Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fungal Isolates
2.3. Generation of Fungal Preparations
2.4. In Vitro Culture of Microglia Cells (CHME-3) and Fungal Cocultivation
2.5. Total RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Immunofluorescence Assay
2.8. Heatmap and Network Analysis for Differentially Expressed Genes (DEGs)
2.9. Statistical Analysis
3. Results
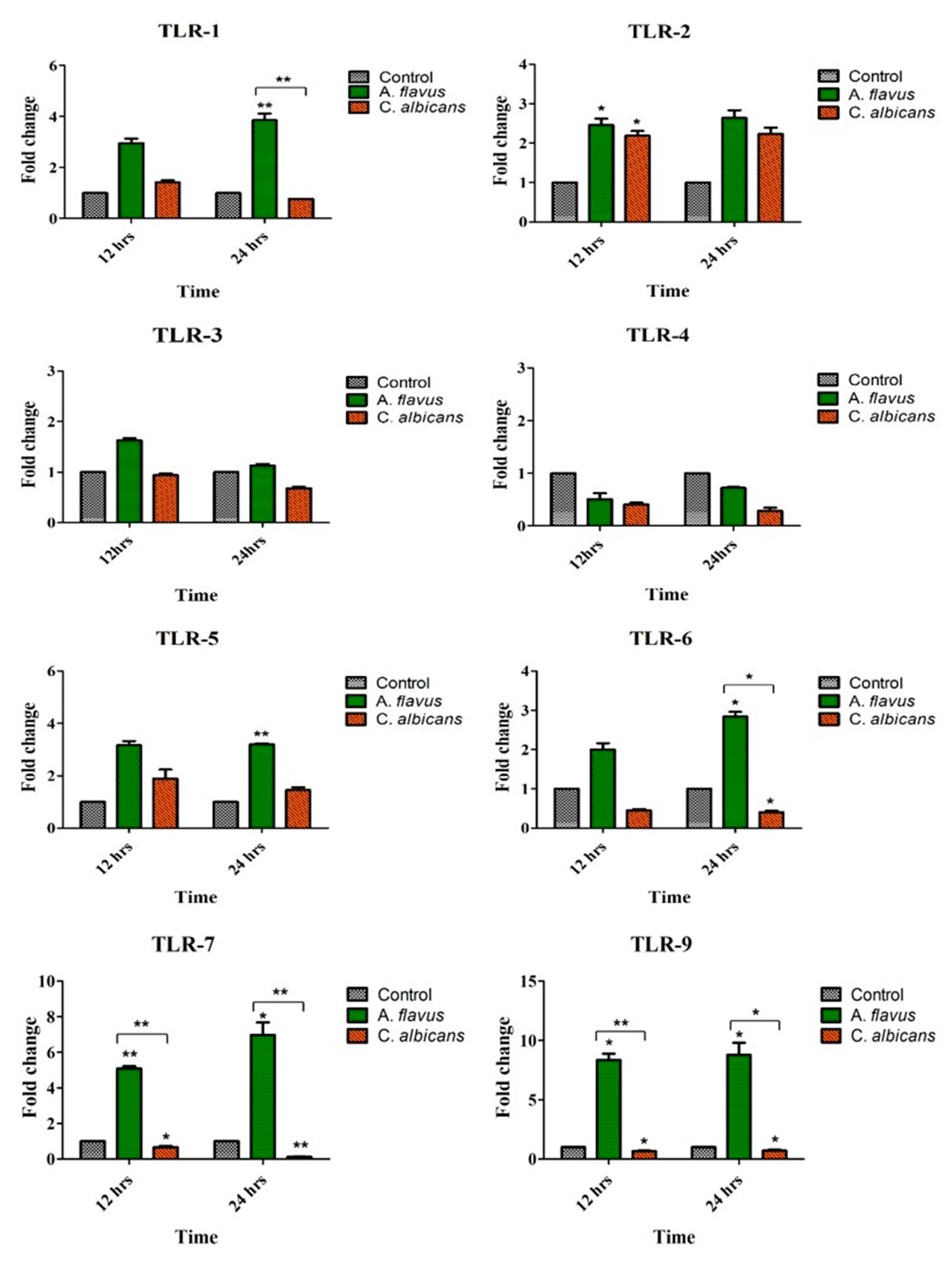
3.1. Toll-Like Receptors’ (TLRs) Expression Profile in Response to Aspergillus flavus and Candida albicans in Microglia
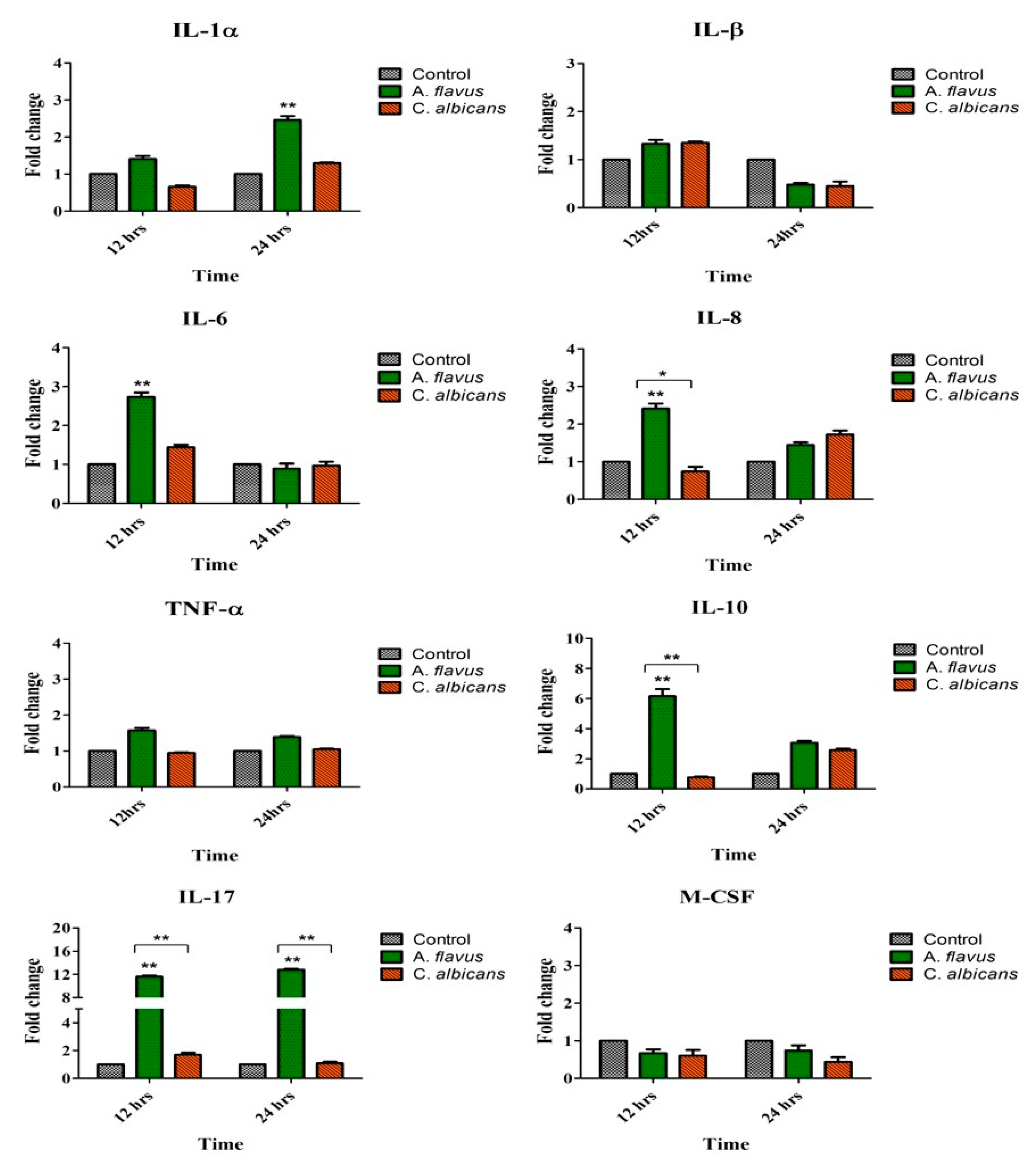
3.2. Induction of Immune Mediators by Microglia Challenged with A. flavus and C. albicans
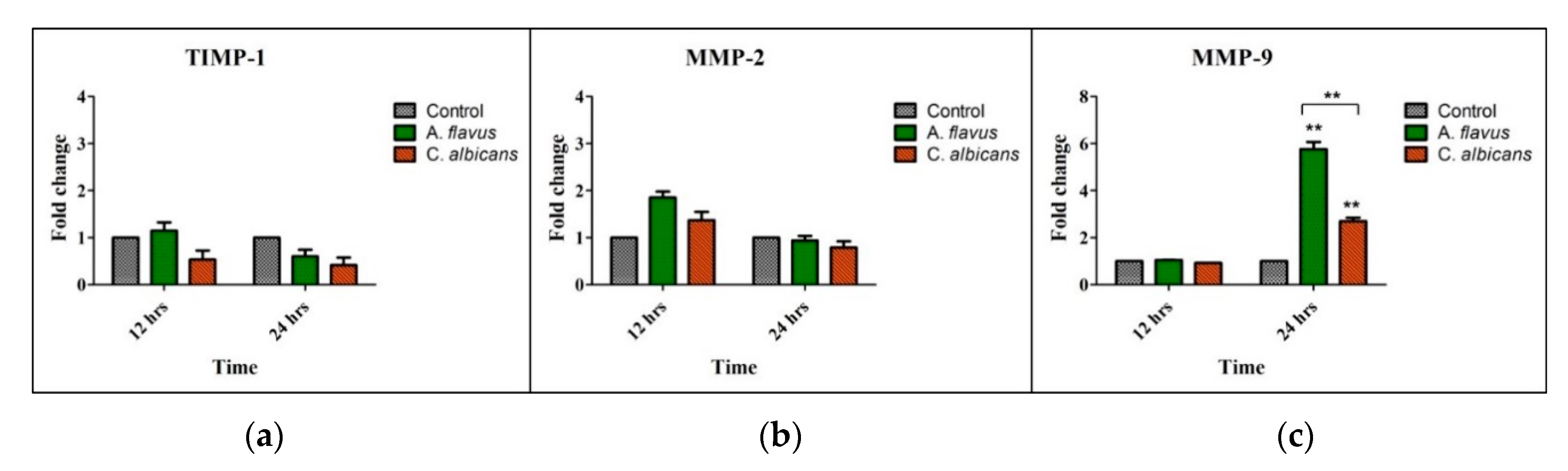
3.3. Expression of Tissue Remodeling Enzymes by A. flavus and C. albicans Infected Microglia
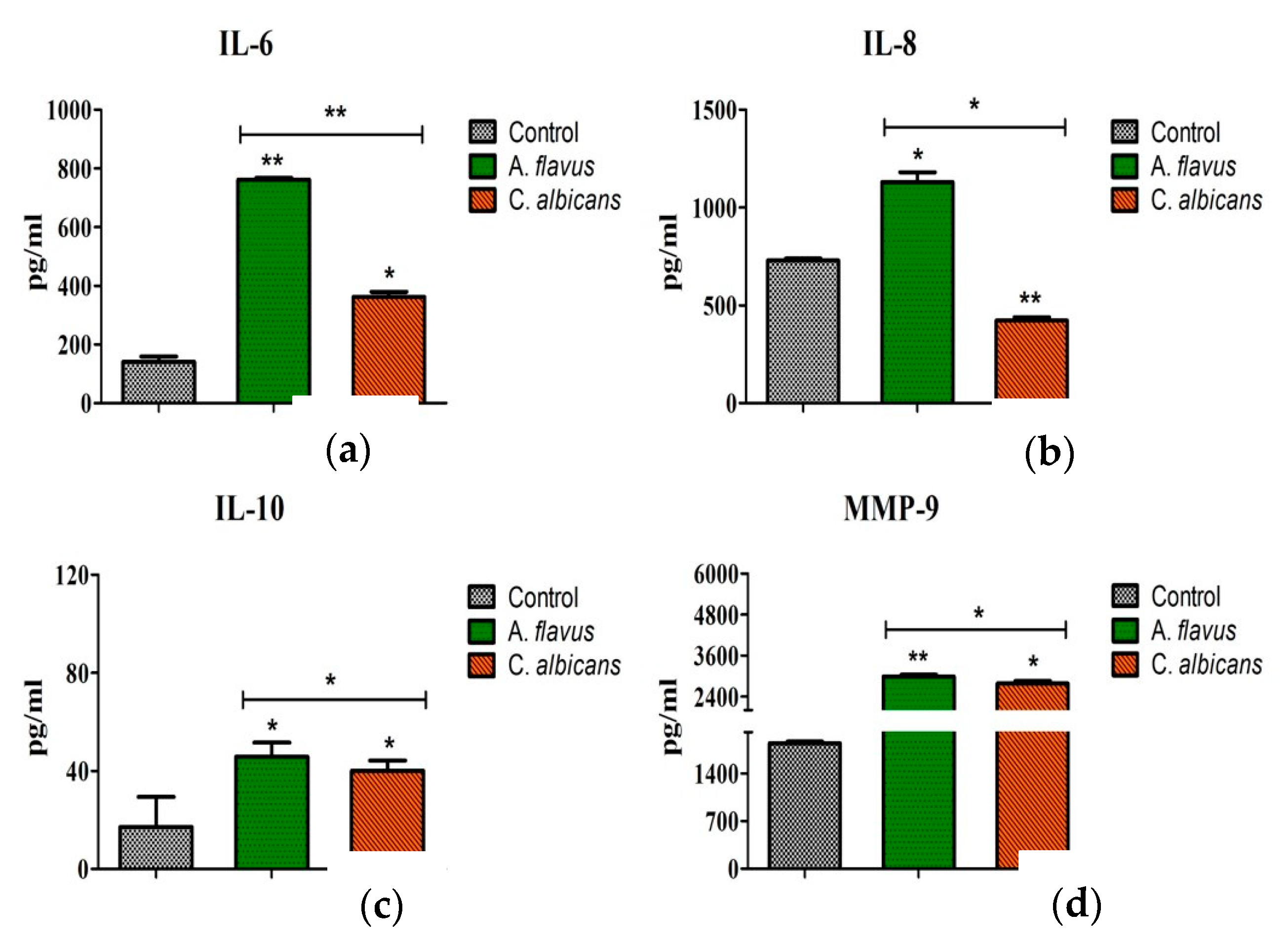
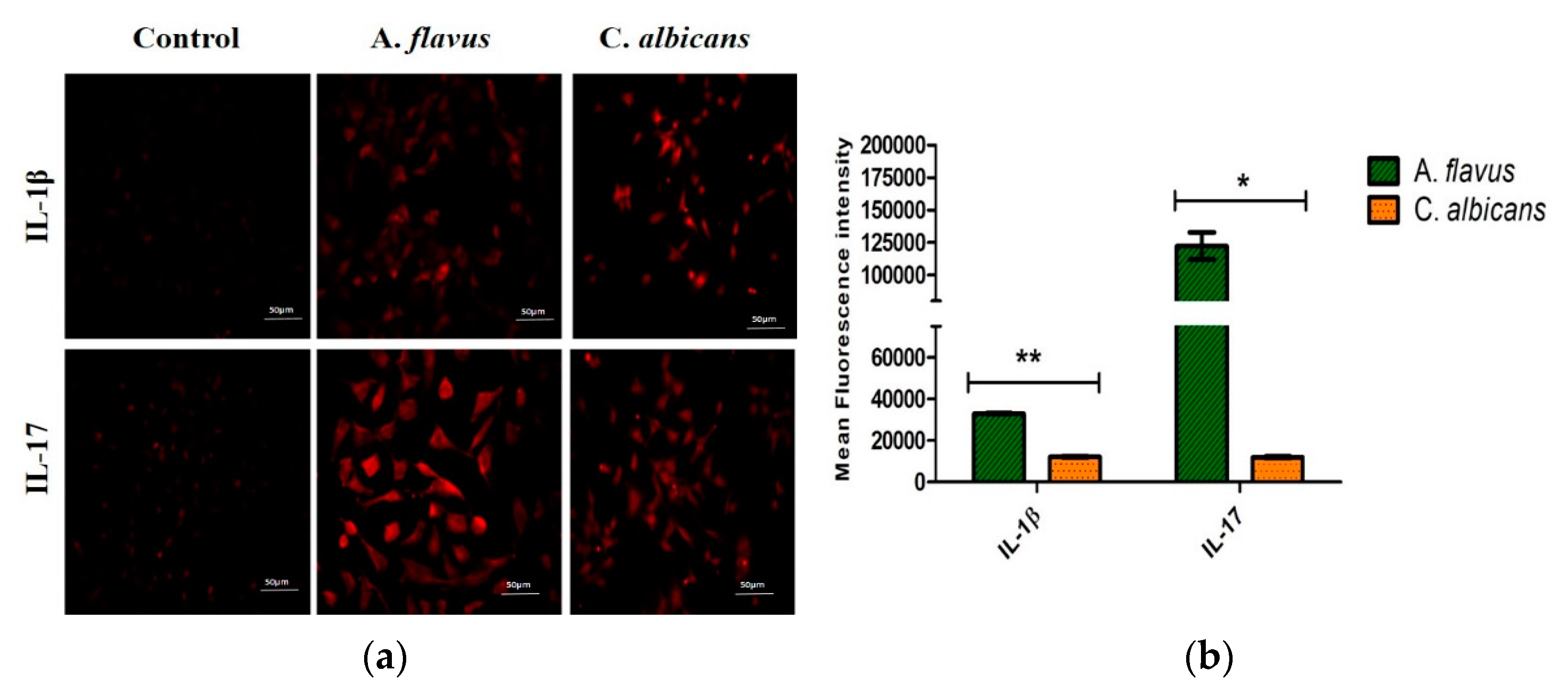
3.4. Protein Expression of Immune Mediators in Microglia Infected with Fungus
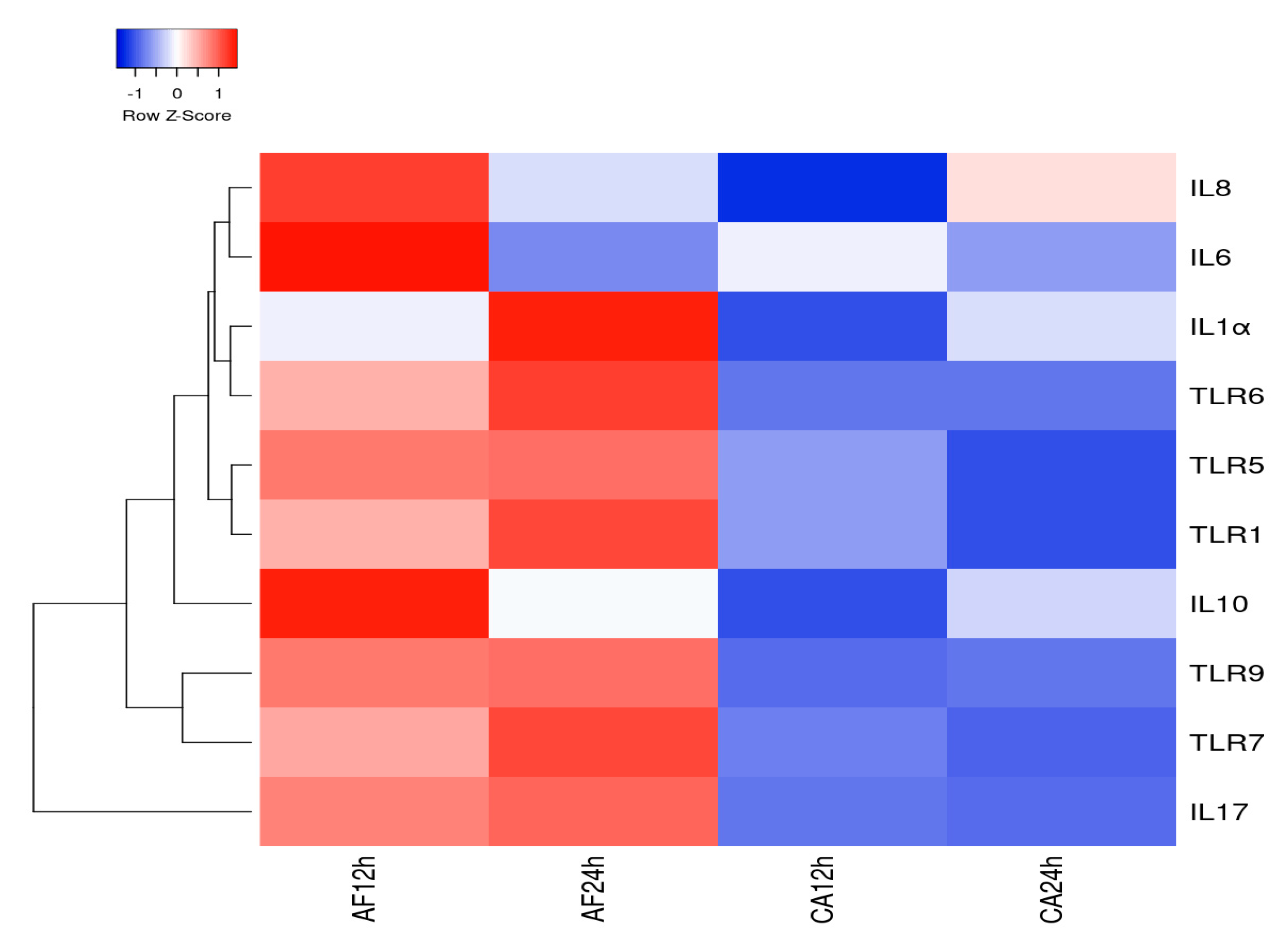
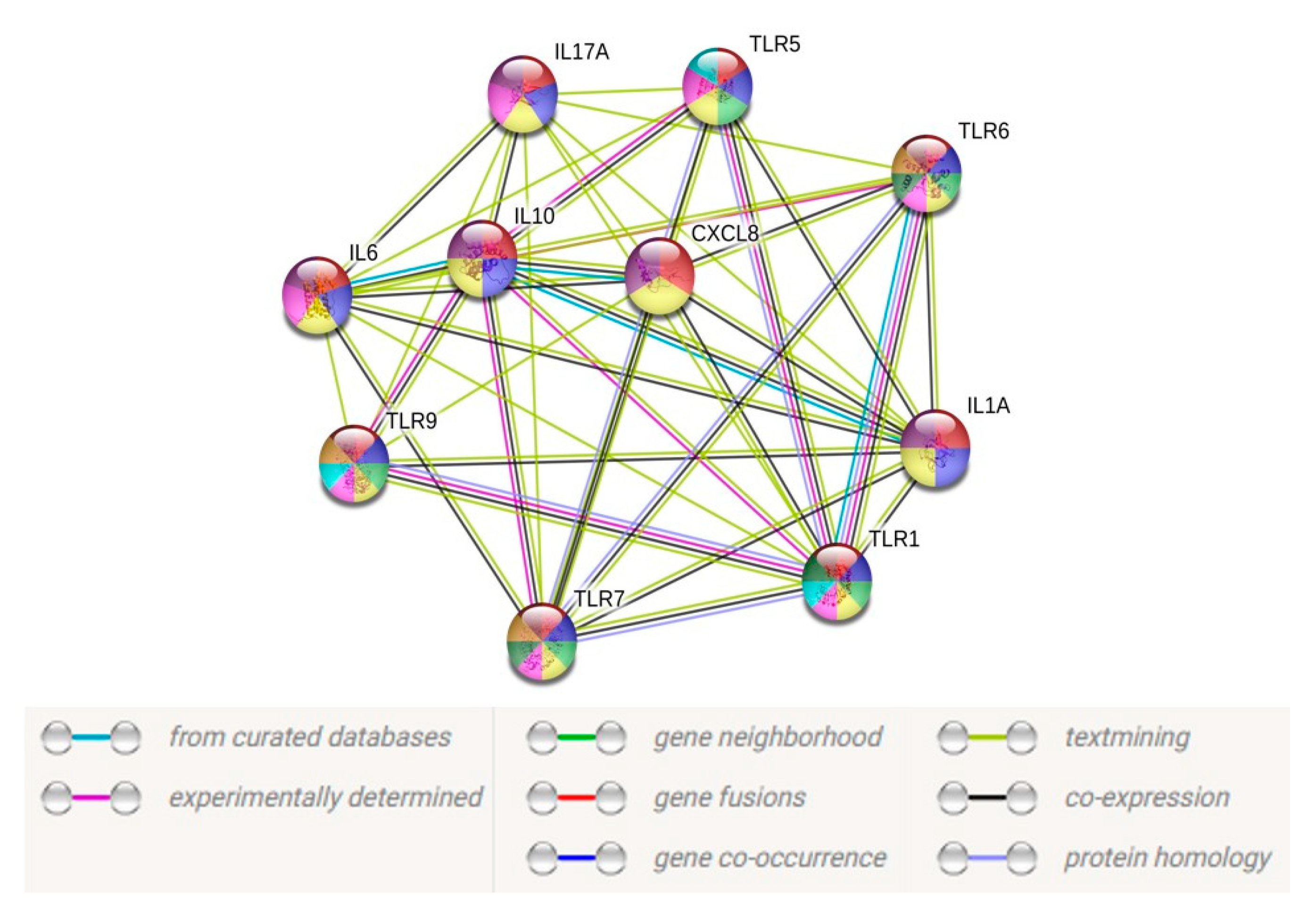
3.5. Heatmap and PPI Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Durand, M.L. Endophthalmitis. Clin. Microbiol. Infect. 2013, 19, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Kunimoto, D.Y.; Das, T.; Sharma, S.; Jalali, S.; Majji, A.B.; Gopinathan, U.; Athmanathan, S.; Rao, T.N. Microbiologic spectrum and susceptibility of isolates: Part I. Postoperative endophthalmitis. Endophthalmitis Research Group. Am. J. Ophthalmol. 1999, 128, 240–242. [Google Scholar] [CrossRef]
- Gandhi, J.; Jayasudha, R.; Naik, P.; Sharma, S.; Dave, V.P.; Joseph, J. Targeted high-throughput sequencing identifies predominantly fungal pathogens in patients with clinically infectious, culture-negative endophthalmitis in South India. Microorganisms 2019, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Shivaprakash, M.R.; Singh, R.; Tarai, B.; George, V.K.; Fomda, B.A.; Gupta, A. Fungal endophthalmitis: Fourteen years’ experience from a center in India. Retina 2008, 28, 1400–1407. [Google Scholar] [CrossRef]
- Usai, D.; Donadu, M.; Bua, A.; Molicotti, P.; Zanetti, S.; Piras, S.; Corona, P.; Ibba, R.; Carta, A. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J. Infect. Dev. Ctries. 2019, 13, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Bui, D.K.; Carvounis, P.E. Evidence for and against intravitreous corticosteroids in addition to intravitreous antibiotics for acute endophthalmitis. Int. Ophthalmol. Clin. 2014, 54, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, K.E. TNFalpha and MIP-2: Role in particle-induced inflammation and regulation by oxidative stress. Toxicol. Lett. 2000, 112–113, 177–183. [Google Scholar] [CrossRef]
- Hernandez-Chavez, M.J.; Perez-Garcia, L.A.; Nino-Vega, G.A.; Mora-Montes, H.M. Fungal strategies to evade the host immune recognition. J. Fungi (Basel) 2017, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochan, T.; Singla, A.; Tosi, J.; Kumar, A. Toll-like receptor 2 ligand pretreatment attenuates retinal microglial inflammatory response but enhances phagocytic activity toward Staphylococcus aureus. Infect. Immun. 2012, 80, 2076–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsouras, G.W.; Ramos, R.L.; Martinez, L.R. Role of microglia in fungal infections of the central nervous system. Virulence 2017, 8, 705–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo-Garcia, J.L.; Asencio-Duran, M.; Pastora-Salvador, N.; Vinciguerra, P.; Romano, M.R. Role of inflammation in endophthalmitis. Mediat. Inflamm. 2012, 2012, 196094. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Eter, N.; Heiduschka, P. The microglia in healthy and diseased retina. Exp. Eye Res. 2015, 136, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Sontam, B.; Guda, S.J.M.; Gandhi, J.; Sharma, S.; Tyagi, M.; Dave, V.P.; Das, T. Trends in microbiological spectrum of endophthalmitis at a single tertiary care ophthalmic hospital in India: A review of 25 years. Eye (London) 2019, 33, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Singh, P.K.; Revankar, S.G.; Chandrasekar, P.H.; Kumar, A. Pathobiology of Aspergillus fumigatus endophthalmitis in immunocompetent and immunocompromised mice. Microorganisms 2019, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Cramer, R.A.; Rivera, A.; Hohl, T.M. Immune responses against Aspergillus fumigatus: What have we learned? Curr. Opin. Infect. Dis. 2011, 24, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S.; et al. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on Th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef] [Green Version]
- Dello Russo, C.; Cappoli, N.; Coletta, I.; Mezzogori, D.; Paciello, F.; Pozzoli, G.; Navarra, P.; Battaglia, A. The human microglial HMC3 cell line: Where do we stand? A systematic literature review. J. Neuroinflamm. 2018, 15, 259. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Meis, J.F.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat. Rec. 2013, 296, 378–381. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.L. Bacterial and fungal endophthalmitis. Clin. Microbiol. Rev. 2017, 30, 597–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essman, T.F.; Flynn, H.W., Jr.; Smiddy, W.E.; Brod, R.D.; Murray, T.G.; Davis, J.L.; Rubsamen, P.E. Treatment outcomes in a 10-year study of endogenous fungal endophthalmitis. Ophthalmic Surg. Lasers Imaging Retin. 1997, 28, 185–194. [Google Scholar]
- Guest, J.M.; Singh, P.K.; Revankar, S.G.; Chandrasekar, P.H.; Kumar, A. Isavuconazole for treatment of experimental fungal endophthalmitis caused by Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.J.; Cerar, D.; Anderson, M.J.; Albarrag, A.; Fisher, M.C.; Pasqualotto, A.C.; Laverdiere, M.; Arendrup, M.C.; Perlin, D.S.; Denning, D.W. Frequency and evolution of Azole resistance in Aspergillus fumigatus associated with treatment failure. Emerg. Infect. Dis. 2009, 15, 1068–1076. [Google Scholar] [CrossRef]
- Horn, F.; Heinekamp, T.; Kniemeyer, O.; Pollmacher, J.; Valiante, V.; Brakhage, A.A. Systems biology of fungal infection. Front. Microbiol. 2012, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Maneu, V.; Noailles, A.; Megias, J.; Gomez-Vicente, V.; Carpena, N.; Gil, M.L.; Gozalbo, D.; Cuenca, N. Retinal microglia are activated by systemic fungal infection. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3578–3585. [Google Scholar] [CrossRef]
- Shah, V.B.; Huang, Y.; Keshwara, R.; Ozment-Skelton, T.; Williams, D.L.; Keshvara, L. Beta-glucan activates microglia without inducing cytokine production in Dectin-1-dependent manner. J. Immunol. 2008, 180, 2777–2785. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.B.; Williams, D.L.; Keshvara, L. beta-Glucan attenuates TLR2- and TLR4-mediated cytokine production by microglia. Neurosci. Lett. 2009, 458, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Galan-Diez, M.; Arana, D.M.; Serrano-Gomez, D.; Kremer, L.; Casasnovas, J.M.; Ortega, M.; Cuesta-Dominguez, A.; Corbi, A.L.; Pla, J.; Fernandez-Ruiz, E. Candida albicans beta-glucan exposure is controlled by the fungal CEK1-mediated mitogen-activated protein kinase pathway that modulates immune responses triggered through dectin-1. Infect. Immun. 2010, 78, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Toth, R.; Gacser, A. Mechanisms of pathogenic Candida species to evade the host complement attack. Front. Cell. Infect. Microbiol. 2020, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Kinjo, Y.; Ueno, K.; Takatsuka, S.; Nakamura, S.; Ogura, S.; Kimura, M.; Araoka, H.; Sadamoto, S.; Shinozaki, M.; et al. Differences in ocular complications between Candida albicans and non-albicans Candida infection analyzed by epidemiology and a mouse ocular candidiasis model. Front. Microbiol. 2018, 9, 2477. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2004, 4, 1–23. [Google Scholar] [CrossRef]
- Yang, I.; Han, S.J.; Kaur, G.; Crane, C.; Parsa, A.T. The role of microglia in central nervous system immunity and glioma immunology. J. Clin. Neurosci. 2010, 17, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Ramaprakash, H.; Ito, T.; Standiford, T.J.; Kunkel, S.L.; Hogaboam, C.M. Toll-like receptor 9 modulates immune responses to Aspergillus fumigatus conidia in immunodeficient and allergic mice. Infect. Immun. 2009, 77, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, C.; Kuchler, K. Fungal pathogens-a sweet and sour treat for toll-like receptors. Front. Cell. Infect. Microbiol. 2012, 2, 142. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Gow, N.A.; Munro, C.A.; Bates, S.; Collins, C.; Ferwerda, G.; Hobson, R.P.; Bertram, G.; Hughes, H.B.; Jansen, T.; et al. Immune sensing of Candida albicans requires cooperative recognition of mannans and glucans by lectin and Toll-like receptors. J. Clin. Investig. 2006, 116, 1642–1650. [Google Scholar] [CrossRef]
- Netea, M.G.; Warris, A.; Van der Meer, J.W.; Fenton, M.J.; Verver-Janssen, T.J.; Jacobs, L.E.; Andresen, T.; Verweij, P.E.; Kullberg, B.J. Aspergillus fumigatus evades immune recognition during germination through loss of toll-like receptor-4-mediated signal transduction. J. Infectig. Dis. 2003, 188, 320–326. [Google Scholar] [CrossRef] [Green Version]
- van de Veerdonk, F.L.; Kullberg, B.J.; van der Meer, J.W.; Gow, N.A.; Netea, M.G. Host-microbe interactions: Innate pattern recognition of fungal pathogens. Curr. Opin. Microbiol. 2008, 11, 305–312. [Google Scholar] [CrossRef]
- Chai, L.Y.; Vonk, A.G.; Kullberg, B.J.; Verweij, P.E.; Verschueren, I.; van der Meer, J.W.; Joosten, L.A.; Latge, J.P.; Netea, M.G. Aspergillus fumigatus cell wall components differentially modulate host TLR2 and TLR4 responses. Microbes Infect. 2011, 13, 151–159. [Google Scholar] [CrossRef]
- Orsi, C.F.; Borghi, E.; Colombari, B.; Neglia, R.G.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida albicans hyphal wall protein 1 (HWP1) genotype on biofilm production and fungal susceptibility to microglial cells. Microb. Pathog. 2014, 69–70, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, A.; Blatzer, M.; Posch, W.; Schubert, R.; Lass-Florl, C.; Schmidt, S.; Lehrnbecher, T. Aspergillus fumigatus responds to natural killer (NK) cells with upregulation of stress related genes and inhibits the immunoregulatory function of NK cells. Oncotarget 2016, 7, 71062–71071. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, B.G.; Conti, H.R. The role of IL-17 in protection against mucosal Candida infections. J. Fungi (Basel) 2017, 3, 52. [Google Scholar] [CrossRef]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Salazar, F.; Brown, G.D. Antifungal innate immunity: A perspective from the last 10 years. J. Innate Immun. 2018, 10, 373–397. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef]
- Li, F.; Jiang, D.; Samuel, M.A. Microglia in the developing retina. Neural Dev. 2019, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xu, Z.; Xiong, S.; Qin, G.; Sun, F.; Yang, J.; Yuan, T.F.; Zhao, L.; Wang, K.; Liang, Y.X.; et al. Dual extra-retinal origins of microglia in the model of retinal microglia repopulation. Cell Discov. 2018, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-cell RNA sequencing of microglia throughout the mouse lifespan and in the injured brain reveals complex cell-state changes. Immunity 2019, 50, 253–271.e2. [Google Scholar] [CrossRef] [Green Version]
- O’Koren, E.G.; Yu, C.; Klingeborn, M.; Wong, A.Y.W.; Prigge, C.L.; Mathew, R.; Kalnitsky, J.; Msallam, R.A.; Silvin, A.; Kay, J.N.; et al. Microglial function is distinct in different anatomical locations during retinal homeostasis and degeneration. Immunity 2019, 50, 723–737.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannas, S.; Usai, D.; Pinna, A.; Benvenuti, S.; Tardugno, R.; Donadu, M.; Zanetti, S.; Kaliamurthy, J.; Molicotti, P. Essential oils in ocular pathology: An experimental study. J. Infect. Dev. Ctries. 2015, 9, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadu, M.G.; Usai, D.; Marchetti, M.; Usai, M.; Mazzarello, V.; Molicotti, P.; Montesu, M.A.; Delogu, G.; Zanetti, S. Antifungal activity of oils macerates of North Sardinia plants against Candida species isolated from clinical patients with candidiasis. Nat. Prod. Res. 2019, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bua, A.; Usai, D.; Donadu, M.G.; Delgado Ospina, J.; Paparella, A.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Zanetti, S.; Molicotti, P. Antimicrobial activity of Austroeupatorium inulaefolium (H.B.K.) against intracellular and extracellular organisms. Nat. Prod. Res. 2018, 32, 2869–2871. [Google Scholar] [CrossRef]
- Chaves-Lopez, C.; Usai, D.; Donadu, M.G.; Serio, A.; Gonzalez-Mina, R.T.; Simeoni, M.C.; Molicotti, P.; Zanetti, S.; Pinna, A.; Paparella, A. Potential of Borojoa patinoi Cuatrecasas water extract to inhibit nosocomial antibiotic resistant bacteria and cancer cell proliferation in vitro. Food Funct. 2018, 9, 2725–2734. [Google Scholar] [CrossRef]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; D’Amico-Ricci, G.; Boscia, F.; Zanetti, S. In vitro antimicrobial activity of a new ophthalmic solution containing povidone-iodine 0.6% (IODIM®). Acta Ophthalmol. 2020, 98, e178–e180. [Google Scholar] [CrossRef]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; Boscia, F.; Zanetti, S. In vitro antimicrobial activity of a new ophthalmic solution containing hexamidine diisethionate 0.05% (Keratosept). Cornea 2020. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Characteristics | A. flavus | C. albicans |
|---|---|---|
| Age | 66 years | 54 years |
| Gender | Female | Male |
| Initial Visual Acuity | HM+ | 20/80p |
| Final Visual Acuity | HM+ | PL + PR Inaccurate |
| Microbiology | Aspergillus flavus | Candida albicans |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandhi, J.; Naik, P.; Kaur, I.; Kumar, A.; Joseph, J. Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis. J. Fungi 2020, 6, 162. https://doi.org/10.3390/jof6030162
Gandhi J, Naik P, Kaur I, Kumar A, Joseph J. Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis. Journal of Fungi. 2020; 6(3):162. https://doi.org/10.3390/jof6030162
Chicago/Turabian StyleGandhi, Jaishree, Poonam Naik, Inderjeet Kaur, Ashok Kumar, and Joveeta Joseph. 2020. "Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis" Journal of Fungi 6, no. 3: 162. https://doi.org/10.3390/jof6030162
APA StyleGandhi, J., Naik, P., Kaur, I., Kumar, A., & Joseph, J. (2020). Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis. Journal of Fungi, 6(3), 162. https://doi.org/10.3390/jof6030162