G.A.; Ribeiro, M.D.; Carvalho, S.M.F.; Fracalanzza, S.E.L.; Santos, A. Trailing end-point phenotype antibiotic-sensitive strains of Candida albicans produce different amounts of aspartyl peptidases. Braz. J. Med. Biol. Res. 2009, 42, 765–770. [Google Scholar] [CrossRef]
- Tsakris, A.; Johnson, A.P.; George, R.C.; Mehtar, S.; Vatopoulos, A.C. Distribution and transferability of plasmids encoding trimethoprim resistance in urinary pathogens from greece. J. Med. Microbiol. 1991, 34, 153–157. [Google Scholar] [CrossRef]
- Stevens, D.A.; Espiritu, M.; Parmar, R. Paradoxical effect of caspofungin: Reduced activity against Candida albicans at high drug concentrations. Antimicrob. Agents Chemother. 2004, 48, 3407–3411. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating echinocandin MIC and kinetic inhibition of FKS1 mutant glucan synthases for Candida albicans: Implications for interpretive breakpoints. Antimicrob. Agents Chemother. 2009, 53, 112–122. [Google Scholar] [CrossRef]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. A clinical isolate of candida albicans with mutations in ERG11 (encoding sterol 14alpha-demethylase) and ERG5 (encoding C22 desaturase) is cross resistant to azoles and amphotericin b. Antimicrob. Agents Chemother. 2010, 54, 3578–3583. [Google Scholar] [CrossRef]
- Lackner, M.; Tscherner, M.; Schaller, M.; Kuchler, K.; Mair, C.; Sartori, B.; Istel, F.; Arendrup, M.C.; Lass-Flörl, C. Positions and numbers of fks mutations in Candida albicans selectively influence in vitro and in vivo susceptibilities to echinocandin treatment. Antimicrob. Agents Chemother. 2014, 58, 3626–3635. [Google Scholar] [CrossRef]
- Arthington-Skaggs, B.A.; Lee-Yang, W.; Ciblak, M.A.; Frade, J.P.; Brandt, M.E.; Hajjeh, R.A.; Harrison, L.H.; Sofair, A.N.; Warnock, W.D. Comparison of visual and spectrophotometric methods of broth microdilution MIC end point determination and evaluation of a sterol quantitation method for in vitro susceptibility testing of fluconazole and itraconazole against trailing and nontrailing Candida isolates. Antimicrob. Agents Chemother. 2002, 46, 2477–2481. [Google Scholar] [PubMed]
- Lee, M.-K.; Kim, H.-R.; Kang, J.-O.; Kim, M.-N.; Kim, E.-C.; Kim, J.-S.; Kim, J.-J.; Park, Y.-J.; Song, W.; Shin, J.-H.; et al. Susceptibility and trailing growth of Candida albicans to fluconazole: Results of a korean multicentre study. Mycoses 2007, 50, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Arthington-Skaggs, B.A.; Warnock, D.W.; Morrison, C.J. Quantitation of Candida albicans ergosterol content improves the correlation between in vitro antifungal susceptibility test results and in vivo outcome after fluconazole treatment in a murine model of invasive candidiasis. Antimicrob. Agents Chemother. 2000, 44, 2081–2085. [Google Scholar] [CrossRef] [PubMed]
- Odabasi, Z.; Paetznick, V.L.; Rodriguez, J.R.; Chen, E.; Rex, J.H.; Leitz, G.J.; Ostrosky-Zeichner, L. Lack of correlation of 24- vs. 48-h itraconazole minimum inhibitory concentrations with microbiological and survival outcomes in a Guinea pig model of disseminated candidiasis. Mycoses 2010, 53, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Escribano, P.; Sánchez-Carrillo, C.; Bouza, E.; Guinea, J. Scope and frequency of fluconazole trailing assessed using eucast in invasive Candida spp. Isolates. Med. Mycol. 2016, 54, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Forastiero, A.; Bernal-Martínez, L.; Mellado, E.; Cendejas, E.; Gomez-Lopez, A. In vivo efficacy of voriconazole and posaconazole therapy in a novel invertebrate model of Aspergillus fumigatus infection. Int. J. Antimicrob. Agents 2015, 46, 511–517. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Species | Antifungal Drug | MIC 24 h (µg/mL) | MIC Phenomena (µg/mL) |
|---|---|---|---|---|
| SCH 24 | C. albicans | VOR | 0.06 | trailing a > 8 |
| FLU | 0.5 | trailing a > 32 | ||
| POS | 0.03 | trailing a > 8 | ||
| ITR | 0.125 | trailing a > 16 | ||
| CAS | 0.12 | |||
| ANI | 0.03 | |||
| AMB | 0.25 | |||
| SCH 36 | C. albicans | VOR | 0.01 | trailing a > 8 |
| FLU | 1 | trailing a > 32 | ||
| POS | 0.01 | trailing a > 8 | ||
| ITR | 0.125 | trailing a > 16 | ||
| CAS | 0.06 | paradoxical growth b at 4 and 8 | ||
| ANI | 0.03 | |||
| AMB | 0.25 | |||
| SCH 40 | C. albicans | VOR | 0.01 | trailing a > 8 |
| FLU | 1 | trailing a > 32 | ||
| POS | 0.01 | trailing a > 8 | ||
| ITR | 0.125 | trailing a > 16 | ||
| CAS | 0.03 | paradoxical growth b at 4 and 8 | ||
| ANI | 0.03 | paradoxical growth b at 4 | ||
| AMB | 0.25 | |||
| VOR | 0.0156 | |||
| FLU | 0.5 | |||
| POS | 0.0078 | trailing a > 8 | ||
| SCH 39 | C. albicans | ITR | 0.125 | |
| CAS | 2 | |||
| ANI | 0.06 | |||
| AMB | 0.25 | |||
| VOR | 0.0156 | trailing a > 8 | ||
| FLU | 0.5 | trailing a > 32 | ||
| POS | 0.015 | trailing a > 4 | ||
| CA 53 | C. albicans | ITR | 0.03 | trailing a > 16 |
| CAS | 2 | |||
| ANI | 1 | |||
| AMB | 0.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, U.; Aigner, M.; Risslegger, B.; Hörtnagl, C.; Lass-Flörl, C.; Lackner, M. Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance. J. Fungi 2019, 5, 83. https://doi.org/10.3390/jof5030083
Binder U, Aigner M, Risslegger B, Hörtnagl C, Lass-Flörl C, Lackner M. Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance. Journal of Fungi. 2019; 5(3):83. https://doi.org/10.3390/jof5030083
Chicago/Turabian StyleBinder, Ulrike, Maria Aigner, Brigitte Risslegger, Caroline Hörtnagl, Cornelia Lass-Flörl, and Michaela Lackner. 2019. "Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance" Journal of Fungi 5, no. 3: 83. https://doi.org/10.3390/jof5030083
APA StyleBinder, U., Aigner, M., Risslegger, B., Hörtnagl, C., Lass-Flörl, C., & Lackner, M. (2019). Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance. Journal of Fungi, 5(3), 83. https://doi.org/10.3390/jof5030083