Efficacy of LAMB against Emerging Azole- and Multidrug-Resistant Candida parapsilosis Isolates in the Galleria mellonella Model

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. C. parapsilosis Isolates and Clinical Data
2.2. Galleria mellonella In Vivo Treatment Studies
3. Results
3.1. In Vitro Antifungal Susceptibility Pattern and Alterations in ERG11 or FKS1
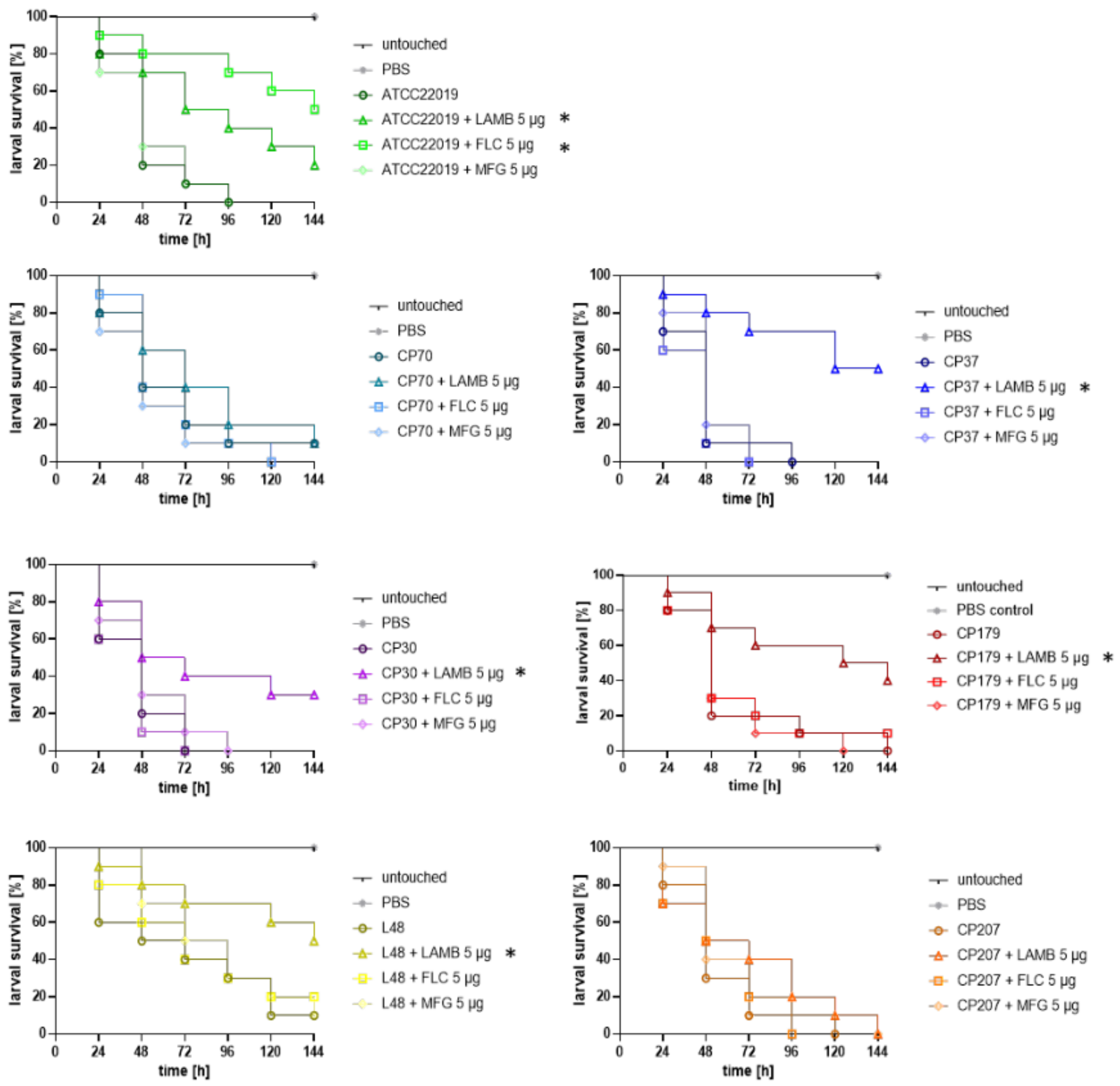
3.2. In Vivo Efficacy of Antifungal Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species From 1997–2016. Open Forum Infect. Dis. 2019, 6, S79–S94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.; Ahmad, S.; Al-Sweih, N.; Mokaddas, E.; Al-Banwan, K.; Alfouzan, W.; Al-Obaid, I.; Al-Obaid, K.; Asadzadeh, M.; Jeragh, A.; et al. Changing trends in epidemiology and antifungal susceptibility patterns of six bloodstream Candida species isolates over a 12-year period in Kuwait. PLoS ONE 2019, 14, e0216250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Chen, S.C.-A.; Kong, F.; Xu, X.-L.; Yan, L.; Kong, H.-S.; Fan, X.; Hou, X.; Cheng, J.-W.; Zhou, M.-L.; et al. Distribution and Antifungal Susceptibility of Candida Species Causing Candidemia in China: An Update From the CHIF-NET Study. J. Infect. Dis. 2020, 221, S139–S147. [Google Scholar] [CrossRef] [PubMed]
- Kakeya, H.; Shibata, W.; Yamada, K.; Kaneko, Y. National Trends in the Japanese Distribution of Major Candida Species Causing Candidemia During 2003–2017: A Report by the. Open Forum Infect. Dis. 2019, 40. [Google Scholar]
- Puig-Asensio, M.; Padilla, B.; Garnacho-Montero, J.; Zaragoza, O.; Aguado, J.; Montejo, M.; Muñoz, P.; Ruiz-Camps, I.; Cuenca-Estrella, M.; Almirante, B. Epidemiology and predictive factors for early and late mortality in Candida bloodstream infections: A population-based surveillance in Spain. Clin. Microbiol. Infect. 2014, 20, O245–O254. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, G.; Velegraki, A.; Falagas, M.E. A 10-Year Survey of Antifungal Susceptibility of Candidemia Isolates from Intensive Care Unit Patients in Greece. Antimicrob. Agents Chemother. 2008, 53, 1242–1244. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou-Olivgeris, M.; Spiliopoulou, A.; Kolonitsiou, F.; Bartzavali, C.; Lambropoulou, A.; Xaplanteri, P.; Anastassiou, E.D.; Marangos, M.; Spiliopoulou, I.; Christofidou, M. Increasing incidence of candidaemia and shifting epidemiology in favor of Candida non-albicans in a 9-year period (2009–2017) in a university Greek hospital. Infection 2019, 47, 209–216. [Google Scholar] [CrossRef]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of azole-resistant Candida parapsilosis causing bloodstream infection: Results from laboratory-based sentinel surveillance in South Africa. J. Antimicrob. Chemother. 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of Candidemia in Latin America: A Laboratory-Based Survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef] [Green Version]
- Pinhati, H.M.S.; Casulari, L.A.; Souza, A.C.R.; Siqueira, R.A.; Damasceno, C.M.G.; Colombo, A.L. Outbreak of candidemia caused by fluconazole resistant Candida parapsilosis strains in an intensive care unit. BMC Infect. Dis. 2016, 16, 433. [Google Scholar] [CrossRef]
- Grossman, N.T.; Pham, C.D.; Cleveland, A.A.; Lockhart, S.R. Molecular Mechanisms of Fluconazole Resistance in Candida parapsilosis Isolates from a U.S. Surveillance System. Antimicrob. Agents Chemother. 2014, 59, 1030–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Kim, Y.-J.; Yong, D.; Byun, J.-H.; Kim, T.S.; Chang, Y.S.; Choi, M.J.; Byeon, S.A.; Won, E.J.; Kim, S.H.; et al. Fluconazole-Resistant Candida parapsilosis Bloodstream Isolates with Y132F Mutation in ERG11 Gene, South Korea. Emerg. Infect. Dis. 2018, 24, 1768–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomaz, D.Y.; De Almeida, J.N.J.; Lima, G.M.E.; Nunes, M.D.O.; Camargo, C.H.; Grenfell, R.D.C.; Benard, G.; Del Negro, G.M.B. An Azole-Resistant Candida parapsilosis Outbreak: Clonal Persistence in the Intensive Care Unit of a Brazilian Teaching Hospital. Front. Microbiol. 2018, 9, 2997. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, P.K.; De Groot, T.; Kumar, A.; Mathur, P.; Tarai, B.; Sachdeva, N.; Upadhyaya, G.; Sarma, S.; Meis, J.F.; et al. Emergence of clonal fluconazole-resistant Candida parapsilosis clinical isolates in a multicentre laboratory-based surveillance study in India. J. Antimicrob. Chemother. 2019, 74, 1260–1268. [Google Scholar] [CrossRef] [Green Version]
- Martini, C.; Torelli, R.; De Groot, T.; De Carolis, E.; Morandotti, G.A.; De Angelis, G.; Posteraro, B.; Meis, J.F.; Sanguinetti, M. Prevalence and Clonal Distribution of Azole-Resistant Candida parapsilosis Isolates Causing Bloodstream Infections in a Large Italian Hospital. Front. Cell. Infect. Microbiol. 2020, 10, 232. [Google Scholar] [CrossRef]
- Magobo, R.E.; Lockhart, S.R.; Govender, N.P.; Wadula, J.; Rensburg, V.; Van Rensburg, C.J.; Whitelaw, A.; Zietsman, I.; Miller, N.; Smith, P.; et al. Fluconazole-resistant Candida parapsilosis strains with a Y132F substitution in the ERG11 gene causing invasive infections in a neonatal unit, South Africa. Mycoses 2020, 63, 471–477. [Google Scholar] [CrossRef]
- Arastehfar, A.; Hilmioğlu-Polat, S.; Daneshnia, F.; Pan, W.; Hafez, A.; Fang, W.; Liao, W.; Şahbudak-Bal, Z.; Metin, D.Y.; de Almeida, J.N., Jr.; et al. Clonal candidemia outbreak by Candida parapsilosis carrying Y132F in Turkey: Evolution of a persisting challenge. J. Fungi 2020. under review. [Google Scholar]
- Arastehfar, A.; Daneshnia, F.; Hilmioğlu-Polat, S.; Fang, W.; Yaşar, M.; Polat, F.; Yeşim Metin, D.; Rigole, P.; Coenye, T.; Ilkit, M.; et al. Frist report of candidemia clonal outbreak caused by emerging fluconazole-resistant Candida parapsilosis isolates harboring Y132F and/or Y132F+K143R in Turkey. Antimicrob. Agents Chemother. 2020, 64, e01001-20. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, M.; Watts, M.R.; Wang, H.; Fan, X.; Kong, F.; Xu, Y.-C. Development of fluconazole resistance in a series of Candida parapsilosis isolates from a persistent candidemia patient with prolonged antifungal therapy. BMC Infect. Dis. 2015, 15, 340. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Suh, J.W.; Kim, J.Y.; Lee, H.; Kim, S.B.; Sohn, J.W.; Kim, M.J. 1698. Epidemiology and Antifungal Susceptibility of Candidemia Among Adult Patients at a Tertiary Care Hospital in South Korea During an 8-Year Period. Open Forum Infect. Dis. 2019, 6, S621–S622. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Najafzadeh, M.J.; Hagen, F.; Mahmoudi, S.; Salehi, M.; Zarrinfar, H.; Namvar, Z.; Zareshahrabadi, Z.; Khodavaisy, S.; et al. Evaluation of Molecular Epidemiology, Clinical Characteristics, Antifungal Susceptibility Profiles, and Molecular Mechanisms of Antifungal Resistance of Iranian Candida parapsilosis Species Complex Blood Isolates. Front. Cell. Infect. Microbiol. 2020, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Daneshnia, F.; Hilmioglu-Polat, S.; Ilkit, M.; Yasar, M.; Polat, F.; Metin, D.Y.; Dokumcu, Ü.Z.; Pan, W.; Hagen, F.; et al. Genetically related micafungin-resistant Candida parapsilosis blood isolates harbouring novel mutation R658G in hotspot 1 of Fks1p: a new challenge? J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.F.; Ilkit, M.; Boekhout, T.; Gabaldón, T.; Perlin, D. The Quiet and Underappreciated Rise of Drug-Resistant Invasive Fungal Pathogens. J. Fungi 2020, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.J.; Zhao, M.; Andes, D.R. Pharmacodynamic Evaluation of Rezafungin (CD101) against Candida auris in the Neutropenic Mouse Invasive Candidiasis Model. Antimicrob. Agents Chemother. 2018, 62, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, D.A.; Koh, M.; Ostermann, M.; Cockwell, P. Reminder of important clinical lesson Optimal management of acute kidney injury in critically ill patients with invasive fungal infections being treated with liposomal amphotericin B. Br. Med. J. 2020, 13, 1–5. [Google Scholar]
- Almirante, B.; Rodríguez, D.; Rodríguez-Pardo, D. Antifungal Agents in Neonates. Pediatr. Drugs 2007, 9, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-S.; Yoon, Y.; Kwak, S.J.; Kim, Y.-J. Comparison of Candidemia in Patients in Neonatal Intensive Care Unit and Pediatric Patients and Big Data Analysis on Candidiasis and Candidemia in Korean Children. Open Forum Infect. Dis. 2019, 6, S621. [Google Scholar] [CrossRef] [Green Version]
- Jemel, S.; Guillot, J.; Kallel, K.; Botterel, F.; Dannaoui, E. Galleria mellonella for the Evaluation of Antifungal E ffi cacy against Medically Important Fungi, a Narrative Review. Microorganisms 2020, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Daneshnia, F.F.; Salehi, M.; Yasar, M.; Hoşbul, T.; Ilkit, M.; Pan, W.; Hagen, F.; Arslan, N.; Türk-Dağı, H.; et al. Low level of antifungal resistance of Candida glabrata blood isolates in Turkey: Fluconazole minimum inhibitory concentration and FKS mutations can predict therapeutic failure. Mycoses 2020, 63, 911–920. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard, 3rd ed.; M27-A3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Fourth Informational Supplement; CLSI document M27-S4; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morio, F.; Lombardi, L.; Binder, U.; Loge, C.; Robert, E.; Graessle, D.; Bodin, M.; Lass-Flörl, C.; Butler, G.; Le Pape, P. Precise genome editing using a CRISPR-Cas9 method highlights the role of CoERG11 amino acid substitutions in azole resistance in Candida orthopsilosis. J. Antimicrob. Chemother. 2019, 74, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of Clinically Derived Mutations inERG11to Azole Resistance in Candida albicans. Antimicrob. Agents Chemother. 2014, 59, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Forastiero, A.; Bernal-Martínez, L.; Mellado, E.; Cendejas, E.; Gomez-Lopez, A. In vivo efficacy of voriconazole and posaconazole therapy in a novel invertebrate model of Aspergillus fumigatus infection. Int. J. Antimicrob. Agents 2015, 46, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.B.; Li, Y.; Li, D.; Johnston, T.; Hendricks, G.L.; Li, G.; Rajamuthiah, R.; Mylonakis, E. Micafungin Elicits an Immunomodulatory Effect in Galleria mellonella and Mice. Mycopathologia 2015, 181, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Aigner, M.; Risslegger, B.; Hörtnagl, C.; Lass-Flörl, C.; Lackner, M. Minimal Inhibitory Concentration (MIC)-Phenomena in Candida albicans and Their Impact on the Diagnosis of Antifungal Resistance. J. Fungi 2019, 5, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatzer, M.; Blum, G.; Jukic, E.; Posch, W.; Gruber, P.; Nagl, M.; Binder, U.; Maurer, E.; Sarg, B.; Lindner, H.; et al. Blocking Hsp70 Enhances the Efficiency of Amphotericin B Treatment against Resistant Aspergillus terreus Strains. Antimicrob. Agents Chemother. 2015, 59, 3778–3788. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Strain | Resistance Phenotype | Age/Sex | Underlying Conditions | AP | AT | Outcome | MIC (μg/mL) | AA Exchanges in Hot Spot Regions | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FLC | VRC | MFG | AMB | Fks1-HS1 | Erg11 | |||||||
| CP30 | FLCR, VRCR | 16 Y/M | Aplastic anemia | VRC | VRC | Died | >32 | 1 | 0.5 | 0.25 # | WT | Y132F + K143R |
| CP37 | FLCR | 27 Y/F | Cardiomypathy | FLC | FLC | Died | >32 | 0.125 | 1 | 0.5 | WT | Y132F |
| CP 179 | FLCR, VRCR | 3 M/F | Bowel perforation | None | MFG | Died | >32 | 2 | 0.5 | 0.5 | WT | Y132F + G307A |
| L48 | FLCR, VRCR | 14 Y/F | Epileptic seizure and malnutrition | None | AMB and FLC | Survived | >64 | 4 | 1 | 1 | WT | G307A + G458S |
| CP70 | FLCR | 64 Y/M | Cardiovascular complications | None | CSP | Died | >32 | 0.125 | 1 | 0.5 | WT | WT |
| CP207 | MDR | 2 Y/M | Pneumonia, empyema | None | FLC | Survived | >32 | 0.5 | >8 | 0.5 | R658G | Y132F + K143R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, U.; Arastehfar, A.; Schnegg, L.; Hörtnagl, C.; Hilmioğlu-Polat, S.; Perlin, D.S.; Lass-Flörl, C. Efficacy of LAMB against Emerging Azole- and Multidrug-Resistant Candida parapsilosis Isolates in the Galleria mellonella Model. J. Fungi 2020, 6, 377. https://doi.org/10.3390/jof6040377
Binder U, Arastehfar A, Schnegg L, Hörtnagl C, Hilmioğlu-Polat S, Perlin DS, Lass-Flörl C. Efficacy of LAMB against Emerging Azole- and Multidrug-Resistant Candida parapsilosis Isolates in the Galleria mellonella Model. Journal of Fungi. 2020; 6(4):377. https://doi.org/10.3390/jof6040377
Chicago/Turabian StyleBinder, Ulrike, Amir Arastehfar, Lisa Schnegg, Caroline Hörtnagl, Suleyha Hilmioğlu-Polat, David S. Perlin, and Cornelia Lass-Flörl. 2020. "Efficacy of LAMB against Emerging Azole- and Multidrug-Resistant Candida parapsilosis Isolates in the Galleria mellonella Model" Journal of Fungi 6, no. 4: 377. https://doi.org/10.3390/jof6040377
APA StyleBinder, U., Arastehfar, A., Schnegg, L., Hörtnagl, C., Hilmioğlu-Polat, S., Perlin, D. S., & Lass-Flörl, C. (2020). Efficacy of LAMB against Emerging Azole- and Multidrug-Resistant Candida parapsilosis Isolates in the Galleria mellonella Model. Journal of Fungi, 6(4), 377. https://doi.org/10.3390/jof6040377