Paracoccidioides brasiliensis Isolated from Nine-Banded Armadillos (Dasypus novemcinctus) Reveal Population Structure and Admixture in the Amazon Basin

 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Armadillos Capture, Euthanasia, and Fungal Isolation
2.2. DNA Extraction, Library Preparation and Genome Sequencing
2.3. Read Mapping and Single Nucleotide Polymorphism (SNP) Calling
2.4. Phylogenetic Analysis
2.5. Admixture Analyses
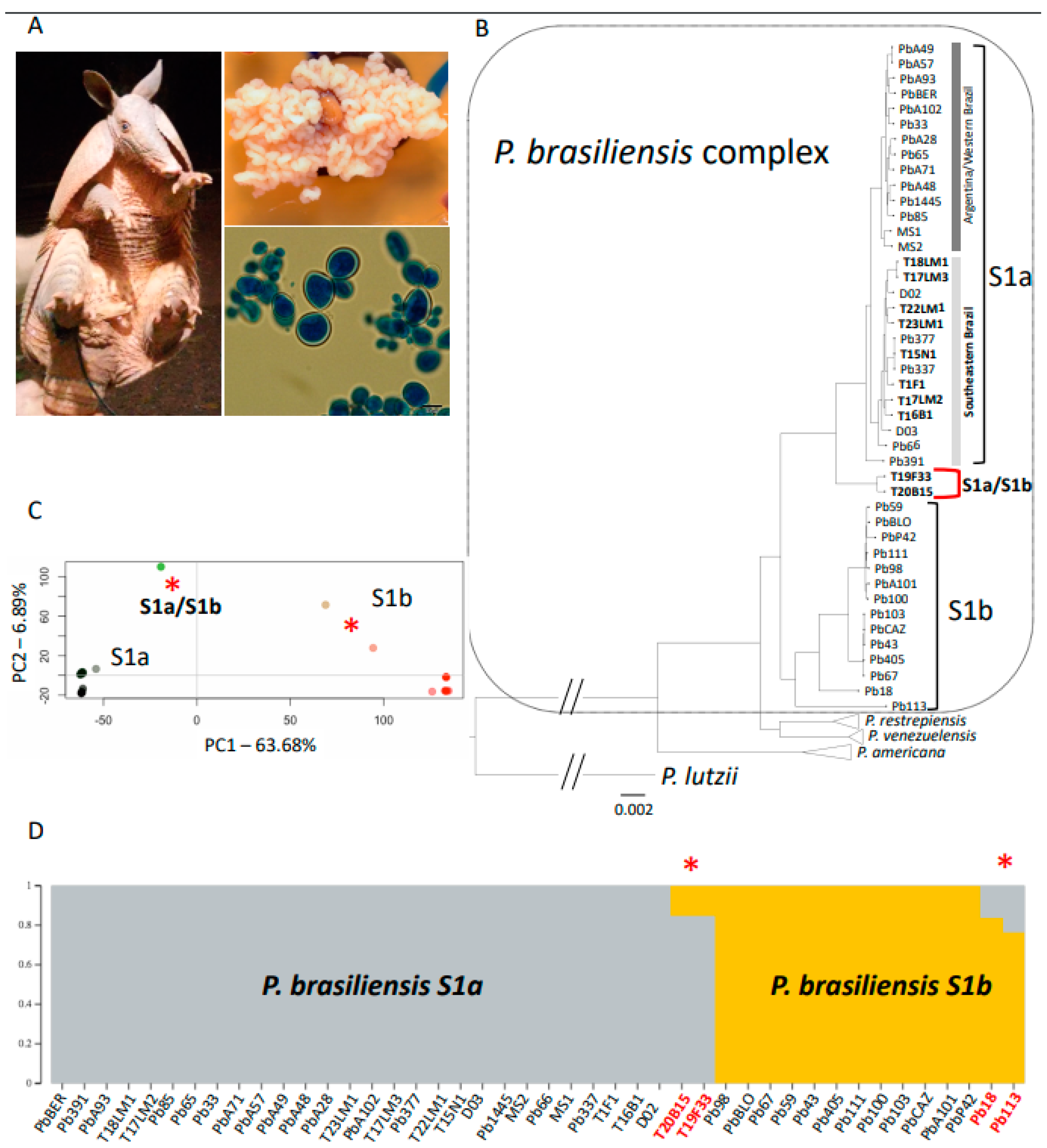
3. Results
3.1. Genomic Data
3.2. Phylogenetic Tree
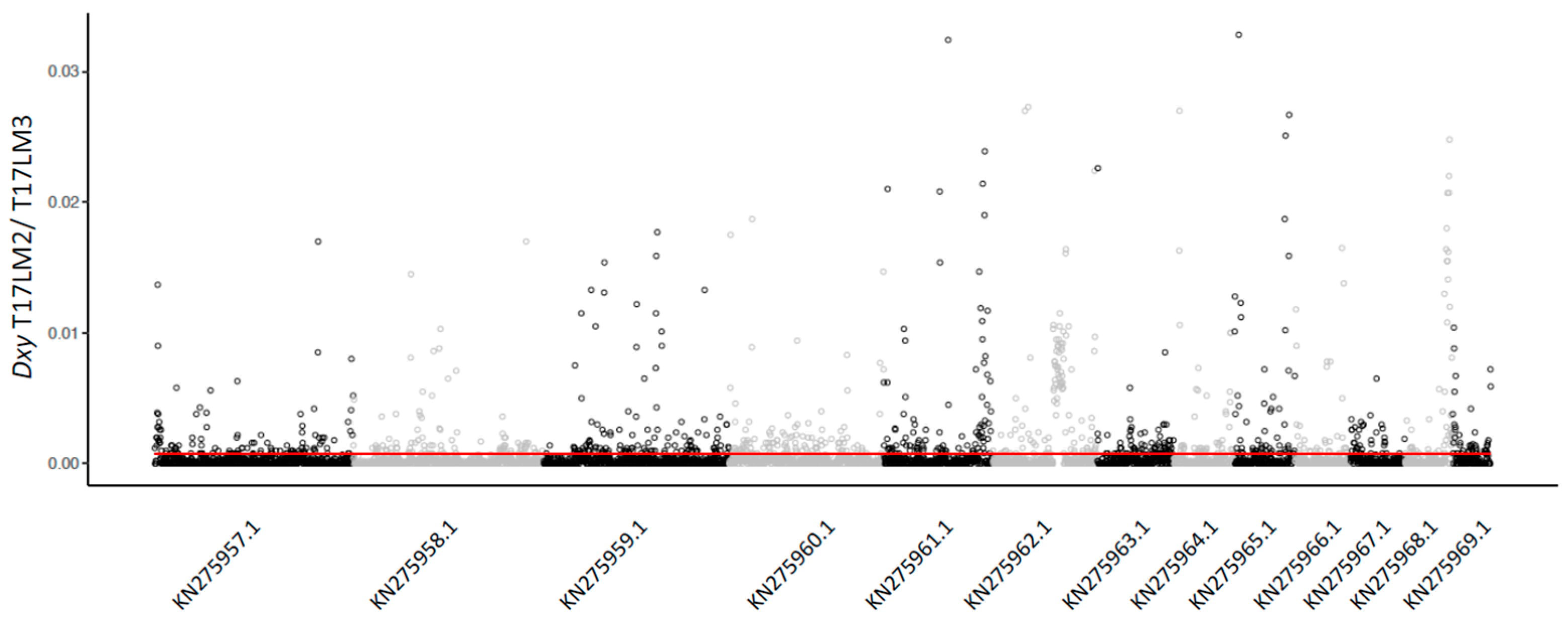
3.3. Admixture
4. Discussion
4.1. Paracoccidioides and Armadillos
4.2. Coinfection by Multiple Isolates
4.3. Population Structure and Admixture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats Posed by the Fungal Kingdom to Humans, Wildlife, and Agriculture. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, M.C.C.; Teixeira, M.M.; Barker, B.M. Fantastic yeasts and where to find them: The hidden diversity of dimorphic fungal pathogens. Curr. Opin. Microbiol. 2019, 52, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Sil, A.; Andrianopoulos, A. Thermally Dimorphic Human Fungal Pathogens—Polyphyletic Pathogens with a Convergent Pathogenicity Trait. Cold Spring Harb. Perspect. Med. 2014, 5, a019794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, G.M. Dimorphism in fungal pathogens of mammals, plants, and insects. PLoS Pathog. 2015, 11, e1004608. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Nino-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S. Paracoccidioides species complex: Ecology, phylogeny, sexual reproduction, and virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef] [Green Version]
- Theodoro, R.C.; Teixeira Mde, M.; Felipe, M.S.; Paduan Kdos, S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus paracoccidioides: Species recognition and biogeographic aspects. PLoS ONE 2012, 7, e37694. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef]
- Teixeira Mde, M.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Soares Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Melo Teixeira, M.; Emilia Cattana, M.; Matute, D.R.; Fernando Munoz, J.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; de Los Angeles Sosa, M.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fungal Genet. Biol. 2020. [Google Scholar] [CrossRef]
- Munoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.; et al. Genome Diversity, Recombination, and Virulence across the Major Lineages of Paracoccidioides. mSphere 2016, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arantes, T.D.; Theodoro, R.C.; Teixeira Mde, M.; Bosco Sde, M.; Bagagli, E. Environmental Mapping of Paracoccidioides spp. in Brazil Reveals New Clues into Genetic Diversity, Biogeography and Wild Host Association. PLoS Negl. Trop. Dis. 2016, 10, e0004606. [Google Scholar] [CrossRef] [Green Version]
- Bagagli, E.; Bosco, S.M.; Theodoro, R.C.; Franco, M. Phylogenetic and evolutionary aspects of Paracoccidioides brasiliensis reveal a long coexistence with animal hosts that explain several biological features of the pathogen. Infect. Genet. Evol. 2006, 6, 344–351. [Google Scholar] [CrossRef]
- Bagagli, E.; Sano, A.; Coelho, K.I.; Alquati, S.; Miyaji, M.; de Camargo, Z.P.; Gomes, G.M.; Franco, M.; Montenegro, M.R. Isolation of Paracoccidioides brasiliensis from armadillos (Dasypus noveminctus) captured in an endemic area of paracoccidioidomycosis. Am. J. Trop. Med. Hyg. 1998, 58, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naiff, R.D.; Ferreira, L.C.L.; Barrett, T.V.; Naiff, M.F.; Arias, J.R. Paracoccidioidomicose enzoótica em tatus (Dasypus novemcinctus) no estado do Pará. Rev. Inst. Med. Trop. São Paulo 1986, 28, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Corredor, G.G.; Peralta, L.A.; Castano, J.H.; Zuluaga, J.S.; Henao, B.; Arango, M.; Tabares, A.M.; Matute, D.R.; McEwen, J.G.; Restrepo, A. The naked-tailed armadillo Cabassous centralis (Miller 1899): A new host to Paracoccidioides brasiliensis. Molecular identification of the isolate. Med. Mycol. 2005, 43, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Hrycyk, M.F.; Garcia Garces, H.; Bosco, S.M.G.; de Oliveira, S.L.; Marques, S.A.; Bagagli, E. Ecology of Paracoccidioides brasiliensis, P. lutzii and related species: Infection in armadillos, soil occurrence and mycological aspects. Med. Mycol. 2018. [Google Scholar] [CrossRef]
- Eulalio, K.D.; de Macedo, R.L.; Cavalcanti, M.A.; Martins, L.M.; Lazera, M.S.; Wanke, B. Coccidioides immitis isolated from armadillos (Dasypus novemcinctus) in the state of Piaui, northeast Brazil. Mycopathologia 2001, 149, 57–61. [Google Scholar]
- Arias, J.R.; Naiff, R.D.; Naiff, M.F.; Mok, W.Y.; Almeida, M.M. Isolation of Histoplasma capsulatum from an armadillo (Dasypus novemcinctus) in the eastern Amazon of Brazil. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 705–706. [Google Scholar] [CrossRef]
- Vieira Gde, D.; Alves Tda, C.; Lima, S.M.; Camargo, L.M.; Sousa, C.M. Paracoccidioidomycosis in a western Brazilian Amazon State: Clinical-epidemiologic profile and spatial distribution of the disease. Rev. Soc. Bras. Med. Trop. 2014, 47, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Sahl, J.W.; Lemmer, D.; Travis, J.; Schupp, J.M.; Gillece, J.D.; Aziz, M.; Driebe, E.M.; Drees, K.P.; Hicks, N.D.; Williamson, C.H.D.; et al. NASP: An accurate, rapid method for the identification of SNPs in WGS datasets that supports flexible input and output formats. Microb. Genom. 2016, 2, e000074. [Google Scholar] [CrossRef] [PubMed]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepulveda, V.E.; Marquez, R.; Turissini, D.A.; Goldman, W.E.; Matute, D.R. Genome Sequences Reveal Cryptic Speciation in the Human Pathogen Histoplasma capsulatum. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, R.K.; Ramasamy, S.; Bindroo, B.B.; Naik, V.G. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. Springerplus 2014, 3, 431. [Google Scholar] [CrossRef] [Green Version]
- Borelli, D. Algunos aspectos ecologicos de la paracoccidioidosis. Derm. Venez. 1971, 10, 11. [Google Scholar]
- Richini-Pereira, V.B.; Bosco Sde, M.; Griese, J.; Theodoro, R.C.; Macoris, S.A.; da Silva, R.J.; Barrozo, L.; Tavares, P.M.; Zancope-Oliveira, R.M.; Bagagli, E. Molecular detection of Paracoccidioides brasiliensis in road-killed wild animals. Med. Mycol. 2008, 46, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Corredor, G.G.; Castano, J.H.; Peralta, L.A.; Diez, S.; Arango, M.; McEwen, J.; Restrepo, A. Isolation of Paracoccidioides brasiliensis from the nine-banded armadillo Dasypus novemcinctus, in an endemic area for paracoccidioidomycosis in Colombia. Rev. Iberoam. Micol. 1999, 16, 216–220. [Google Scholar]
- de Farias, M.R.; Condas, L.A.; Ribeiro, M.G.; Bosco Sde, M.; Muro, M.D.; Werner, J.; Theodoro, R.C.; Bagagli, E.; Marques, S.A.; Franco, M. Paracoccidioidomycosis in a dog: Case report of generalized lymphadenomegaly. Mycopathologia 2011, 172, 147–152. [Google Scholar] [CrossRef]
- Ono, M.A.; Bracarense, A.P.; Morais, H.S.; Trapp, S.M.; Belitardo, D.R.; Camargo, Z.P. Canine paracoccidioidomycosis: A seroepidemiologic study. Med. Mycol. 2001, 39, 277–282. [Google Scholar] [CrossRef]
- Trejo-Chavez, A.; Ramirez-Romero, R.; Ancer-Rodriguez, J.; Nevarez-Garza, A.M.; Rodriguez-Tovar, L.E. Disseminated paracoccidioidomycosis in a Southern two-toed sloth (Choloepus didactylus). J. Comp. Pathol. 2011, 144, 231–234. [Google Scholar] [CrossRef]
- Mendes, J.F.; Klafke, G.B.; Albano, A.P.; Cabana, A.L.; Teles, A.J.; de Camargo, Z.P.; Xavier, M.O.; Meireles, M.C. Paracoccidioidomycosis infection in domestic and wild mammals by Paracoccidioides lutzii. Mycoses 2017. [Google Scholar] [CrossRef] [PubMed]
- Hull, C.M.; Raisner, R.M.; Johnson, A.D. Evidence for mating of the "asexual" yeast Candida albicans in a mammalian host. Science 2000, 289, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Mavengere, H.; Mattox, K.; Teixeira, M.M.; Sepúlveda, V.E.; Gomez, O.M.; Hernandez, O.; McEwen, J.; Matute, D.R. Paracoccidioides genomes reflect high levels of species divergence and little interspecific gene flow. bioRxiv 2020. [Google Scholar] [CrossRef]
- Maxwell, C.S.; Sepulveda, V.E.; Turissini, D.A.; Goldman, W.E.; Matute, D.R. Recent admixture between species of the fungal pathogen Histoplasma. Evol. Lett. 2018, 2, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, C.S.; Mattox, K.; Turissini, D.A.; Teixeira, M.M.; Barker, B.M.; Matute, D.R. Gene exchange between two divergent species of the fungal human pathogen, Coccidioides. Evolution 2018. [Google Scholar] [CrossRef] [Green Version]
- Matute, D.R.; Comeault, A.A.; Earley, E.; Serrato-Capuchina, A.; Peede, D.; Monroy-Eklund, A.; Huang, W.; Jones, C.D.; Mackay, T.F.C.; Coyne, J.A. Rapid and Predictable Evolution of Admixed Populations Between Two Drosophila Species Pairs. Genetics 2020, 214, 211–230. [Google Scholar] [CrossRef]
- Martin, S.H.; Davey, J.W.; Salazar, C.; Jiggins, C.D. Recombination rate variation shapes barriers to introgression across butterfly genomes. PLoS Biol. 2019, 17, e2006288. [Google Scholar] [CrossRef] [Green Version]
- Schumer, M.; Xu, C.; Powell, D.L.; Durvasula, A.; Skov, L.; Holland, C.; Blazier, J.C.; Sankararaman, S.; Andolfatto, P.; Rosenthal, G.G.; et al. Natural selection interacts with recombination to shape the evolution of hybrid genomes. Science 2018, 360, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Moyle, L.C.; Nakazato, T. Hybrid incompatibility “snowballs” between Solanum species. Science 2010, 329, 1521–1523. [Google Scholar] [CrossRef]
- Matute, D.R.; Butler, I.A.; Turissini, D.A.; Coyne, J.A. A test of the snowball theory for the rate of evolution of hybrid incompatibilities. Science 2010, 329, 1518–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.J.; White, M.A.; Payseur, B.A. The Pace of Hybrid Incompatibility Evolution in House Mice. Genetics 2015, 201, 229–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhina, A.K.; Thompson, K.A.; Schluter, D. Adaptive divergence and the evolution of hybrid phenotypes in threespine stickleback. bioRxiv 2020. [Google Scholar] [CrossRef]
- Macedo, R.L.; Lazera Mdos, S.; Trilles, L.; Reisr, R.S.; Bulcão, A.S.; Silva, N.J.; Oliveira, N.A.; Wanke, B. Infecção natural de tatus por Paracoccidioides brasiliensis em serra da mesa, Minaçu, Goiás: Estudo preliminar. In Proceedings of the II Congresso Brasileiro de Micologia, Rio de Janeiro, Brazil, 17–21 April 1998. [Google Scholar]
- Vergara, M.L.; Martinez, R. Role of the armadillo Dasypus novemcinctus in the epidemiology of paracoccidioidomycosis. Mycopathologia 1998, 144, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Naiff, R.D.; Barret, T.V. Novos registros de Paracoccidioides brasiliensis em tatus (Dasypus novemcinctus). In Proceedings of the Congresso Brasileiro Parasitologia, Rio de Janeiro, Brazil, 30 July–3 August 1989. [Google Scholar]



{kind=link}
{kind=link}
| Strain | Host | Sex | Host Weight (kg) | Geographic Area | Coordinates * | Organ | Genotype |
|---|---|---|---|---|---|---|---|
| T17LM2 | D. novemcinctus | male | 4.7 | Southeast, SP (Botucatu) | 22°48′02″ S/48°23′24″ W | Mesenteric Lymph node | S1a |
| T17LM3 | D. novemcinctus | male | 4.7 | Southeast, SP (Botucatu) | 22°48′02″ S/48°23′24″ W | Mesenteric Lymph node | S1a |
| T18LM1 | D. novemcinctus | male | 3.9 | Southeast, SP (Botucatu) | 22°48′02″ S/48°23′24″ W | Mesenteric Lymph node | S1a |
| T22LM1 | D. novemcinctus | female | 3.9 | Southeast, SP (São Manoel) | 22°34′16″ S/48°25′18″ W | Mesenteric Lymph node | S1a |
| T23LM1 | D. novemcinctus | male | 4.3 | Southeast, SP (São Manoel) | 22°34′16″ S/48°25′18″ W | Mesenteric Lymph node | S1a |
| T19F33 | D. novemcinctus | male | 6.0 | Midwest, MT (Alta Floresta) | 10°04′23″ S/56°09′34″ W | Liver | S1a/S1b |
| T20B15 | D. novemcinctus | male | 6.0 | Midwest, MT (Alta Floresta) | 10°04′23″ S/56°09′34″ W | Spleen | S1a/S1b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagagli, E.; Matute, D.R.; Garces, H.G.; Tenório, B.G.; Garces, A.G.; Alves, L.G.d.B.; Yamauchi, D.H.; Hrycyk, M.F.; Barker, B.M.; Teixeira, M.d.M. Paracoccidioides brasiliensis Isolated from Nine-Banded Armadillos (Dasypus novemcinctus) Reveal Population Structure and Admixture in the Amazon Basin. J. Fungi 2021, 7, 54. https://doi.org/10.3390/jof7010054
Bagagli E, Matute DR, Garces HG, Tenório BG, Garces AG, Alves LGdB, Yamauchi DH, Hrycyk MF, Barker BM, Teixeira MdM. Paracoccidioides brasiliensis Isolated from Nine-Banded Armadillos (Dasypus novemcinctus) Reveal Population Structure and Admixture in the Amazon Basin. Journal of Fungi. 2021; 7(1):54. https://doi.org/10.3390/jof7010054
Chicago/Turabian StyleBagagli, Eduardo, Daniel Ricardo Matute, Hans Garcia Garces, Bernardo Guerra Tenório, Adalberto Garcia Garces, Lucas Gomes de Brito Alves, Danielle Hamae Yamauchi, Marluce Francisca Hrycyk, Bridget Marie Barker, and Marcus de Melo Teixeira. 2021. "Paracoccidioides brasiliensis Isolated from Nine-Banded Armadillos (Dasypus novemcinctus) Reveal Population Structure and Admixture in the Amazon Basin" Journal of Fungi 7, no. 1: 54. https://doi.org/10.3390/jof7010054
APA StyleBagagli, E., Matute, D. R., Garces, H. G., Tenório, B. G., Garces, A. G., Alves, L. G. d. B., Yamauchi, D. H., Hrycyk, M. F., Barker, B. M., & Teixeira, M. d. M. (2021). Paracoccidioides brasiliensis Isolated from Nine-Banded Armadillos (Dasypus novemcinctus) Reveal Population Structure and Admixture in the Amazon Basin. Journal of Fungi, 7(1), 54. https://doi.org/10.3390/jof7010054