Clonal Expansion of Environmental Triazole Resistant Aspergillus fumigatus in Iran

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolate Collection
2.2. In Vitro Antifungal Susceptibility Testing
2.3. Detection of Cyp51a Gene Mutations
2.4. Microsatellite Genotyping
3. Results
3.1. Triazole Resistant A. fumigatus with Mutation in cyp51A Gene
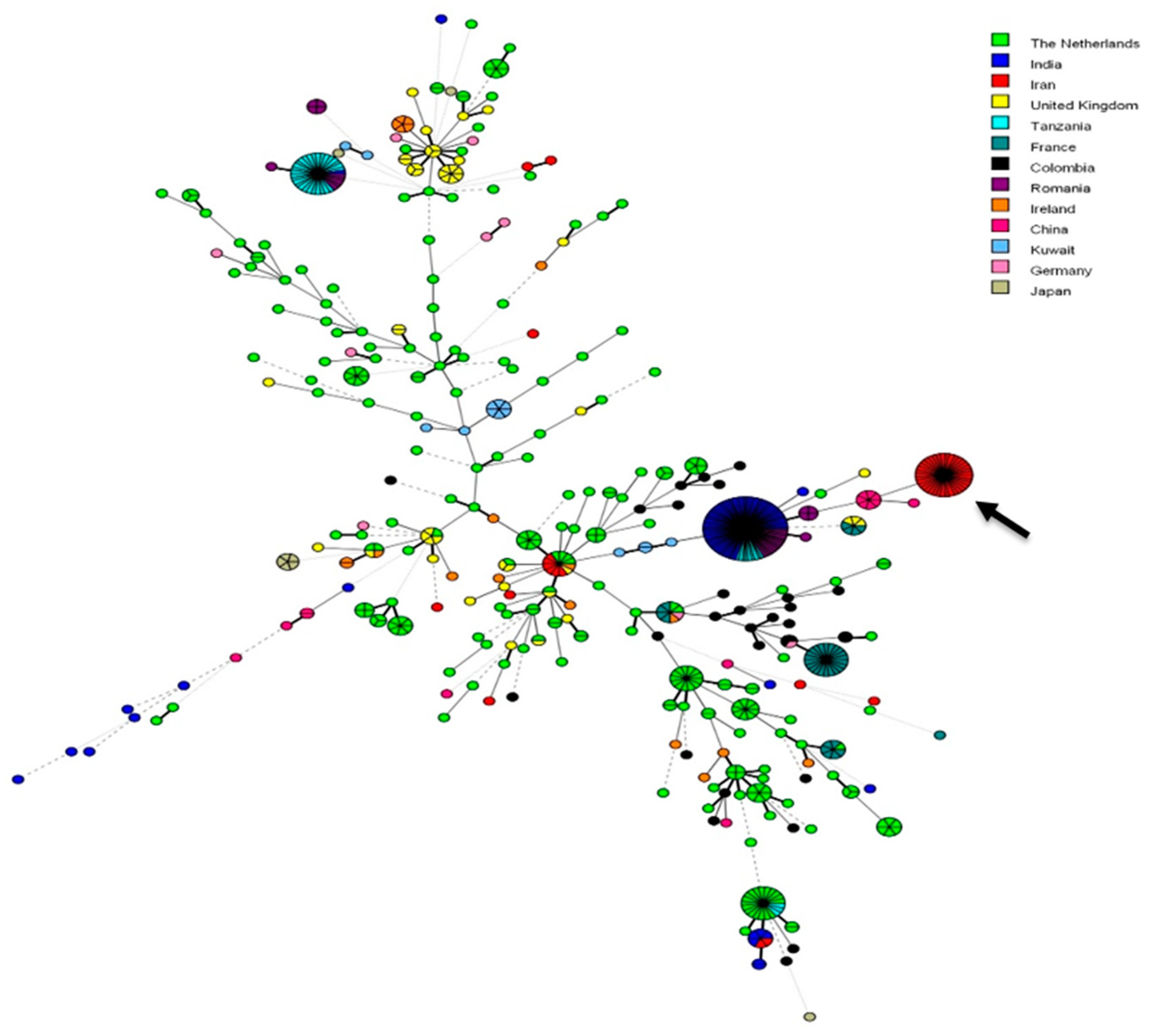
3.2. Microsatellite Typing Results and Evidence for Clonal Spread of a Single Triazole-Resistant A. fumigatus Genotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24 (Suppl. 1), e1–e38. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Sharma, B.; Mahto, K.K.; Meis, J.F.; Chowdhary, A. High-Frequency Direct Detection of Triazole Resistance in Aspergillus fumigatus from Patients with Chronic Pulmonary Fungal Diseases in India. J. Fungi 2020, 6, 67. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.; Meis, J.F. Azole Resistance in Aspergillus fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, C.H.; Gibbons, J.G.; Fedorova, N.D.; Meis, J.F.; Rokas, A. Evidence for genetic differentiation and variable recombination rates among Dutch populations of the opportunistic human pathogen Aspergillus fumigatus. Mol. Ecol. 2012, 21, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole Resistance in Aspergillus spp.: A Worldwide Problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Lestrade, P.P.A.; Meis, J.F.; Melchers, W.J.G.; Verweij, P.E. Triazole resistance in Aspergillus fumigatus: Recent insights and challenges for patient management. Clin. Microbiol. Infect. 2019, 25, 799–806. [Google Scholar] [CrossRef]
- Resendiz-Sharpe, A.; Mercier, T.; Lestrade, P.P.A.; van der Beek, M.T.; von dem Borne, P.A.; Cornelissen, J.J.; De Kort, E.; Rijnders, B.J.A.; Schauwvlieghe, A.F.A.D.; Verweij, P.E.; et al. Prevalence of voriconazole-resistant invasive aspergillosis and its impact on mortality in haematology patients. J. Antimicrob. Chemother. 2019, 74, 2759–2766. [Google Scholar] [CrossRef]
- Buil, J.B.; Hagen, F.; Chowdhary, A.; Verweij, P.E.; Meis, J.F. Itraconazole, Voriconazole, and Posaconazole CLSI MIC Distributions for Wild-Type and Azole-Resistant Aspergillus fumigatus Isolates. J. Fungi 2018, 4, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; van Dijk, K.; Hagen, F.; van der Beek, M.; Kampinga, G.A.; Zoll, J.; et al. A Novel Environmental Azole Resistance Mutation in Aspergillus fumigatus and a Possible Role of Sexual Reproduction in Its Emergence. mBio 2017, 8, e00791-17. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Meis, J.F. Emergence of azole resistant Aspergillus fumigatus and One Health: Time to implement environmental stewardship. Environ. Microbiol. 2018, 20, 1299–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trovato, L.; Scalia, G.; Domina, M.; Oliveri, S. Environmental Isolates of Multi-Azole-Resistant Aspergillus spp. in Southern Italy. J. Fungi 2018, 4, 131. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Sharma, C.; Sundar, G.; Singh, P.K.; Gaur, S.H.N.; Hagen, F.; Klaassen, C.H.; Meis, J.F. Clonal expansion and emergence of environmental multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR₃₄/L98H mutations in the cyp51A gene in India. PLoS ONE 2012, 7, e52871. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P.; Verweij, P.E. Aspergillus fumigatus and pan-azole resistance: Who should be concerned? Curr. Opin. Infect. Dis. 2020, 33, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Badali, H.; Vaezi, A.; Haghani, I.; Yazdanparast, S.A.; Hedayati, M.T.; Mousavi, B.; Ansari, S.; Hagen, F.; Meis, J.F.; Chowdhary, A. Environmental study of azole-resistant Aspergillus fumigatus with TR34/L98H mutations in the cyp51A gene in Iran. Mycoses 2013, 56, 659–663. [Google Scholar] [CrossRef]
- Ahangarkani, F.; Puts, Y.; Nabili, M.; Khodavaisy, S.; Moazeni, M.; Salehi, Z.; Laal Kargar, M.; Badali, H.; Meis, J.F. First azole-resistant Aspergillus fumigatus isolates with the environmental TR46 /Y121F/T289A mutation in Iran. Mycoses 2020, 63, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Hashemi, S.J.; Zibafar, E.; Zoll, J.; Hedayati, M.T.; Mouton, J.W.; Melchers, W.J.G.; Verweij, P.E. Azole-resistant Aspergillus fumigatus, Iran. Emerg. Infect. Dis. 2013, 19, 832–834. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Debets, A.J.M.; Rijs, A.J.M.M.; Zhang, J.; Snelders, E.; Leendertse, P.C.; Melchers, W.J.G.; Rietveld, A.G.; Zwaan, B.J.; Verweij, P.E. Environmental Hotspots for Azole Resistance Selection of Aspergillus fumigatus, the Netherlands. Emerg. Infect. Dis. 2019, 25, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Sewell, T.R.; Zhang, Y.; Brackin, A.P.; Shelton, J.M.G.; Rhodes, J.; Fisher, M.C. Elevated Prevalence of Azole-Resistant Aspergillus fumigatus in Urban versus Rural Environments in the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00548-19. [Google Scholar] [CrossRef] [Green Version]
- Vaezi, A.; Fakhim, H.; Javidnia, J.; Khodavaisy, S.; Abtahian, Z.; Vojoodi, M.; Nourbakhsh, F.; Badali, H. Pesticide behavior in paddy fields and development of azole-resistant Aspergillus fumigatus: Should we be concerned? J. Mycol. Med. 2018, 28, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Vanni, A.; Fontana, F.; Gamberini, R.; Calabria, A. Occurrence of dicarboximide fungicides and their metabolites’ residues in commercial compost. Agronomie 2004, 24, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Camps, S.M.; Rijs, A.J.; Klaassen, C.H.; Meis, J.F.; O’Gorman, C.M.; Dyer, P.S.; Melchers, W.J.; Verweij, P.E. Molecular epidemiology of Aspergillus fumigatus isolates harboring the TR34/L98H azole resistance mechanism. J. Clin. Microbiol. 2012, 50, 2674–2680. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Azole-resistant Aspergillus fumigatus with the environmental TR46/Y121F/T289A mutation in India. J. Antimicrob. Chemother. 2014, 69, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; CLSI standard M38; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Klaassen, C.H.; de Valk, H.A.; Curfs-Breuker, I.M.; Meis, J.F. Novel mixed-format real-time PCR assay to detect mutations conferring resistance to triazoles in Aspergillus fumigatus and prevalence of multi-triazole resistance among clinical isolates in the Netherlands. J. Antimicrob. Chemother. 2010, 65, 901–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavergne, R.A.; Morio, F.; Favennec, L.; Dominique, S.; Meis, J.F.; Gargala, G.; Verweij, P.E.; Le Pape, P. First description of azole-resistant Aspergillus fumigatus due to TR46/Y121F/T289A mutation in France. Antimicrob. Agents Chemother. 2015, 59, 4331–4335. [Google Scholar] [CrossRef] [Green Version]
- de Valk, H.A.; Meis, J.F.; Curfs, I.M.; Muehlethaler, K.; Mouton, J.W.; Klaassen, C.H. Use of a novel panel of nine short tandem repeats for exact and high-resolution fingerprinting of Aspergillus fumigatus isolates. J. Clin. Microbiol. 2005, 43, 4112–4120. [Google Scholar] [CrossRef] [Green Version]
- Balajee, S.A.; de Valk, H.A.; Lasker, B.A.; Meis, J.F.; Klaassen, C.H.W. Utility of a microsatellite assay for identifying clonally related outbreak isolates of Aspergillus fumigatus. J. Microbiol. Methods 2008, 73, 252–256. [Google Scholar] [CrossRef]
- Guinea, J.; García de Viedma, D.; Peláez, T.; Escribano, P.; Muñoz, P.; Meis, J.F.; Klaassen, C.H.W.; Bouza, E. Molecular epidemiology of Aspergillus fumigatus: An in-depth genotypic analysis of isolates involved in an outbreak of invasive aspergillosis. J. Clin. Microbiol. 2011, 49, 3498–3503. [Google Scholar] [CrossRef] [Green Version]
- Nabili, M.; Shokohi, T.; Moazeni, M.; Khodavaisy, S.; Aliyali, M.; Badiee, P.; Zarrinfar, H.; Hagen, F.; Badali, H. High prevalence of clinical and environmental triazole-resistant Aspergillus fumigatus in Iran: Is it a challenging issue? J. Med. Microbiol. 2016, 65, 468–475. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-resistant aspergillosis: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216 (suppl. 3), S436–S444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, S.S. Global Aspects of Triazole Resistance in Aspergillus fumigatus with Focus on Latin American Countries. J. Fungi 2017, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Abdolrasouli, A.; Rhodes, J.; Beale, M.A.; Hagen, F.; Rogers, T.R.; Chowdhary, A.; Meis, J.F.; Armstrong-James, D.; Fisher, M.C. Genomic Context of Azole Resistance Mutations in Aspergillus fumigatus Determined Using Whole-Genome Sequencing. mBio 2015, 6, e00939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Rubio, R.; Escribano, P.; Gomez, A.; Guinea, J.; Mellado, E. Comparison of Two Highly Discriminatory Typing Methods to Analyze Aspergillus fumigatus Azole Resistance. Front. Microbiol. 2018, 9, 1626. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global Population Genetic Analysis of Aspergillus fumigatus. mSphere 2017, 2, e00019-17. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Tashiro, M.; Urano, R.; Kikuchi, M.; Ito, N.; Moriya, E.; Shirahige, T.; Mishima, M.; Takazono, T.; Miyazaki, T.; et al. Characteristics of azole-resistant Aspergillus fumigatus attached to agricultural products imported to Japan. J. Infect. Chemother. 2020, 26, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Dunne, K.; Hagen, F.; Pomeroy, N.; Meis, J.F.; Rogers, T.R. Intercountry transfer of triazole-resistant Aspergillus fumigatus on plant bulbs. Clin. Infect. Dis. 2017, 65, 147–149. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.H.; Hagen, F.; Astvad, K.M.; Tyron, A.; Meis, J.F.; Arendrup, M.C. Azole-resistant Aspergillus fumigatus in Denmark: A laboratory-based study on resistance mechanisms and genotypes. Clin. Microbiol. Infect. 2016, 22, 570-e1. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef] [Green Version]
- Özmerdiven, G.E.; Ak, S.; Ener, B.; Ağca, H.; Cilo, B.D.; Tunca, B.; Akalın, H. First determination of azole resistance in Aspergillus fumigatus strains carrying the TR34/L98H mutations in Turkey. J. Infect. Chemother. 2015, 21, 581–586. [Google Scholar] [CrossRef]
- Perveen, I.; Sehar, S.; Naz, I.; Ahmed, S. Prospective evaluation of azole resistance in Aspergillus fumigatus clinical isolates in Pakistan. In Proceedings of the 7th Advances Against Aspergillosis, Manchester, UK, 3–5 March 2016. [Google Scholar]


{kind=link}
| Strain | Longitude and Latitude of Sampling | MIC (mg/L) | 3 STRAf | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 ITR | 2 VOR | Mutation in cyp51A | 2A | 2B | 2C | 3A | 3B | 3C | 4A | 4B | 4C | ||
| mn224 | 35.9548° N, 52.1100° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn225 | 36.6717° N, 52.4439° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn226 | 36.6717° N, 52.4439° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn229 | 36.6717° N, 52.4439° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn227 | 36.7049° N, 52.6547° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn228 | 36.6329° N, 52.2667° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn231 | 36.4684° N, 52.8634° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn235 | 36.4684° N, 52.8634° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn232 | 36.4684° N, 52.8634° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn241 | 36.4684° N, 52.8634° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn233 | 36.6858° N, 52.5265° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn234 | 36.6858° N, 52.5265° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn236 | 36.4676° N, 52.3507° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn246 | 36.4676° N, 52.3507° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn247 | 36.5971° N, 52.6654° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn250 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn251 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn252 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn253 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn254 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn255 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn256 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn257 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn258 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn260 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn261 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn263 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn265 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn266 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn267 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn268 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn269 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn270 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn271 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn272 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn273 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn274 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn277 | 36.5659° N, 53.0586° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn279 | 36.9268° N, 50.6431° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn280 | 36.7284° N, 53.8102° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| mn281 | 36.7284° N, 53.8102° E | 16 | 2 | TR34/L98H | 22 | 10 | 9 | 9 | 9 | 23 | 8 | 10 | 8 |
| IFRC: 1854 | 35.6892° N, 51.3890° E | 16 | 2 | TR34/L98H | 14 | 10 | 8 | 9 | 10 | 5 | 8 | 10 | 27 |
| IFRC: 1858 | 35.6892° N, 51.3890° E | 8 | 1 | TR34/L98H | 13 | 21 | 8 | 32 | 9 | 6 | 8 | 10 | 10 |
| IFRC: 1866 | 35.6892° N, 51.3890° E | 16 | 8 | TR34/L98H | 14 | 24 | 14 | 31 | 9 | 31 | 10 | 9 | 5 |
| mn248 | 36.6329° N, 52.2667° E | 16 | 16 | M172V | 11 | 15 | 16 | 19 | 29 | 4 | 7 | 5 | 8 |
| IFRC: 1867 | 35.6892° N, 51.3890° E | 16 | 16 | M172V | 11 | 16 | 9 | 19 | 20 | 5 | 7 | 5 | 5 |
| IFRC: 1860 | 35.6892° N, 51.3890° E | 16 | 0.125 | Wild type | 27 | 18 | 16 | 7 | 12 | 28 | 27 | 5 | 8 |
| IFRC: 1868 | 35.6892° N, 51.3890° E | 16 | 16 | Wild type | 27 | 18 | 16 | 7 | 12 | 28 | 27 | 5 | 8 |
| IFRC: 1862 | 35.6892° N, 51.3890° E | 16 | 0.5 | Wild type | 27 | 20 | 13 | 8 | 14 | 35 | 10 | 11 | 10 |
| IFRC: 1864 | 35.6892° N, 51.3890° E | 16 | 0.25 | Wild type | 20 | 10 | 8 | 37 | 9 | 6 | 10 | 9 | 15 |
| IFRC: 1859 | 35.6892° N, 51.3890° E | 16 | 0.25 | Wild type | 21 | 20 | 14 | 30 | 21 | 5 | 11 | 6 | 5 |
| mn245 | 36.7049° N, 52.6547° E | 16 | 1 | Wild type | 13 | 19 | 8 | 34 | 29 | 7 | 10 | 9 | 8 |
| mn276 | 36.5659° N, 53.0586° E | 16 | 2 | Wild type | 18 | 22 | 15 | 43 | 13 | 27 | 13 | 8 | 10 |
| mn278 | 36.5659° N, 53.0586° E | 16 | 2 | Wild type | 24 | 10 | 10 | 28 | 11 | 6 | 8 | 7 | 15 |
| mn249 | 36.5659° N, 53.0586° E | 0.125 | 0.5 | Wild type | 23 | 22 | 14 | 44 | 12 | 27 | 13 | 8 | 7 |
| mn223 | 36.5659° N, 53.0586° E | 0.125 | 0.5 | Wild type | 22 | 23 | 11 | 9 | 10 | 6 | 11 | 7 | 6 |
| mn240 | 36.6329° N, 52.2667° E | 0.5 | 1 | Wild type | 11 | 15 | 16 | 19 | 29 | 4 | 7 | 5 | 5 |
| mn242 | 36.6329° N, 52.2667° E | 0.125 | 0.5 | Wild type | 24 | 22 | 18 | 24 | 13 | 17 | 9 | 8 | 10 |
| mn230 | 36.6717° N, 52.4439° E | 0.5 | 0.5 | Wild type | 24 | 20 | 18 | 22 | 10 | 6 | 9 | 12 | 9 |
| IFRC: 1863 | 35.6892° N, 51.3890° E | 4 | 0.125 | Wild type | 24 | 18 | 15 | 94 | 10 | 6 | 14 | 11 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahangarkani, F.; Badali, H.; Abbasi, K.; Nabili, M.; Khodavaisy, S.; de Groot, T.; Meis, J.F. Clonal Expansion of Environmental Triazole Resistant Aspergillus fumigatus in Iran. J. Fungi 2020, 6, 199. https://doi.org/10.3390/jof6040199
Ahangarkani F, Badali H, Abbasi K, Nabili M, Khodavaisy S, de Groot T, Meis JF. Clonal Expansion of Environmental Triazole Resistant Aspergillus fumigatus in Iran. Journal of Fungi. 2020; 6(4):199. https://doi.org/10.3390/jof6040199
Chicago/Turabian StyleAhangarkani, Fatemeh, Hamid Badali, Kiana Abbasi, Mojtaba Nabili, Sadegh Khodavaisy, Theun de Groot, and Jacques F. Meis. 2020. "Clonal Expansion of Environmental Triazole Resistant Aspergillus fumigatus in Iran" Journal of Fungi 6, no. 4: 199. https://doi.org/10.3390/jof6040199
APA StyleAhangarkani, F., Badali, H., Abbasi, K., Nabili, M., Khodavaisy, S., de Groot, T., & Meis, J. F. (2020). Clonal Expansion of Environmental Triazole Resistant Aspergillus fumigatus in Iran. Journal of Fungi, 6(4), 199. https://doi.org/10.3390/jof6040199