Polymorphisms within the TNFSF4 and MAPKAPK2 Loci Influence the Risk of Developing Invasive Aspergillosis: A Two-Stage Case Control Study in the Context of the aspBIOmics Consortium

 ,
,  , ,
, ,  , ,
, ,  ,
,  , , ,
, , ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Methods
2.1. Discovery and Replication Populations
2.2. DNA Extraction, SNP Selection, Genotyping, and Quality Control
2.3. Hardy–Weinberg Equilibrium, Genetic Association Analysis, and Meta-Analysis
2.4. Linkage Disequilibrium (LD) and Haplotype Analysis
2.5. Cell Isolation and Differentiation and Functional Analysis of the TNFSF4 and MAPKAPK2 Variants
2.6. Correlation between TNFSF4 and MAPKAPK2 Polymorphisms and Cell Counts of 91 Blood-Derived Immune Cell Populations and Serum/Plasmatic Proteomic Profile
2.7. Assessment of Fungicidal Activity
2.8. In Silico Functional Analysis
3. Results
3.1. Characteristics of Study Subjects
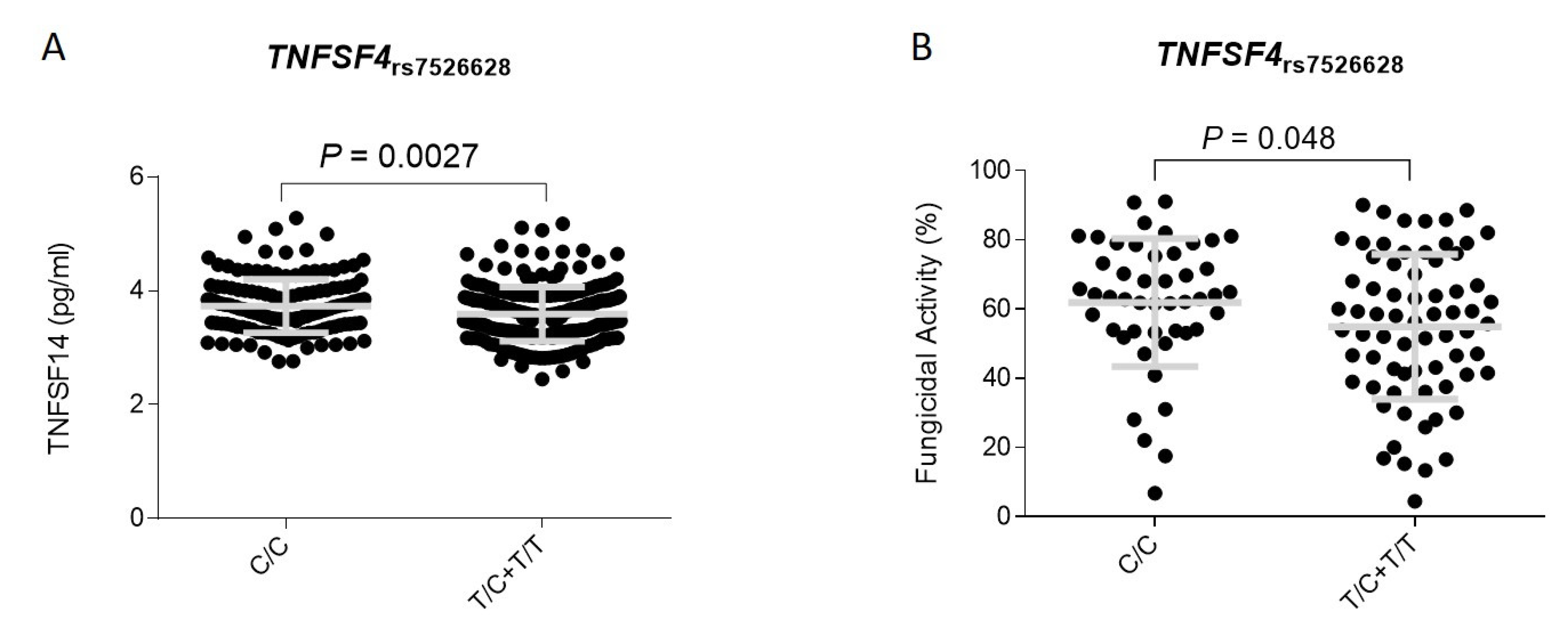
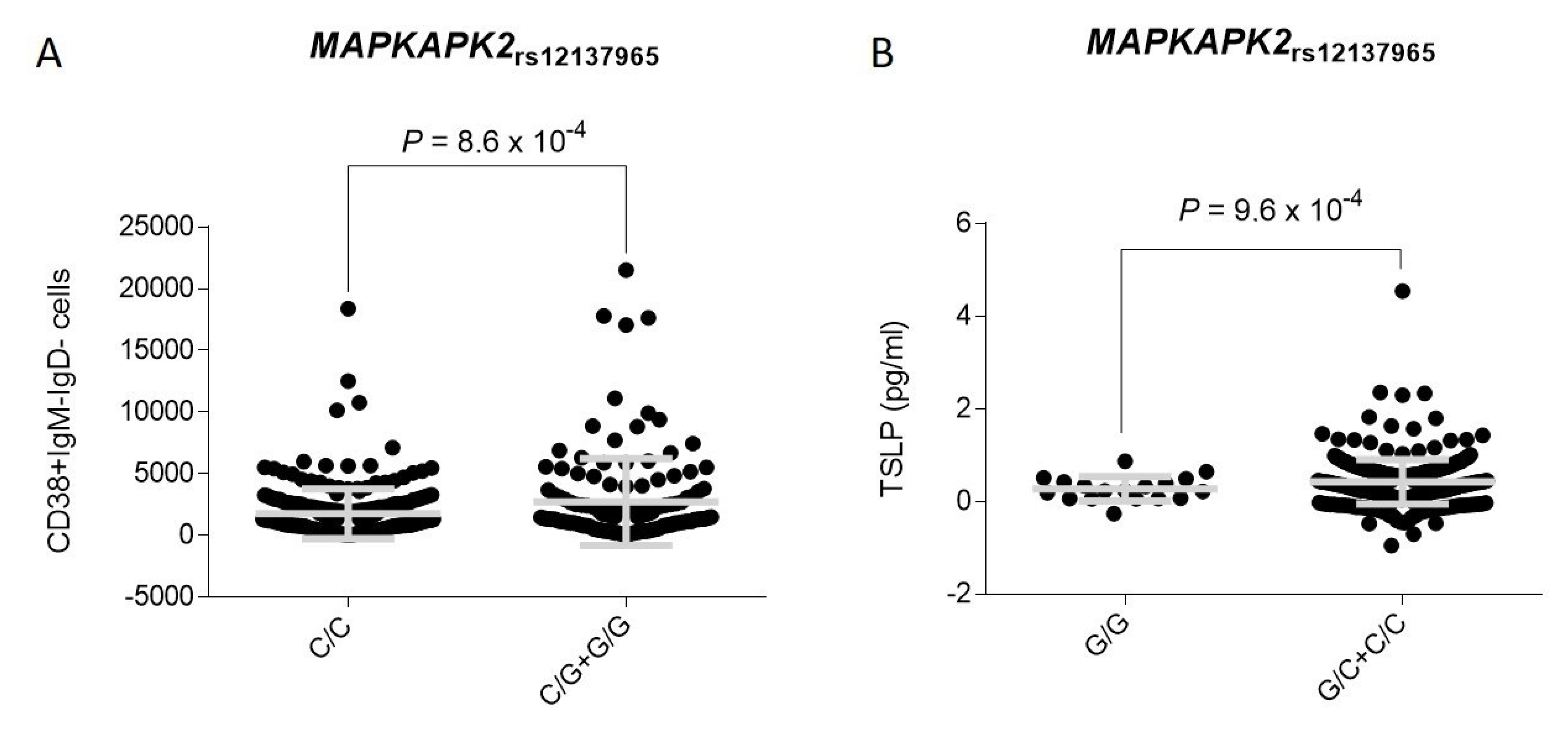
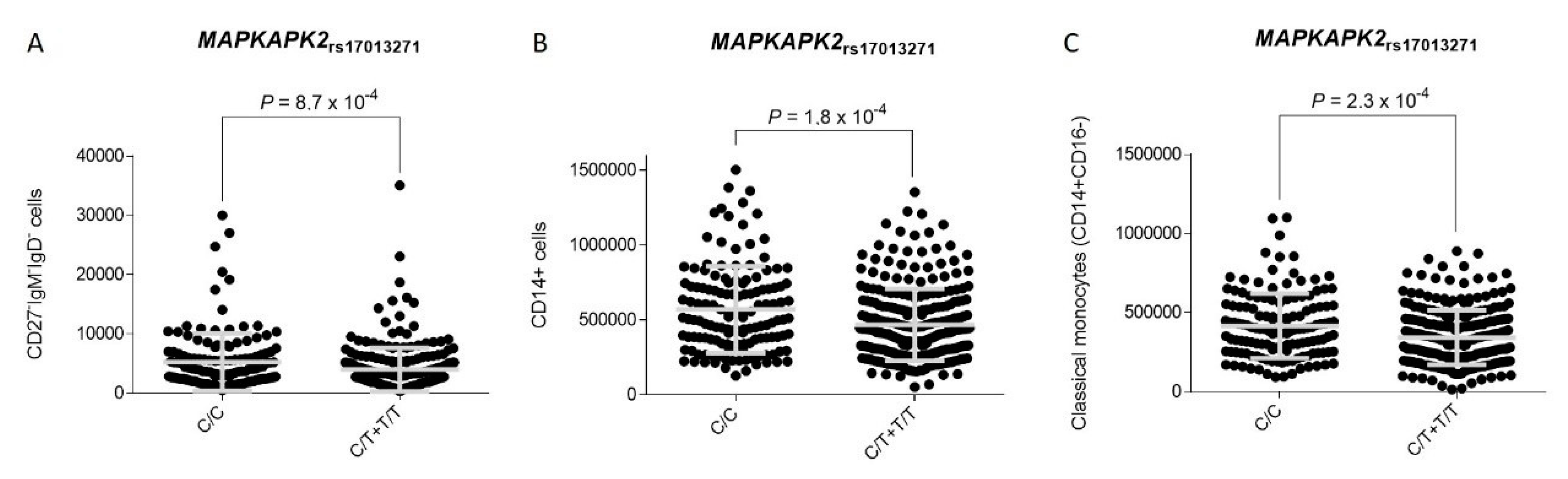
3.2. Association Analysis and Functional Impact of TNFSF4 and MAPKAPK2 Polymorphisms


{kind=link}
{kind=link}
{kind=link}
| SNP_rsID | Gene | Risk Allele | OR (95% CI) δ | p | OR (95% CI) Ϯ | p | OR (95% CI) ¥ | p |
|---|---|---|---|---|---|---|---|---|
| rs10489269 | TNFSF4 | A | 1.23 (0.56–2.70) | 0.61 | 1.08 (0.47–2.51) | 0.86 | NA (NA-NA) | NA |
| rs7518045 | TNFSF4 | G | 1.08 (0.54–2.15) | 0.83 | 0.91 (0.41–1.98) | 0.80 | NA (NA-NA) | NA |
| rs10489266 | TNFSF4 | G | 0.56 (0.27–1.17) | 0.10 | 0.58 (0.27–1.26) | 0.15 | NA (NA-NA) | NA |
| rs2205959 | TNFSF4 | G | 0.85 (0.57–1.27) | 0.44 | 1.11 (0.61–2.00) | 0.73 | 0.46 (0.20–1.09) | 0.058 |
| rs61828280 | TNFSF4 | G | 0.63 (0.30–1.32) | 0.20 | 0.66 (0.30–1.42) | 0.27 | NA (NA-NA) | NA |
| rs4916320 | TNFSF4 | G | 0.76 (0.40–1.44) | 0.38 | 0.77 (0.40–1.49) | 0.43 | NA (NA-NA) | NA |
| rs4357565 | TNFSF4 | C | 1.73 (1.13–2.63) | 0.011 | 2.13 (1.21–3.74) | 0.0077 | 1.69 (0.67–4.24) | 0.28 |
| rs1342038 | TNFSF4 | A | 0.68 (0.44–1.03) | 0.065 | 0.73 (0.41–1.30) | 0.29 | 0.40 (0.16–1.02) | 0.056 |
| rs947505 | TNFSF4 | A | 1.17 (0.68–2.01) | 0.58 | 1.07 (0.60–1.92) | 0.82 | 4.17 (0.62–28.3) | 0.17 |
| rs56307807 | TNFSF4 | C | 0.95 (0.45–2.02) | 0.90 | 0.85 (0.38–1.94) | 0.70 | 4.32 (0.26–72.4) | 0.33 |
| rs6425219 | TNFSF4 | T | 1.08 (0.72–1.63) | 0.71 | 1.07 (0.62–1.87) | 0.80 | 1.19 (0.51–2.76) | 0.69 |
| rs1418191 | TNFSF4 | C | 1.47 (0.85–2.54) | 0.18 | 1.63 (0.88–3.00) | 0.13 | 0.87 (0.09–8.34) | 0.90 |
| rs1578624 | TNFSF4 | C | 0.86 (0.58–1.27) | 0.44 | 0.73 (0.41–1.29) | 0.29 | 0.97 (0.49–1.92) | 0.92 |
| rs7526628 | TNFSF4 | T | 1.72 (1.16–2.55) | 0.0072 | 1.87 (1.02–3.42) | 0.038 | 2.27 (1.22–4.20) | 0.0068 |
| rs7549074 | TNFSF4 | C | 1.43 (0.90–2.26) | 0.14 | 1.34 (0.77–2.31) | 0.30 | 2.95 (0.90–9.66) | 0.089 |
| rs6700269 | TNFSF4 | T | 1.74 (0.97–3.12) | 0.075 | 1.94 (0.9–3.77) | 0.059 | 1.56 (0.17–14.2) | 0.71 |
| rs12140760 | TNFSF4 | A | 0.83 (0.56–1.21) | 0.33 | 0.79 (0.45–1.37) | 0.40 | 0.75 (0.35–1.58) | 0.44 |
| rs17013271 | DYRK3 | T | 0.48 (0.32–0.73) | 0.0004 | 0.48 (0.28–0.83) | 0.0087 | 0.18 (0.05–0.59) | 0.0005 |
| rs6540512 | DYRK3 | A | 1.29 (078–2.11) | 0.32 | 1.47 (0.83–2.58) | 0.19 | 0.52 (0.06–4.37) | 0.51 |
| rs12126682 | DYRK3 | G | 1.47 (1.01–2.14) | 0.044 | 1.62 (0.89–2.97) | 0.11 | 1.77 (0.94–3.34) | 0.084 |
| rs4845123 | DYRK3 | A | 0.59 (0.38–0.90) | 0.012 | 0.60 (0.35–1.03) | 0.063 | 0.27 (0.08–0.89) | 0.012 |
| rs17435120 | DYRK3|MAPKAPK2 | G | 1.42 (0.98–2.06) | 0.065 | 1.44 (0.80–2.58) | 0.21 | 1.85 (0.98–3.49) | 0.065 |
| rs12407425 | DYRK3|MAPKAPK2 | A | 0.85 (0.46–1.58) | 0.61 | 0.81 (0.41–1.59) | 0.53 | 1.29 (0.14–11.8) | 0.82 |
| rs11119267 | DYRK3|MAPKAPK2 | C | 1.49 (1.00–2.22) | 0.047 | 2.40 (1.26–4.59) | 0.0051 | 1.07 (0.51–2.25) | 0.86 |
| rs12038489 | DYRK3|MAPKAPK2 | A | 1.03 (0.58–1.81) | 0.92 | 1.08 (0.57–2.04) | 0.82 | 0.67 (0.07–6.02) | 0.71 |
| rs61815610 | MAPKAPK2 | T | 0.79 (0.41–1.52) | 0.47 | 0.82 (0.42–1.62) | 0.56 | NA (NA-NA) | NA |
| rs10863788 | MAPKAPK2 | A | 0.73 (0.49–1.09) | 0.12 | 0.83 (0.48–1.44) | 0.51 | 0.37 (0.14–0.98) | 0.027 |
| rs12123706 | MAPKAPK2 | T | 1.74 (1.12–2.70) | 0.014 | 2.35 (1.35–4.10) | 0.0022 | 0.86 (0.24–3.12) | 0.81 |
| rs4072677 | MAPKAPK2 | G | 0.85 (0.58–1.24) | 0.39 | 0.92 (0.52–1.63) | 0.77 | 0.65 (0.31–1.35) | 0.23 |
| rs61815626 | MAPKAPK2 | A | 0.35 (0.10–1.17) | 0.049 | 0.34 (0.10–1.18) | 0.054 | NA (NA-NA) | NA |
| rs12030124 | MAPKAPK2 | T | 1.21 (0.72–2.03) | 0.48 | 1.29 (0.72–2.28) | 0.40 | 0. (0.09–6.73) | 0.81 |
| rs4256810 | MAPKAPK2 | C | 0.72 (0.46–1.13) | 0.14 | 0.75 (0.44–1.29) | 0.29 | 0.36 (0.08–1.56) | 0.76 |
| rs28394820 | MAPKAPK2 | C | 0.80 (0.47–1.35) | 0.39 | 0.88 (0.50–1.55) | 0.65 | NA (NA-NA) | NA |
| rs4073250 | MAPKAPK2 | T | 0.69 (0.43–1.11) | 0.12 | 0.70 (0.39–1.25) | 0.22 | 0.35 (0.08–1.55) | 0.11 |
| rs7515374 | MAPKAPK2||IL10 | C | 0.86 (0.58–1.28) | 0.46 | 0.88 (0.50–1.54) | 0.66 | 0.71 (0.58–1.28) | 0.46 |
| rs12137965 | MAPKAPK2||IL10 | G | 2.06 (1.34–3.16) | 0.0011 | 3.01 (1.68–5.38) | 0.0001 | 1.38 (0.48–3.95) | 0.56 |
| Discovery Population (n = 423) | Replication Population (n = 488) | Meta-Analysis (n = 911) | ||||||
|---|---|---|---|---|---|---|---|---|
| SNP_rsID | Risk Allele | OR (95% CI) δ | p | OR (95% CI) δ | p | OR (95% CI) δ | p | PHet |
| rs4357565 | C | 1.73 (1.13–2.63) | 0.011 | 0.95 (0.64–1.42) | 0.81 | 1.26 (0.94–1.68) | 0.118 | 0.043 |
| rs4357565 | C | 1.69 (0.67–4.24) ¥ | 0.28 | 1.31 (0.55–3.11) ¥ | 0.55 | 1.48 (0.79–2.78) ¥ | 0.227 | 0.693 |
| rs7526628 | T | 1.72 (1.16–2.55) | 0.0072 | 1.38 (0.98–1.94) | 0.068 | 1.52 (1.17–1.96) | 0.0015 | 0.41 |
| rs7526628 | T | 2.27 (1.22–4.20) ¥ | 0.0068 | 2.23 (1.22–4.06) ¥ | 0.011 | 2.25 (1.46–3.46) ¥ | 0.00022 | 0.97 |
| rs17013271 | T | 0.48 (0.32–0.73) | 0.0004 | 0.85 (0.58–1.23) | 0.38 | 0.66 (0.50–0.87) | 0.0029 | 0.05 |
| rs17013271 | T | 0.48 (0.28–0.83) Ϯ | 0.0087 | 0.73 (0.44–1.23) Ϯ | 0.24 | 0.60 (0.41–0.87) Ϯ | 0.0071 | 0.27 |
| rs12126682 | G | 1.47 (1.01–2.14) | 0.044 | 0.82 (0.56–1.19) | 0.29 | 1.10 (0.84–1.43) | 0.486 | 0.032 |
| rs12126682 | G | 1.62 (0.89–2.97) Ϯ | 0.11 | 0.81 (0.48–1.34) Ϯ | 0.41 | 1.08 (0.73–1.60) | 0.685 | 0.086 |
| rs11119267 | C | 1.49 (1.00–2.22) | 0.047 | 1.40 (0.97–2.02) | 0.069 | 1.44 (1.10–1.89) | 0.0080 | 0.822 |
| rs11119267 | C | 2.40 (1.26–4.59) Ϯ | 0.0051 | 1.55 (0.86–2.78) Ϯ | 0.13 | 1.89 (1.22–2.92) | 0.0040 | 0.326 |
| rs12123706 | T | 1.74 (1.12–2.70) | 0.014 | 0.84 (0.55–1.29) | 0.43 | 1.20 (0.88–1.62) | 0.254 | 0.020 |
| rs12123706 | T | 2.35 (1.35–4.10) Ϯ | 0.0022 | 0.88 (0.52–1.48) Ϯ | 0.63 | 1.40 (0.95–2.04) | 0.086 | 0.012 |
| rs12137965 | G | 2.06 (1.34–3.16) | 0.0011 | 1.28 (0.85–1.91) | 0.24 | 1.60 (1.19–2.15) | 0.0017 | 0.11 |
| rs12137965 | G | 3.01 (1.68–5.38) Ϯ | 0.0001 | 1.26 (0.77–2.08) Ϯ | 0.36 | 1.81 (1.25–2.65) Ϯ | 0.0019 | 0.030 |
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baddley, J.W.; Andes, D.R.; Marr, K.A.; Kontoyiannis, D.P.; Alexander, B.D.; Kauffman, C.A.; Oster, R.A.; Anaissie, E.J.; Walsh, T.J.; Schuster, M.G.; et al. Factors associated with mortality in transplant patients with invasive aspergillosis. Clin. Infect. Dis. 2010, 50, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Marr, K.A.; Anaissie, E.J.; Azie, N.; Quan, S.P.; Meier-Kriesche, H.U.; Apewokin, S.; Horn, D.L. Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J. Infect. 2012, 65, 453–464. [Google Scholar] [CrossRef]
- Dutkiewicz, R.; Hage, C.A. Aspergillus infections in the critically ill. Proc. Am. Thorac. Soc. 2010, 7, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Maertens, J.A.; Raad, I.I.; Marr, K.A.; Patterson, T.F.; Kontoyiannis, D.P.; Cornely, O.A.; Bow, E.J.; Rahav, G.; Neofytos, D.; Aoun, M.; et al. Isavuconazole versus voriconazole for primary treatment of invasive mould disease caused by Aspergillus and other filamentous fungi (SECURE): A phase 3, randomised-controlled, non-inferiority trial. Lancet 2016, 387, 760–769. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Levitz, S.M. Host Control of Fungal Infections: Lessons from Basic Studies and Human Cohorts. Annu. Rev. Immunol. 2018, 36, 157–191. [Google Scholar] [CrossRef]
- Cunha, C.; Aversa, F.; Romani, L.; Carvalho, A. Human genetic susceptibility to invasive aspergillosis. PLoS Pathog. 2013, 9, e1003434. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.; Aversa, F.; Lacerda, J.F.; Busca, A.; Kurzai, O.; Grube, M.; Loffler, J.; Maertens, J.A.; Bell, A.S.; Inforzato, A.; et al. Genetic PTX3 deficiency and aspergillosis in stem-cell transplantation. N. Engl. J. Med. 2014, 370, 421–432. [Google Scholar] [CrossRef]
- Fisher, C.E.; Hohl, T.M.; Fan, W.; Storer, B.E.; Levine, D.M.; Zhao, L.P.; Martin, P.J.; Warren, E.H.; Boeckh, M.; Hansen, J.A. Validation of single nucleotide polymorphisms in invasive aspergillosis following hematopoietic cell transplantation. Blood 2017, 129, 2693–2701. [Google Scholar] [CrossRef]
- Bochud, P.Y.; Chien, J.W.; Marr, K.A.; Leisenring, W.M.; Upton, A.; Janer, M.; Rodrigues, S.D.; Li, S.; Hansen, J.A.; Zhao, L.P.; et al. Toll-like receptor 4 polymorphisms and aspergillosis in stem-cell transplantation. N. Engl. J. Med. 2008, 359, 1766–1777. [Google Scholar] [CrossRef] [Green Version]
- Sainz, J.; Lupianez, C.B.; Segura-Catena, J.; Vazquez, L.; Rios, R.; Oyonarte, S.; Hemminki, K.; Forsti, A.; Jurado, M. Dectin-1 and DC-SIGN polymorphisms associated with invasive pulmonary Aspergillosis infection. PLoS ONE 2012, 7, e32273. [Google Scholar] [CrossRef]
- Gresnigt, M.S.; Cunha, C.; Jaeger, M.; Goncalves, S.M.; Malireddi, R.K.S.; Ammerdorffer, A.; Lubbers, R.; Oosting, M.; Rasid, O.; Jouvion, G.; et al. Genetic deficiency of NOD2 confers resistance to invasive aspergillosis. Nat. Commun. 2018, 9, 2636. [Google Scholar] [CrossRef] [PubMed]
- Cunha, D.O.; Leao-Cordeiro, J.A.B.; Paula, H.; Ataides, F.S.; Saddi, V.A.; Vilanova-Costa, C.; Silva, A. Association between polymorphisms in the genes encoding toll-like receptors and dectin-1 and susceptibility to invasive aspergillosis: A systematic review. Rev. Soc. Bras. Med. Trop. 2018, 51, 725–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, C.; Di Ianni, M.; Bozza, S.; Giovannini, G.; Zagarella, S.; Zelante, T.; D’Angelo, C.; Pierini, A.; Pitzurra, L.; Falzetti, F.; et al. Dectin-1 Y238X polymorphism associates with susceptibility to invasive aspergillosis in hematopoietic transplantation through impairment of both recipient- and donor-dependent mechanisms of antifungal immunity. Blood 2010, 116, 5394–5402. [Google Scholar] [CrossRef] [PubMed]
- Stappers, M.H.T.; Clark, A.E.; Aimanianda, V.; Bidula, S.; Reid, D.M.; Asamaphan, P.; Hardison, S.E.; Dambuza, I.M.; Valsecchi, I.; Kerscher, B.; et al. Recognition of DHN-melanin by a C-type lectin receptor is required for immunity to Aspergillus. Nature 2018, 555, 382–386. [Google Scholar] [CrossRef]
- Sainz, J.; Perez, E.; Gomez-Lopera, S.; Jurado, M. IL1 gene cluster polymorphisms and its haplotypes may predict the risk to develop invasive pulmonary aspergillosis and modulate C-reactive protein level. J. Clin. Immunol. 2008, 28, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Sainz, J.; Perez, E.; Gomez-Lopera, S.; Lopez-Fernandez, E.; Moratalla, L.; Oyonarte, S.; Jurado, M. Genetic variants of IL6 gene promoter influence on C-reactive protein levels but are not associated with susceptibility to invasive pulmonary aspergillosis in haematological patients. Cytokine 2008, 41, 268–278. [Google Scholar] [CrossRef]
- Cunha, C.; Goncalves, S.M.; Duarte-Oliveira, C.; Leite, L.; Lagrou, K.; Marques, A.; Lupianez, C.B.; Mesquita, I.; Gaifem, J.; Barbosa, A.M.; et al. IL-10 overexpression predisposes to invasive aspergillosis by suppressing antifungal immunity. J. Allergy Clin. Immunol. 2017, 140, 867–870.e9. [Google Scholar] [CrossRef] [Green Version]
- Mezger, M.; Steffens, M.; Beyer, M.; Manger, C.; Eberle, J.; Toliat, M.R.; Wienker, T.F.; Ljungman, P.; Hebart, H.; Dornbusch, H.J.; et al. Polymorphisms in the chemokine (C-X-C motif) ligand 10 are associated with invasive aspergillosis after allogeneic stem-cell transplantation and influence CXCL10 expression in monocyte-derived dendritic cells. Blood 2008, 111, 534–536. [Google Scholar] [CrossRef]
- Cunha, C.; Carvalho, A. Genetic defects in fungal recognition and susceptibility to invasive pulmonary aspergillosis. Med. Mycol. 2019, 57, S211–S218. [Google Scholar] [CrossRef]
- Netea, M.G.; Wijmenga, C.; O’Neill, L.A. Genetic variation in Toll-like receptors and disease susceptibility. Nat. Immunol. 2012, 13, 535–542. [Google Scholar] [CrossRef]
- Wojtowicz, A.; Bochud, P.Y. Host genetics of invasive Aspergillus and Candida infections. Semin. Immunopathol. 2015, 37, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Lupianez, C.B.; Martinez-Bueno, M.; Sanchez-Maldonado, J.M.; Badiola, J.; Cunha, C.; Springer, J.; Lackner, M.; Segura-Catena, J.; Canet, L.M.; Alcazar-Fuoli, L.; et al. Polymorphisms within the ARNT2 and CX3CR1 Genes Are Associated with the Risk of Developing Invasive Aspergillosis. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef] [PubMed]
- Sainz, J.; Salas-Alvarado, I.; Lopez-Fernandez, E.; Olmedo, C.; Comino, A.; Garcia, F.; Blanco, A.; Gomez-Lopera, S.; Oyonarte, S.; Bueno, P.; et al. TNFR1 mRNA expression level and TNFR1 gene polymorphisms are predictive markers for susceptibility to develop invasive pulmonary aspergillosis. Int. J. Immunopathol. Pharmacol. 2010, 23, 423–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupianez, C.B.; Villaescusa, M.T.; Carvalho, A.; Springer, J.; Lackner, M.; Sanchez-Maldonado, J.M.; Canet, L.M.; Cunha, C.; Segura-Catena, J.; Alcazar-Fuoli, L.; et al. Common Genetic Polymorphisms within NFkappaB-Related Genes and the Risk of Developing Invasive Aspergillosis. Front. Microbiol. 2016, 7, 1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, C.; Giovannini, G.; Pierini, A.; Bell, A.S.; Sorci, G.; Riuzzi, F.; Donato, R.; Rodrigues, F.; Velardi, A.; Aversa, F.; et al. Genetically-determined hyperfunction of the S100B/RAGE axis is a risk factor for aspergillosis in stem cell transplant recipients. PLoS ONE 2011, 6, e27962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolioni, V.; Pariano, M.; Borghi, M.; Oikonomou, V.; Galosi, C.; De Luca, A.; Stincardini, C.; Vacca, C.; Renga, G.; Lucidi, V.; et al. Genetic Polymorphisms Affecting IDO1 or IDO2 Activity Differently Associate With Aspergillosis in Humans. Front. Immunol. 2019, 10, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrios, C.S.; Johnson, B.D.; Henderson, J.D.; Fink, J.N.; Kelly, K.J.; Kurup, V.P. The costimulatory molecules CD80, CD86 and OX40L are up-regulated in Aspergillus fumigatus sensitized mice. Clin. Exp. Immunol. 2005, 142, 242–250. [Google Scholar] [CrossRef]
- Chiang, L.Y.; Sheppard, D.C.; Gravelat, F.N.; Patterson, T.F.; Filler, S.G. Aspergillus fumigatus stimulates leukocyte adhesion molecules and cytokine production by endothelial cells in vitro and during invasive pulmonary disease. Infect. Immun. 2008, 76, 3429–3438. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Chen, C.; Wu, J.; Dai, C.; Wu, X. TSLP-activated dendritic cells induce T helper type 2 inflammation in Aspergillus fumigatus keratitis. Exp. Eye Res. 2018, 171, 120–130. [Google Scholar] [CrossRef]
- Menon, M.B.; Gaestel, M. MK2-TNF-Signaling Comes Full Circle. Trends Biochem. Sci. 2018, 43, 170–179. [Google Scholar] [CrossRef]
- Wu, Y.; He, H.; Ding, Y.; Liu, S.; Zhang, D.; Wang, J.; Jiang, H.; Zhang, D.; Sun, L.; Ye, R.D.; et al. MK2 mediates macrophage activation and acute lung injury by regulating let-7e miRNA. Am. J. Physiol. Lung Cell. Mol. Physiol 2018, 315, L371–L381. [Google Scholar] [CrossRef] [PubMed]
- Ronkina, N.; Shushakova, N.; Tiedje, C.; Yakovleva, T.; Tollenaere, M.A.X.; Scott, A.; Batth, T.S.; Olsen, J.V.; Helmke, A.; Bekker-Jensen, S.H.; et al. The Role of TTP Phosphorylation in the Regulation of Inflammatory Cytokine Production by MK2/3. J. Immunol. 2019, 203, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Lupianez, C.B.; Canet, L.M.; Carvalho, A.; Alcazar-Fuoli, L.; Springer, J.; Lackner, M.; Segura-Catena, J.; Comino, A.; Olmedo, C.; Rios, R.; et al. Polymorphisms in Host Immunity-Modulating Genes and Risk of Invasive Aspergillosis: Results from the AspBIOmics Consortium. Infect. Immun. 2015, 84, 643–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.A.; et al. Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) Consensus Group. Clin. Infect. Dis. 2008, 46, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, S.; Benedict, K.; Derado, G.; Mody, R.K. Trends in Hospitalizations Related to Invasive Aspergillosis and Mucormycosis in the United States, 2000–2013. Open Forum. Infect. Dis. 2017, 4, ofw268. [Google Scholar] [CrossRef] [Green Version]
- Orru, V.; Steri, M.; Sole, G.; Sidore, C.; Virdis, F.; Dei, M.; Lai, S.; Zoledziewska, M.; Busonero, F.; Mulas, A.; et al. Genetic variants regulating immune cell levels in health and disease. Cell 2013, 155, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Gamboa, R.; Joosten, I.; Urbano, P.C.M.; van der Molen, R.G.; van Rijssen, E.; van Cranenbroek, B.; Oosting, M.; Smeekens, S.; Jaeger, M.; Zorro, M.; et al. Differential Effects of Environmental and Genetic Factors on T and B Cell Immune Traits. Cell Rep. 2016, 17, 2474–2487. [Google Scholar] [CrossRef] [Green Version]
- Sugamura, K.; Ishii, N.; Weinberg, A.D. Therapeutic targeting of the effector T-cell co-stimulatory molecule OX40. Nat. Rev. Immunol. 2004, 4, 420–431. [Google Scholar] [CrossRef]
- Murata, K.; Ishii, N.; Takano, H.; Miura, S.; Ndhlovu, L.C.; Nose, M.; Noda, T.; Sugamura, K. Impairment of antigen-presenting cell function in mice lacking expression of OX40 ligand. J. Exp. Med. 2000, 191, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Takeda, I.; Ine, S.; Killeen, N.; Ndhlovu, L.C.; Murata, K.; Satomi, S.; Sugamura, K.; Ishii, N. Distinct roles for the OX40-OX40 ligand interaction in regulatory and nonregulatory T cells. J. Immunol. 2004, 172, 3580–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, S.J.; Perona-Wright, G.; Worsley, A.G.; Ishii, N.; MacDonald, A.S. Dendritic cell expression of OX40 ligand acts as a costimulatory, not polarizing, signal for optimal Th2 priming and memory induction in vivo. J. Immunol. 2007, 179, 3515–3523. [Google Scholar] [CrossRef] [PubMed]
- Salek-Ardakani, S.; Song, J.; Halteman, B.S.; Jember, A.G.; Akiba, H.; Yagita, H.; Croft, M. OX40 (CD134) controls memory T helper 2 cells that drive lung inflammation. J. Exp. Med. 2003, 198, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Oakley, M.S.; Majam, V.; Mahajan, B.; Gerald, N.; Anantharaman, V.; Ward, J.M.; Faucette, L.J.; McCutchan, T.F.; Zheng, H.; Terabe, M.; et al. Pathogenic roles of CD14, galectin-3, and OX40 during experimental cerebral malaria in mice. PLoS ONE 2009, 4, e6793. [Google Scholar] [CrossRef] [Green Version]
- Duffus, K.; Lopez-Isac, E.; Teruel, M.; Simeon, C.P.; Carreria, P.; Ortego-Centeno, N.; Vicente, E.; Worthington, J.; Herrick, A.L.; Martin, J. Association of TNFSF4 (OX40L) polymorphisms with systemic sclerosis-related calcinosis. Rheumatology 2019, 58, 1299–1301. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Tsao, B.P. Genetic susceptibility to systemic lupus erythematosus in the genomic era. Nat. Rev. Rheumatol. 2010, 6, 683–692. [Google Scholar] [CrossRef]
- Jindra, P.T.; Conway, S.E.; Ricklefs, S.M.; Porcella, S.F.; Anzick, S.L.; Haagenson, M.; Wang, T.; Spellman, S.; Milford, E.; Kraft, P.; et al. Analysis of a Genetic Polymorphism in the Costimulatory Molecule TNFSF4 with Hematopoietic Stem Cell Transplant Outcomes. Biol. Blood Marrow Transplant. 2016, 22, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.A.; Soroosh, P.; Khorram, N.; Fukuyama, S.; Rosenthal, P.; Cho, J.Y.; Norris, P.S.; Choi, H.; Scheu, S.; Pfeffer, K.; et al. The tumor necrosis factor family member LIGHT is a target for asthmatic airway remodeling. Nat. Med. 2011, 17, 596–603. [Google Scholar] [CrossRef]
- Herro, R.; Da Silva Antunes, R.; Aguilera, A.R.; Tamada, K.; Croft, M. Tumor necrosis factor superfamily 14 (LIGHT) controls thymic stromal lymphopoietin to drive pulmonary fibrosis. J. Allergy Clin. Immunol. 2015, 136, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.L.; Chen, K.; McAleer, J.; Kolls, J.K. Vitamin D regulation of OX40 ligand in immune responses to Aspergillus fumigatus. Infect. Immun. 2013, 81, 1510–1519. [Google Scholar] [CrossRef] [Green Version]
- Kreindler, J.L.; Steele, C.; Nguyen, N.; Chan, Y.R.; Pilewski, J.M.; Alcorn, J.F.; Vyas, Y.M.; Aujla, S.J.; Finelli, P.; Blanchard, M.; et al. Vitamin D3 attenuates Th2 responses to Aspergillus fumigatus mounted by CD4+ T cells from cystic fibrosis patients with allergic bronchopulmonary aspergillosis. J. Clin. Investig. 2010, 120, 3242–3254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.C.; Ziegler, S.F. Inducible expression of the proallergic cytokine thymic stromal lymphopoietin in airway epithelial cells is controlled by NFkappaB. Proc. Natl. Acad. Sci. USA 2007, 104, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Shan, L.; Rahman, M.S.; Unruh, H.; Halayko, A.J.; Gounni, A.S. Constitutive and inducible thymic stromal lymphopoietin expression in human airway smooth muscle cells: Role in chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L375–L382. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, M.; Rochman, Y.; Spolski, R.; Samsel, L.; Leonard, W.J. Thymic stromal lymphopoietin is produced by dendritic cells. J. Immunol. 2011, 187, 1207–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Cho, K.A.; Kim, J.Y.; Kim, J.Y.; Baek, J.H.; Woo, S.Y.; Kim, J.W. Filaggrin knockdown and Toll-like receptor 3 (TLR3) stimulation enhanced the production of thymic stromal lymphopoietin (TSLP) from epidermal layers. Exp. Dermatol. 2011, 20, 149–151. [Google Scholar] [CrossRef]
- Ni, G.; Chen, Y.; Wu, F.; Zhu, P.; Song, L. NOD2 promotes cell proliferation and inflammatory response by mediating expression of TSLP in human airway smooth muscle cells. Cell Immunol. 2017, 312, 35–41. [Google Scholar] [CrossRef]
- Qiao, J.; Li, A.; Jin, X. TSLP from RSV-stimulated rat airway epithelial cells activates myeloid dendritic cells. Immunol. Cell Biol. 2011, 89, 231–238. [Google Scholar] [CrossRef]
- Kato, A.; Favoreto, S., Jr.; Avila, P.C.; Schleimer, R.P. TLR3- and Th2 cytokine-dependent production of thymic stromal lymphopoietin in human airway epithelial cells. J. Immunol. 2007, 179, 1080–1087. [Google Scholar] [CrossRef]
- Ito, T.; Wang, Y.H.; Duramad, O.; Hori, T.; Delespesse, G.J.; Watanabe, N.; Qin, F.X.; Yao, Z.; Cao, W.; Liu, Y.J. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J. Exp. Med. 2005, 202, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Ito, T.; Wang, Y.H.; Homey, B.; Watanabe, N.; Martin, R.; Barnes, C.J.; McIntyre, B.W.; Gilliet, M.; Kumar, R.; et al. Maintenance and polarization of human TH2 central memory T cells by thymic stromal lymphopoietin-activated dendritic cells. Immunity 2006, 24, 827–838. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Su, H.; Knudsen, G.; Helms, W.; Su, L. Delayed functional maturation of natural regulatory T cells in the medulla of postnatal thymus: Role of TSLP. BMC Immunol. 2006, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, M.J.; Ryan, P.L.; Kirchhof, N.; Panoskaltsis-Mortari, A.; Mortari, F.; Tudor, K.S. Overexpression of murine TSLP impairs lymphopoiesis and myelopoiesis. Blood 2004, 103, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redhu, N.S.; Saleh, A.; Halayko, A.J.; Ali, A.S.; Gounni, A.S. Essential role of NF-kappaB and AP-1 transcription factors in TNF-alpha-induced TSLP expression in human airway smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L479–L485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craxton, A.; Shu, G.; Graves, J.D.; Saklatvala, J.; Krebs, E.G.; Clark, E.A. p38 MAPK is required for CD40-induced gene expression and proliferation in B lymphocytes. J. Immunol. 1998, 161, 3225–3236. [Google Scholar]
- Sutherland, C.L.; Heath, A.W.; Pelech, S.L.; Young, P.R.; Gold, M.R. Differential activation of the ERK, JNK, and p38 mitogen-activated protein kinases by CD40 and the B cell antigen receptor. J. Immunol. 1996, 157, 3381–3390. [Google Scholar]
- Sreekanth, G.P.; Chuncharunee, A.; Sirimontaporn, A.; Panaampon, J.; Noisakran, S.; Yenchitsomanus, P.T.; Limjindaporn, T. SB203580 Modulates p38 MAPK Signaling and Dengue Virus-Induced Liver Injury by Reducing MAPKAPK2, HSP27, and ATF2 Phosphorylation. PLoS ONE 2016, 11, e0149486. [Google Scholar] [CrossRef]
- Pan, W.; Zhu, S.; Dai, D.; Liu, Z.; Li, D.; Li, B.; Gagliani, N.; Zheng, Y.; Tang, Y.; Weirauch, M.T.; et al. MiR-125a targets effector programs to stabilize Treg-mediated immune homeostasis. Nat. Commun. 2015, 6, 7096. [Google Scholar] [CrossRef]
- Swertz, M.A.; Dijkstra, M.; Adamusiak, T.; van der Velde, J.K.; Kanterakis, A.; Roos, E.T.; Lops, J.; Thorisson, G.A.; Arends, D.; Byelas, G.; et al. The MOLGENIS toolkit: Rapid prototyping of biosoftware at the push of a button. BMC Bioinform. 2010. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.W.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016. [Google Scholar] [CrossRef] [Green Version]


| Discovery Population | ||||
|---|---|---|---|---|
| Overall (n = 423) | IA patients (n = 67) | non-IA patients (n = 356) | p value | |
| Demographic variables | ||||
| Age (Average ± SD) | 52.35 ± 15.37 | 53.00 ± 12.69 | 52.23 ± 15.82 | 0.663 |
| Sex ratio (male/female) | 1.11 (222/200) | 1.79 (43/24) | 1.02 (179/176) | 0.037 |
| Hematological disease | ||||
| AML | 328 (77.54) | 49 (73.13) | 279 (78.37) | 0.346 |
| ALL | 58 (13.71) | 14 (20.90) | 44 (12.36) | 0.062 |
| other | 37 (08.75) | 4 (05.97) | 33 (09.27) | 0.380 |
| allo-SCT | 137 (32.39) | 20 (29.85) | 117 (32.87) | 0.629 |
| Ever received prophylaxis † | 244 (64.89) | 22 (43.13) | 222 (68.31) | 0.0004 |
| Replication population | ||||
| Overall (n = 488) | IA patients (n = 97) | non-IA patients (n = 391) | p value | |
| Demographic variables | ||||
| Age (Average ± SD) | 50.90 ± 17.12 | 51.42 ± 15.72 | 50.76 ± 17.44 | 0.734 |
| Sex ratio (male/female) | 1.32 (278/210) | 2.13 (66/31) | 1.18 (212/179) | 0.014 |
| Hematological disease | ||||
| AML | 317 (64.96) | 63 (65.95) | 254 (64.96) | 0.998 |
| ALL | 55 (11.27) | 10 (10.31) | 45 (11.51) | 0.738 |
| other | 116 (23.77) | 24 (24.74) | 92 (23.53) | 0.802 |
| allo-SCT | 251 (51.43) | 54 (55.67) | 197 (50.38) | 0.351 |
| Ever received prophylaxis † | 193 (64.12) | 41 (62.12) | 152 (64.68) | 0.702 |
| Overall population | ||||
| Overall (n = 911) | IA patients (n = 164) | non-IA patients (n = 747) | p value | |
| Demographic variables | ||||
| Age (Average ± SD) | 51.57 ± 16.35 | 52.07 ± 14.58 | 51.46 ± 16.71 | 0.637 |
| Sex ratio (male/female) | 1.22 (500/410) | 1.98 (109/55) | 1.10 (391/355) | 0.001 |
| Hematological disease | ||||
| AML | 645 (70.80) | 112 (68.29) | 533 (71.35) | 0.435 |
| ALL | 113 (12.40) | 24 (14.63) | 89 (11.91) | 0.339 |
| other | 153 (16.80) | 28 (17.07) | 125 (16.74) | 0.916 |
| allo-SCT | 388 (42.59) | 74 (45.12) | 314 (42.03) | 0.469 |
| Ever received prophylaxis † | 437 (64.55) | 63 (53.85) | 374 (66.79) | 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Maldonado, J.M.; Moñiz-Díez, A.; ter Horst, R.; Campa, D.; Cabrera-Serrano, A.J.; Martínez-Bueno, M.; Garrido-Collado, M.d.P.; Hernández-Mohedo, F.; Fernández-Puerta, L.; López-Nevot, M.Á.; et al. Polymorphisms within the TNFSF4 and MAPKAPK2 Loci Influence the Risk of Developing Invasive Aspergillosis: A Two-Stage Case Control Study in the Context of the aspBIOmics Consortium. J. Fungi 2021, 7, 4. https://doi.org/10.3390/jof7010004
Sánchez-Maldonado JM, Moñiz-Díez A, ter Horst R, Campa D, Cabrera-Serrano AJ, Martínez-Bueno M, Garrido-Collado MdP, Hernández-Mohedo F, Fernández-Puerta L, López-Nevot MÁ, et al. Polymorphisms within the TNFSF4 and MAPKAPK2 Loci Influence the Risk of Developing Invasive Aspergillosis: A Two-Stage Case Control Study in the Context of the aspBIOmics Consortium. Journal of Fungi. 2021; 7(1):4. https://doi.org/10.3390/jof7010004
Chicago/Turabian StyleSánchez-Maldonado, Jose Manuel, Ana Moñiz-Díez, Rob ter Horst, Daniele Campa, Antonio José Cabrera-Serrano, Manuel Martínez-Bueno, María del Pilar Garrido-Collado, Francisca Hernández-Mohedo, Laura Fernández-Puerta, Miguel Ángel López-Nevot, and et al. 2021. "Polymorphisms within the TNFSF4 and MAPKAPK2 Loci Influence the Risk of Developing Invasive Aspergillosis: A Two-Stage Case Control Study in the Context of the aspBIOmics Consortium" Journal of Fungi 7, no. 1: 4. https://doi.org/10.3390/jof7010004
APA StyleSánchez-Maldonado, J. M., Moñiz-Díez, A., ter Horst, R., Campa, D., Cabrera-Serrano, A. J., Martínez-Bueno, M., Garrido-Collado, M. d. P., Hernández-Mohedo, F., Fernández-Puerta, L., López-Nevot, M. Á., Cunha, C., González-Sierra, P. A., Springer, J., Lackner, M., Alcazar-Fuoli, L., Fianchi, L., Aguado, J. M., Pagano, L., López-Fernández, E., ... Sainz, J. (2021). Polymorphisms within the TNFSF4 and MAPKAPK2 Loci Influence the Risk of Developing Invasive Aspergillosis: A Two-Stage Case Control Study in the Context of the aspBIOmics Consortium. Journal of Fungi, 7(1), 4. https://doi.org/10.3390/jof7010004