Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Termite Collection and Maintenance
2.2. Fungal Culture and Suspension Preparation
2.3. Screening of Fungal Isolates for Time–Mortality Responses
2.4. Cocoa Seeds and Seedlings
2.5. Inoculation Methods
2.6. Assessment of Colonization
2.7. Morphological Identification of the Inoculated Endophytes
2.8. Molecular Identification of Naturally Occurring Endophytes Isolated from Cocoa Seedlings
2.8.1. DNA Extraction and PCR Amplification
2.8.2. Sequencing and Analysis
2.9. Data Analysis
3. Results
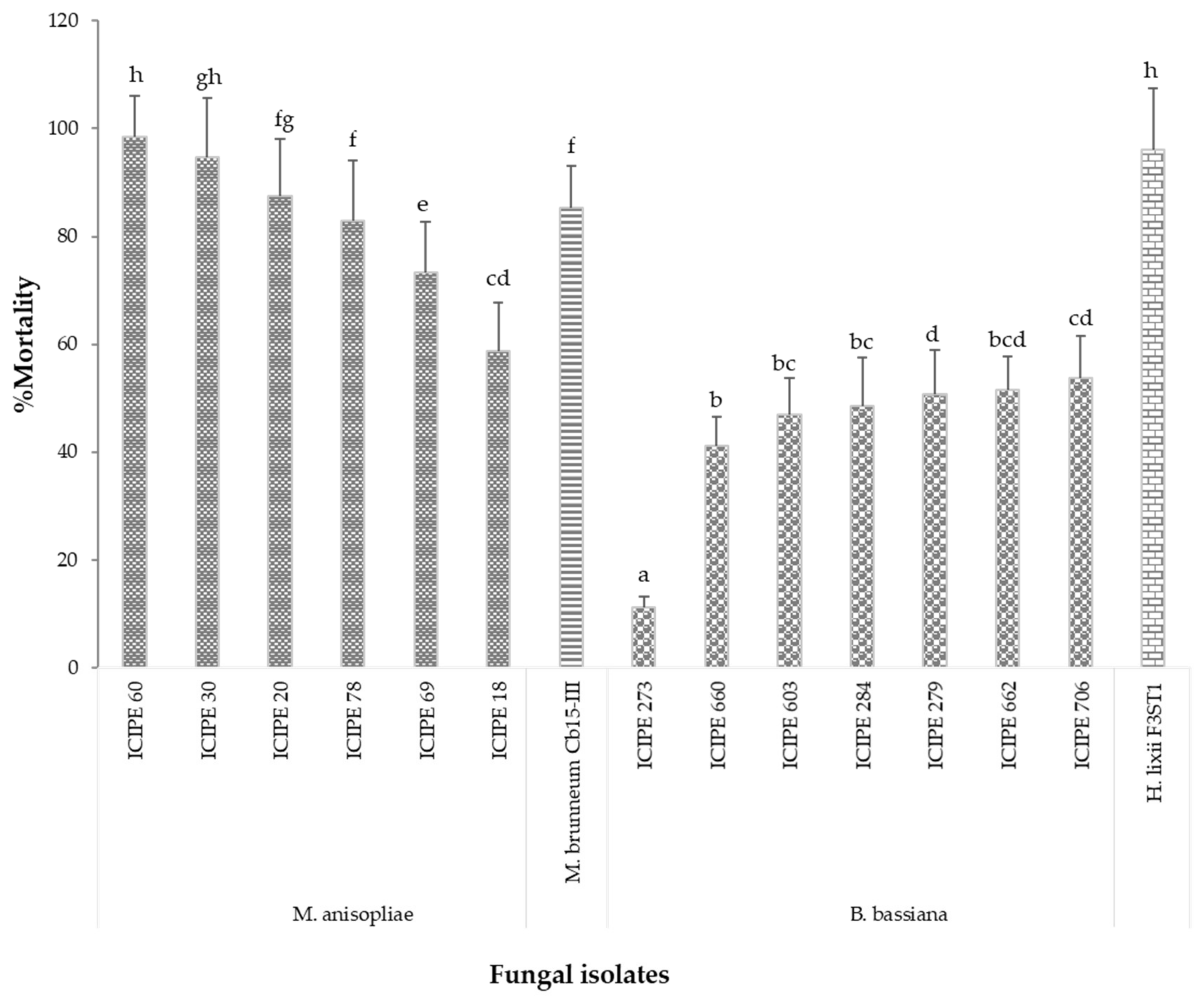
3.1. Pathogenicity of EPF Isolates against Termites
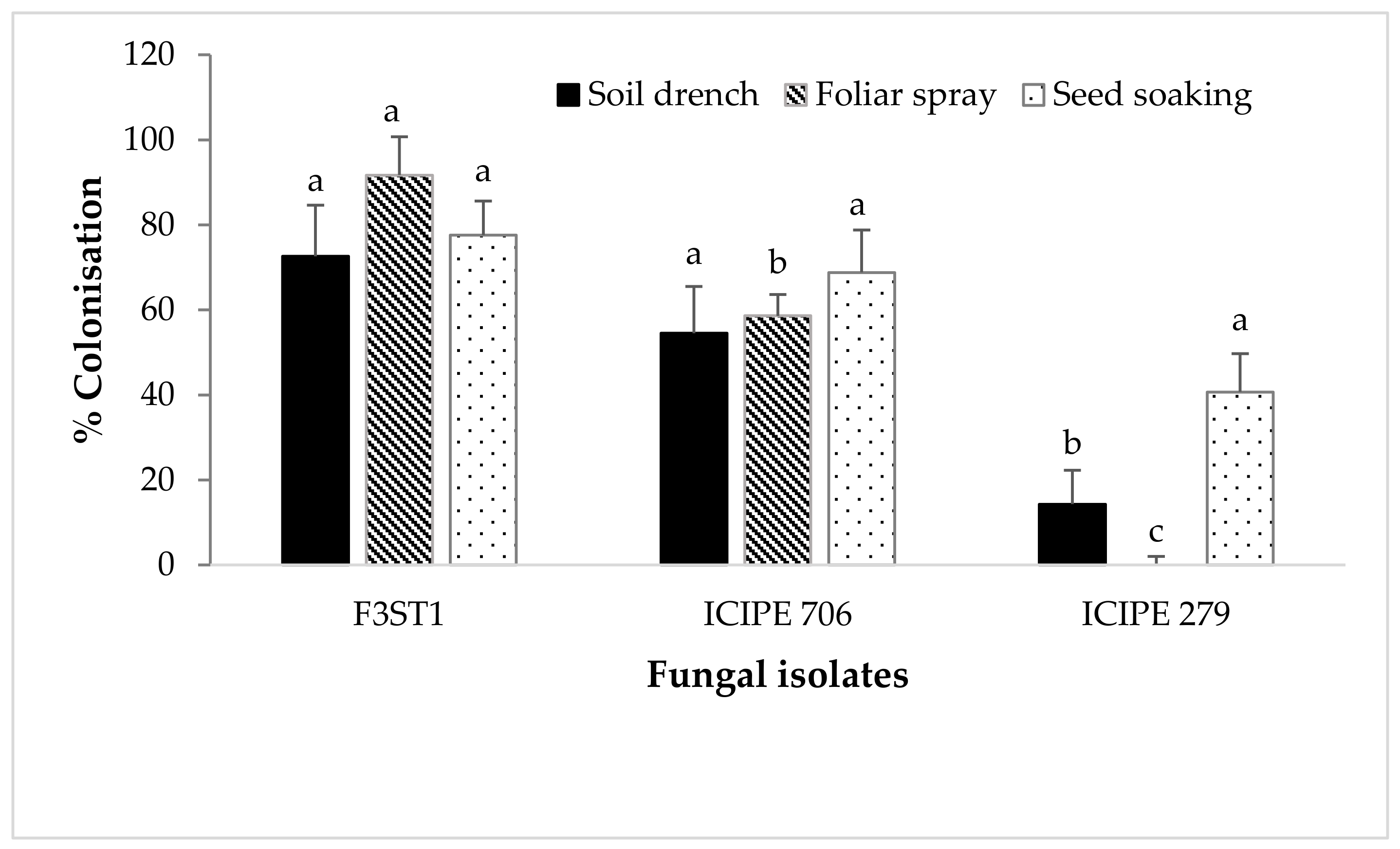
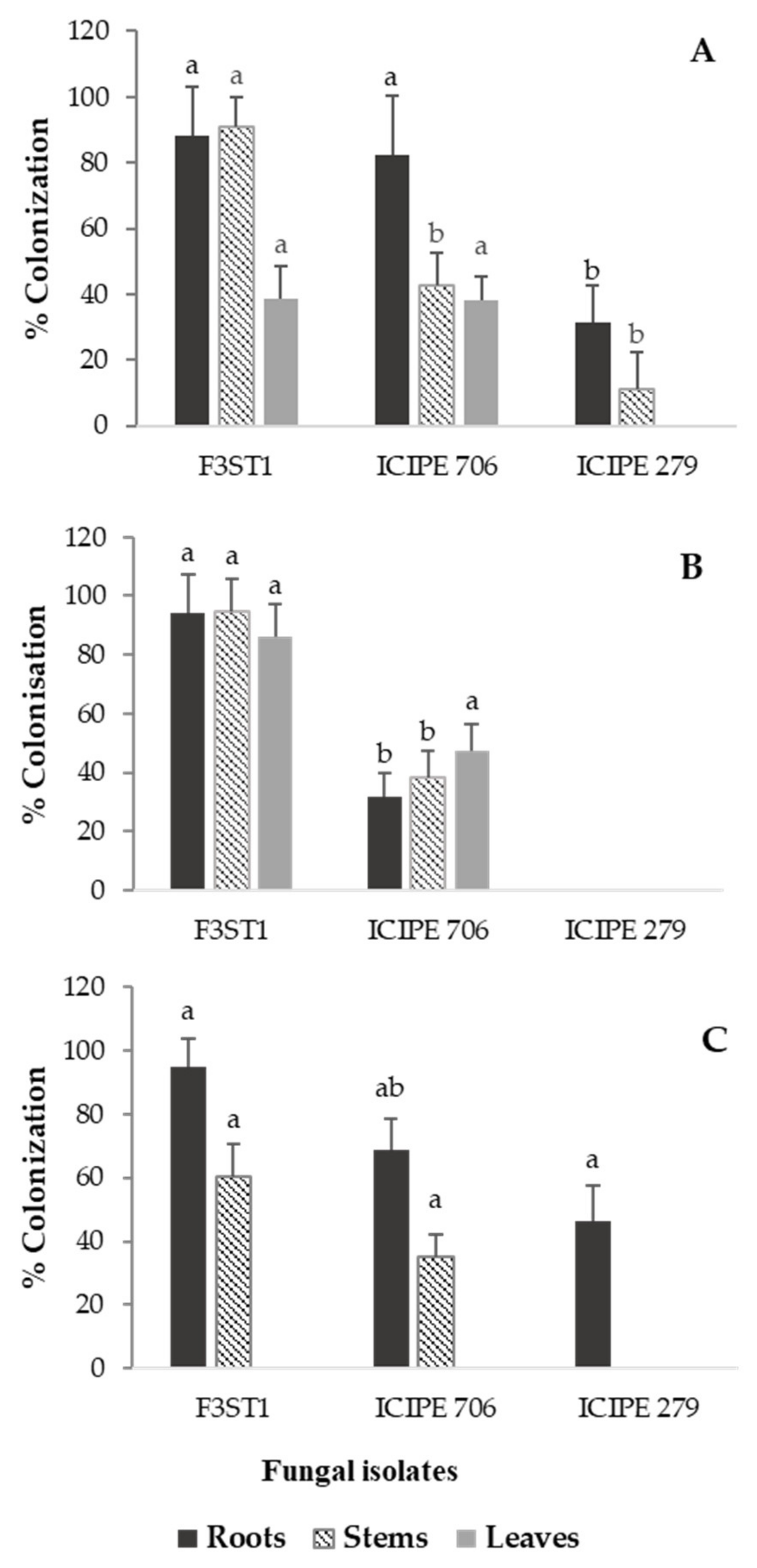
3.2. Endophytic Colonization of Cocoa Seedlings by Artificially Inoculated Fungal Isolates
3.3. Naturally Occurring Fungal Endophytes Isolated from the Cocoa Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tra Bi, C.S.; Soro, S.; Yéboué-Nguessan, L.; Tano, Y.; Konaté, S. Termites impact on different age of cocoa (Theobroma cacao L.) plantations with different fertilizer treatments in semi-deciduous forest zone (Oume, Ivory Coast). HJAFSR 2015, 4, 021–027. [Google Scholar]
- Ambele, F.C.; Bisseleua, D.H.B.; Babalola, O.O.; Ekesi, S. Soil-dwelling insect pests of tree crops in Sub-Saharan Africa, problems and management strategies—A review. J. Appl. Entomol. 2018, 142, 539–552. [Google Scholar] [CrossRef]
- Ambele, F.C.; Hervé, B.D.; Ekesi, S.; Akutse, K.S.; Djuideu, C.T.; Meupia, M.J.; Babalola, O.O. Consequences of shade management on the taxonomic patterns and functional diversity of termites (Blattodea: Termitidae) in cocoa agroforestry systems. Evol. Ecol. 2018, 8, 11582–11595. [Google Scholar]
- Djuideu, T.C.L.; Bisseleua, D.H.B.; Kekeunou, S.; Meupia, M.J.; Difouo, F.G.; Ambele, C.F. Plant community composition and functional characteristics define invasion and infestation of termites in cocoa agroforestry systems. Agrofor. Syst. 2020, 94, 185–201. [Google Scholar] [CrossRef]
- Su, N.Y.; Scheffrahn, R.H. A review of subterranean termite control practices and prospects for integrated pest management programmes. Integr. Pest Manag. Rev. 1998, 3, 1–13. [Google Scholar] [CrossRef]
- Grace, J.K. Biological control strategies for suppression of termites. J. Agric. Entomol. 1997, 14, 281–289. [Google Scholar]
- Kramm, K.R.; West, D.F. Termite pathogens: Effects of ingested Metarhizium, Beauveria, and Gliocladium conidia on worker termites (Reticulitermes sp.). J. Invertebr. Pathol. 1982, 40, 7–11. [Google Scholar] [CrossRef]
- Rath, A.C. The use of entomopathogenic fungi for control of termites. Biocontrol Sci. Technol. 2000, 10, 563–581. [Google Scholar] [CrossRef]
- Smythe, R.V.; Coppel, H.C. The susceptibility of Reticulitermes flavipes (Kollar) and other termite species to an experimental preparation of Bacillus thuringiensis Berliner. J. Invertebr. Pathol. 1965, 7, 423–426. [Google Scholar] [CrossRef]
- Yendol, W.G.; Paschke, J.D. Pathology of an Entomophthora infection in the eastern subterranean termite Reticulitermes flavipes (Kollar). J. Invertebr. Pathol. 1965, 7, 414–422. [Google Scholar] [CrossRef]
- Culliney, T.W.; Grace, J.K. Prospects for the biological control of subterranean termites (Isoptera: Rhinotermitidae), with special reference to Coptotermes formosanus. Bull. Entomol. Res. 2000, 90, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Chouvenc, T.; Su, N.Y.; Grace, J.K. Fifty years of attempted biological control of termites-Analysis of a failure. Biol. Control 2011, 59, 69–82. [Google Scholar] [CrossRef]
- Yanagawa, A.; Yokohari, F.; Shimizu, S. Defense mechanism of the termite, Coptotermes formosanus Shiraki, to entomopathogenic fungi. J. Invertebr. Pathol. 2008, 97, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Mburu, D.M.; Ochola, L.; Maniania, N.K.; Njagi, P.G.N.; Gitonga, L.M.; Ndung’u, M.W.; Hassanali, A. Relationship between virulence and repellency of entomopathogenic isolates of Metarhizium anisopliae and Beauveria bassiana to the termite Macrotermes michaelseni. J. Insect Physiol. 2009, 55, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E. Insect pathology and fungal endophytes. J. Invertebr. Pathol. 2008, 98, 277–279. [Google Scholar] [CrossRef]
- Grace, J.K. Approaches to biological control of termites. Sociobiology 2003, 41, 115–121. [Google Scholar]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.Z.; Fuxa, J.R.; Richter, A.; Ring, D. Interactions of Metarhizium anisoplae and tree-based mulches in repellence and mycoses against Coptotermes formosanus (Isoptera: Rhinotermitidae). Environ. Entomol. 2014, 37, 755–763. [Google Scholar] [CrossRef]
- Maniania, N.K.; Ekesi, S.; Songa, J.M. Managing termites in maize with the entomopathogenic fungus Metarhizium anisopliae. Int. J. Trop. Insect Sci. 2002, 22, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Lewis, L.C. Ecology and Evolution of Fungal Endophytes, and Their Roles Against Insects. Insect-Fungal Associations: Ecology and Evolution; Oxford University Press: New York, NY, USA, 2005; pp. 74–96. [Google Scholar]
- Bing, L.A.; Lewis, L.C. Suppression of Ostrinia nubilalis (Hübner) (Lepidoptera: Pyralidae) by endophytic Beauveria bassiana (Balsamo) Vuillemin. Environ Entomol. 1991, 20, 1207–1211. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Coyne, D.; Kyamanywa, S. Endophytic Beauveria bassiana in banana (Musa spp.) reduces banana weevil (Cosmopolites sordidus) fitness and damage. Crop Prot. 2008, 27, 1437–1441. [Google Scholar] [CrossRef]
- Leckie, B.M. Effects of Beauveria bassiana Mycelia and Metabolites Incorporated into Synthetic Diet and Fed to Larval Helicoverpa zea; and Detection of Endophytic Beauveria bassiana in Tomato Plants Using PCR and ITS Primers. Master’s Thesis, Department of Entomology, The University of Tennessee, Knoxville, TN, USA, 2002. [Google Scholar]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, F.; Vega, F.E. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedlings (Theobroma cacao). Mycologia 2005, 97, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant–endophyte–herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Pell, J.K. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Wemheuer, F.; Berkelmann, D.; Wemheuer, B.; Daniel, R.; Vidal, S.; Bisseleua, D.H.B. Agroforestry Management Systems Drive the Composition, Diversity, and Function of Fungal and Bacterial Endophyte Communities in Theobroma cacao Leaves. Microorganisms 2020, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- Goettel, M.S.; Inglis, G.D. Fungi: Hyphomycetes. In Manual of Techniques in Insect Pathology; Elsevier: Amsterdam, The Netherlands, 1997; pp. 213–249. [Google Scholar]
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Laboratory techniques used for entomopathogenic fungi: Hypocreales. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Springer: Berlin, Germany, 2012; pp. 189–253. [Google Scholar]
- Ambele, C.F.; Bisseleua, H.D.; Akutse, K.S.; Babalola, O.O.; Humbert, P.; Patel, A.; Ekesi, S. Testing a co-formulation of CO2-releasing material with an entomopathogenic fungus for the management of subterranean termite pests. Mycol. Prog. 2019, 18, 1201–1211. [Google Scholar] [CrossRef]
- Sun, J.; Fuxa, J.R.; Henderson, G. Effects of virulence, sporulation, and temperature on Metarhizium anisopliae and Beauveria bassiana laboratory transmission in Coptotermes formosanus. J. Invertebr. Pathol. 2003, 84, 38–46. [Google Scholar] [CrossRef]
- Humber, R.A. Fungi: Identification. In Manual of Techniques in Insect Pathology; Lacey, L., Ed.; Academic Press: Washington, DC, USA, 1997; pp. 153–185. [Google Scholar]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Akutse, K.S.; Van den Berg, J.; Maniania, N.K.; Ekesi, S.; Fiaboe, K.K.M. Morphological and molecular characterization of Vicia faba and Phaseolus vulgaris seed-born fungal endophytes. Res. J. Seed Sci. 2017, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Burgess, L.W.; Liddell, C.M.; Summerell, B.A. Laboratory Manual for Fusarium Research: Incorporating a Key and Descriptions of Common Species Found in Australasia; FAO: Rome, Italy, 1988. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Coloe, S.; Baird, R.; Pedersen, J. Application of PCR to the identification of dermatophyte fungi. J. Med. Microbiol. 2000, 49, 493–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landeweert, R.; Leeflang, P.; Kuyper, T.W.; Hoffland, E.; Rosling, A.; Wernars, K.; Smit, E. Molecular identification of ectomycorrhizal mycelium in soil horizons. Appl. Environ. Microbiol. 2003, 69, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javadi, M.A.; Ghanbary, M.A.T.; Tazick, Z. Isolation and molecular identification of soil inhabitant Penicillia. Ann. Biol. Res. 2012, 3, 5758–5761. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989. [Google Scholar] [CrossRef]
- Fisher, P.J.; Petrini, O. Location of fungal endophytes in tissues of Suaeda fruticosa: A preliminary study. Trans. Br. Mycol. Soc. 1987, 89, 246–249. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. Available online: https://www.R-project.org/ (accessed on 23 June 2019).
- Zoberi, M.H.; Grace, J.K. Fungi associated with the subterranean termite Reticulitermes flavipes in Ontario. Mycologia 1990, 82, 289–294. [Google Scholar] [CrossRef]
- Dong, C.; Zhang, J.; Chen, W.; Huang, H.; Hu, Y. Characterization of a newly discovered China variety of Metarhizium anisopliae (M. anisopliae var. dcjhyium) for virulence to termites, isoenzyme, and phylogenic analysis. Microbiol. Res. 2007, 162, 53–61. [Google Scholar] [CrossRef]
- Hoe, P.; Bong, C.F.J.; Kadir, J.; Amartalingam, R. Evaluation of Metarhizium anisopliae var. anisopliae (Deuteromycotina: Hyphomycete) isolates and their effects on subterranean termite Coptotermes curvignathus (Isoptera: Rhinotermitidae). Am. J. Agric. Biol. Sci. 2009, 4, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Powell, J.E. Isolation and evaluation of Beauveria bassiana for control of Coptotermes formosanus and Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology 2003, 41, 369–382. [Google Scholar]
- Cherry, A.J.; Abalo, P.; Hell, K. A laboratory assessment of the potential of different strains of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to control Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in stored cowpea. J. Stored Prod. Res. 2005, 41, 295–309. [Google Scholar] [CrossRef]
- Singha, D.; Singha, B.; Dutta, B.K. Virulence of isolates of Metarhizium anisopliae (Metschnikoff) Sorokin and Beauveria bassiana (Ballsamo) Vuillemin on tea termite Microcerotermes beesoni Snyder (Isoptera: Termitidae) of Barak Valley of Assam. Indian J. Entomol. 2006, 68, 66–70. [Google Scholar]
- Singha, D.; Singha, B.; Dutta, B.K. Potential of Metarhizium anisopliae and Beauveria bassiana in the control of tea termite Microtermes obesi Holmgren in vitro and under field conditions. J. Pest Sci. 2011, 84, 69–75. [Google Scholar] [CrossRef]
- Sileshi, A.; Sori, W.; Dawd, M. Laboratory evaluation of entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana against Termite, Macrotermes (Isoptera: Termitidae). Asian J. Plant Sci. 2013, 12, 1–10. [Google Scholar]
- Denier, D.; Bulmer, M.S. Variation in subterranean termite susceptibility to fatal infections by local Metarhizium soil isolates. Insectes Soc. 2015, 62, 219–226. [Google Scholar] [CrossRef]
- Kuswanto, E.; Ahmad, I.; Dungani, R. Threat of subterranean termite attack in the Asian countries and their control: A Review. Asian J. Appl. Sci. 2015, 8, 227–239. [Google Scholar]
- Milner, R.J.; Staples, J.A.; Lenz, M. Options for termite management using the insect pathogenic fungus Metarhizium anisopliae. In Proceedings of the 27th Annual Meeting of the International Research Group on Wood Preservation, Guadaloupe, France, 19–24 May 1996. [Google Scholar]
- Staples, J.A.; Milner, R.J. A laboratory evaluation of the repellency of Metarhizium anisopliae conidia to Coptotermes lacteus (Isoptera: Rhinotermitidae). Sociobiology 2000, 36, 133–148. [Google Scholar]
- Myles, T.G. Alarm, aggregation, and defense by Reticulitermes flavipes in response to a naturally occurring isolate of Metarhizium anisopliae. Sociobiology 2002, 40, 243–256. [Google Scholar]
- Gathage, J.W.; Lagat, Z.O.; Fiaboe, K.K.M.; Akutse, K.S.; Ekesi, S.; Maniania, N.K. Prospects of fungal endophytes in the control of Liriomyza leafminer flies in common bean Phaseolus vulgaris under field conditions. Biocontrol 2016, 61, 741–753. [Google Scholar] [CrossRef]
- Mutune, B.; Ekesi, S.; Niassy, S.; Matiru, V.; Bii, C.; Maniania, N.K. Fungal endophytes as promising tools for the management of bean stem maggot Ophiomyia phaseoli on beans Phaseolus vulgaris. J. Pest Sci. 2016, 89, 993–1001. [Google Scholar] [CrossRef]
- Gómez-Vidal, S.; Lopez-Llorca, L.V.; Jansson, H.B.; Salinas, J. Endophytic colonization of date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. Micron 2006, 37, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Posada, F.; Vega, F.E. Inoculation and colonization of coffee seedlings (Coffea arabica L.) with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycoscience 2006, 47, 284–289. [Google Scholar] [CrossRef]
- Brownbridge, M.; Reay, S.D.; Nelson, T.L.; Glare, T.R. Persistence of Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte following inoculation of radiata pine seed and seedlings. Biol. Control 2012, 61, 194–200. [Google Scholar] [CrossRef]
- Akello, J.; Sikora, R. Systemic acropedal influence of endophyte seed treatment on Acyrthosiphon pisum and Aphis fabae offspring development and reproductive fitness. Biol. Control 2012, 61, 215–221. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; López-Díaz, C.; Landa, B.B. The hidden habit of the entomopathogenic fungus Beauveria bassiana: First demonstration of vertical plant transmission. PLoS ONE 2014, 9, e89278. [Google Scholar] [CrossRef] [Green Version]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol. Control 2010, 55, 34–41. [Google Scholar] [CrossRef]
- Ownley, B.H.; Griffin, M.R.; Klingeman, W.E.; Gwinn, K.D.; Moulton, J.K.; Pereira, R.M. Beauveria bassiana: Endophytic colonization and plant disease control. J. Invertebr. Pathol. 2008, 98, 267–270. [Google Scholar] [CrossRef]
- Jaber, L.R. Grapevine leaf tissue colonization by the fungal entomopathogen Beauveria bassiana s. l. and its effect against downy mildew. Biocontrol 2015, 60, 103–112. [Google Scholar] [CrossRef]
- Akutse, K.S.; Maniania, N.K.; Fiaboe, K.K.M.; Van den Berg, J.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Muvea, A.M.; Meyhöfer, R.; Maniania, N.K.; Poehling, H.M.; Ekesi, S.; Subramanian, S. Behavioral responses of Thrips tabaci Lindeman to endophyte-inoculated onion plants. J. Pest Sci. 2015, 88, 555–562. [Google Scholar] [CrossRef]
- Akutse, K.S.; Fiaboe, K.K.; Van den Berg, J.; Ekesi, S.; Maniania, N.K. Effects of endophyte colonization of Vicia faba (Fabaceae) plants on the life–history of leafminer parasitoids Phaedrotoma scabriventris (Hymenoptera: Braconidae) and Diglyphus isaea (Hymenoptera: Eulophidae). PLoS ONE 2014, 9, e109965. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, M.; Gómez-Jiménez, M.I.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control 2016, 95, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elena, G.J.; Beatriz, P.J.; Alejandro, P.; Lecuona, R.E. Metarhizium anisopliae (Metschnikoff) Sorokin promotes growth and has endophytic activity in tomato plants. Adv. Biol. Res. 2011, 5, 22–27. [Google Scholar]
- Batta, Y.A. Efficacy of endophytic and applied Metarhizium anisopliae (Metch.) Sorokin (Ascomycota: Hypocreales) against larvae of Plutella xylostella L. (Yponomeutidae: Lepidoptera) infesting Brassica napus plants. Crop Prot. 2013, 44, 128–134. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant tissue localization of the endophytic insect pathogenic fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Clavicipitaceae) is also an endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.R.; Martin Nieto, L.; Doohan, F.M.; Hodkinson, T.R. Profundae diversitas: The uncharted genetic diversity in a newly studied group of fungal root endophytes. Mycology 2015, 6, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Akello, J.; Dubois, T.; Gold, C.S.; Coyne, D.; Nakavuma, J.; Paparu, P. Beauveria bassiana (Balsamo) Vuillemin as an endophyte in tissue culture banana (Musa spp.). J. Invertebr. Pathol. 2007, 96, 34–42. [Google Scholar] [CrossRef]
- Tefera, T.; Vidal, S. Effect of inoculation method and plant growth medium on endophytic colonization of sorghum by the entomopathogenic fungus Beauveria bassiana. BioControl 2009, 54, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P.; Mahapatra, B.S. Endophytic colonization of white jute (Corchorus capsularis) plants by different Beauveria bassiana strains for managing stem weevil (Apion corchori). Phytoparasitica 2013, 41, 17–21. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Munoz-Ledesma, F.J.; Santiago-Alvarez, C. Systemic protection of Papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an endophytic strain of Beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Muvea, A.M.; Meyhöfer, R.; Subramanian, S.; Poehling, H.M.; Ekesi, S.; Maniania, N.K. Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS ONE 2014, 9, e108242. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Isolates | Source | Locality/Country | Year of Isolation | % Germination |
|---|---|---|---|---|---|
| Metarhizium anisopliae | ICIPE 20 | Soil | Migori (Kenya) | 1989 | 98.25 ± 0.47 bcd |
| ICIPE 30 | Busseola fusca | Kendubay (Kenya) | 1989 | 98.06 ± 0.47 cde | |
| ICIPE 60 | Soil | Kakelo-Seme (Kenya) | 1990 | 99.10 ± 0.29 abc | |
| ICIPE 18 | Soil | Mbita (Kenya) | 1989 | 99.77 ± 0.01 a | |
| ICIPE 69 | Soil | Matete (DRC) | 1990 | 99.34 ± 0.28 ab | |
| ICIPE 78 | Temnoschoita quadripustulata | Ungoe (Kenya) | 1990 | 99.09 ± 0.03 abc | |
| Metarhizium brunneum | Cb15-III | - | Germany | - | 95.25 ± 0.48 f |
| Beauveria bassiana | |||||
| ICIPE 273 | Soil | Mbita (Kenya) | 2006 | 96.96 ± 0.27 e | |
| ICIPE 279 | Coleopteran larva | Kericho (Kenya) | 2005 | 98.25 ± 0.85 bcd | |
| ICIPE 284 | Coleopteran larva | Mauritus | 2005 | 99.22 ± 0.38 ab | |
| ICIPE 603 | Hymenoptera | Taita (Kenya) | 2007 | 97.26 ± 0.48 de | |
| ICIPE 706 | Monocots | Kenya | 2008 | 99.51 ± 0.01 a | |
| ICIPE 660 | Soil | Chemokock (Kenya) | 2008 | 98.70 ± 0.06 abc | |
| ICIPE 662 | Soil | Mariakani (Kenya) | 2008 | 99.33 ± 0.14 ab | |
| Hypocrea lixii | F3ST1 | Maize | Kenya | 2009 | 95.25 ± 0.45 f |
| Fungal Species | Isolates | LT50 (Days) (95% FL) | % Mycosis of Termite Cadavers |
|---|---|---|---|
| Beauveria bassiana | ICIPE 279 | 2.23 (2.19–2.27) | 66.25 ± 5.91 a |
| ICIPE 603 | 3.09 (3.04–3.14) | 47.50 ± 5.20 e | |
| ICIPE 284 | 3.00 (2.96–3.04) | 61.25 ± 9.44 ab | |
| ICIPE 706 | 2.85 (2.82–2.88) | 61.25 ± 4.27 ab | |
| ICIPE 662 | 2.85 (2.82–2.88) | 70.00 ± 4.56 a | |
| ICIPE 660 | 3.00 (2.96–3.04) | 62.50 ± 3.23 ab | |
| ICIPE 273 | 4.41 (4.37–4.45) | 46.25 ± 5.54 bcd | |
| Metarhizium anisopliae | ICIPE 20 | 2.23 (2.19–2.27) | 48.75 ± 9.66 de |
| ICIPE 60 | 1.51 (1.49–1.53) | 52.50 ± 7.77 abcd | |
| ICIPE 30 | 1.72 (1.69–1.75) | 41.25 ± 3.15 cde | |
| ICIPE 18 | 2.84 (2.80–2.88) | 53.75 ± 2.39 abcd | |
| ICIPE 78 | 2.38 (2.35–2.41) | 47.50 ± 4.33 bcd | |
| ICIPE 69 | 2.41 (2.38–2.44) | 67.50 ± 6.61 abc | |
| Metarhizium brunneum | Cb15-III | 1.50 (1.44–1.56) | 57.50 ± 10.9 bcd |
| Hypocrea lixii | F3ST1 | 1.80 (1.51–2.09) | 62.54 ±7.11 ab |
| Sample Code | Identified Fungal Samples from GenBank | Query Coverage % | E-value | Identities (%) | GenBank ID | Sample Accession Numbers |
|---|---|---|---|---|---|---|
| CU | Trichoderma asperellum isolate NBAIR TATP small subunit ribosomal RNA gene, partial sequence; internal transcribed spacer 1, 5.8S ribosomal RNA gene, and internal transcribed spacer 2, complete sequence; and large subunit ribosomal RNA gene, partial sequence | 100 | 0 | 99.82 | MN727373.1 | MT767125 |
| CL | Fusarium solani isolate 7184.01 internal transcribed spacer 1, partial sequence; 5.8S ribosomal RNA gene and internal transcribed spacer 2, complete sequence; and large subunit ribosomal RNA gene, partial sequence | 99 | 0 | 100 | MN729431.1 | MT767126 |
| CR1 | Fusarium redolens strain T1ST190421511 small subunit ribosomal RNA gene, partial sequence; internal transcribed spacer 1, 5.8S ribosomal RNA gene, and internal transcribed spacer 2, complete sequence; and large subunit ribosomal RNA gene, partial sequence | 100 | 0 | 99.64 | MN486568.1 | MT767127 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambele, C.F.; Ekesi, S.; Bisseleua, H.D.B.; Babalola, O.O.; Khamis, F.M.; Djuideu, C.T.L.; Akutse, K.S. Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings. J. Fungi 2020, 6, 126. https://doi.org/10.3390/jof6030126
Ambele CF, Ekesi S, Bisseleua HDB, Babalola OO, Khamis FM, Djuideu CTL, Akutse KS. Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings. Journal of Fungi. 2020; 6(3):126. https://doi.org/10.3390/jof6030126
Chicago/Turabian StyleAmbele, Chaba F., Sunday Ekesi, Hervé D. B. Bisseleua, Olubukola O. Babalola, Fathiya M. Khamis, Christian T. L. Djuideu, and Komivi S. Akutse. 2020. "Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings" Journal of Fungi 6, no. 3: 126. https://doi.org/10.3390/jof6030126
APA StyleAmbele, C. F., Ekesi, S., Bisseleua, H. D. B., Babalola, O. O., Khamis, F. M., Djuideu, C. T. L., & Akutse, K. S. (2020). Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings. Journal of Fungi, 6(3), 126. https://doi.org/10.3390/jof6030126