
Fungal Mobilization of Selenium in the Presence of Hausmannite and Ferric Oxyhydroxides

 ,
,  ,
,  , and
, and 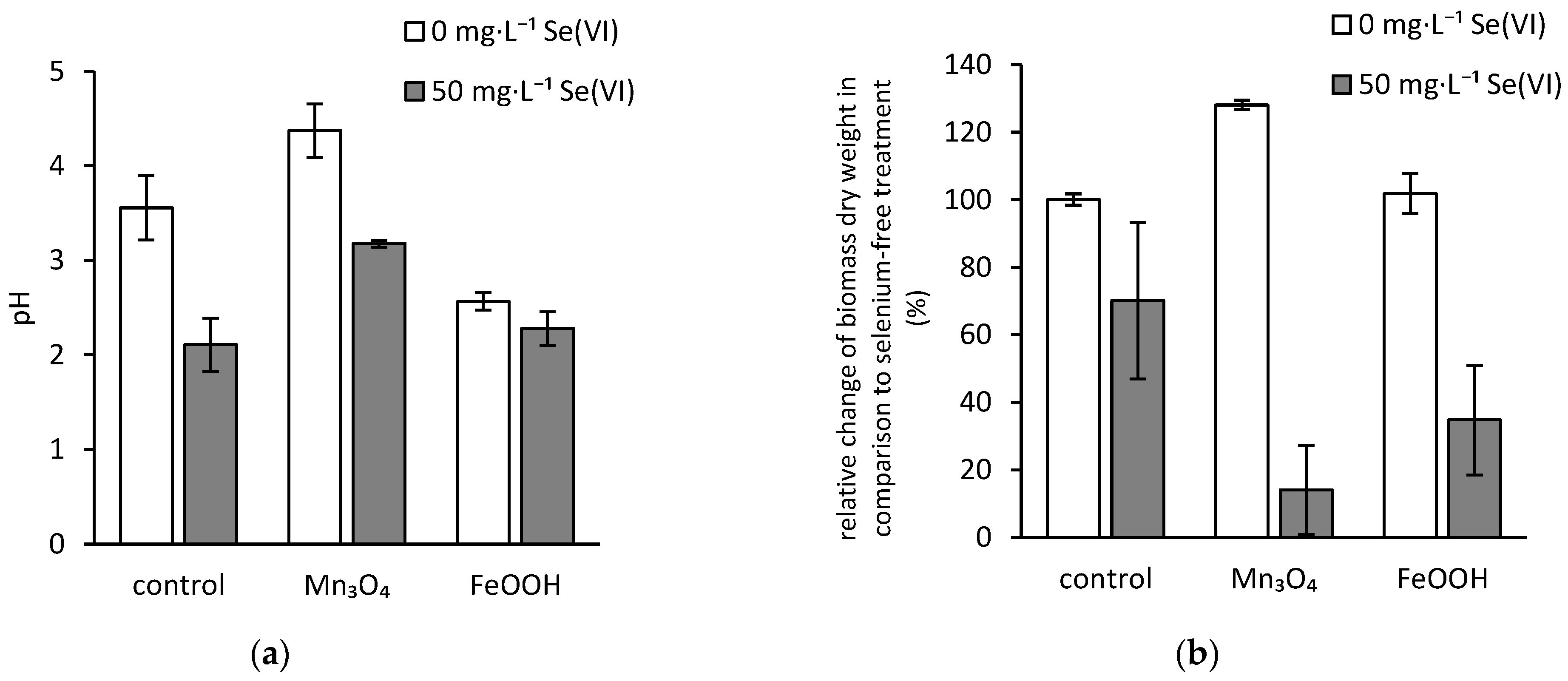{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Ferric Oxyhydroxides and Hausmannite Synthesis
2.3. Bioleaching
2.4. Analytics
3. Results
3.1. pH of the Culture Media
3.2. Dry Biomass Weight
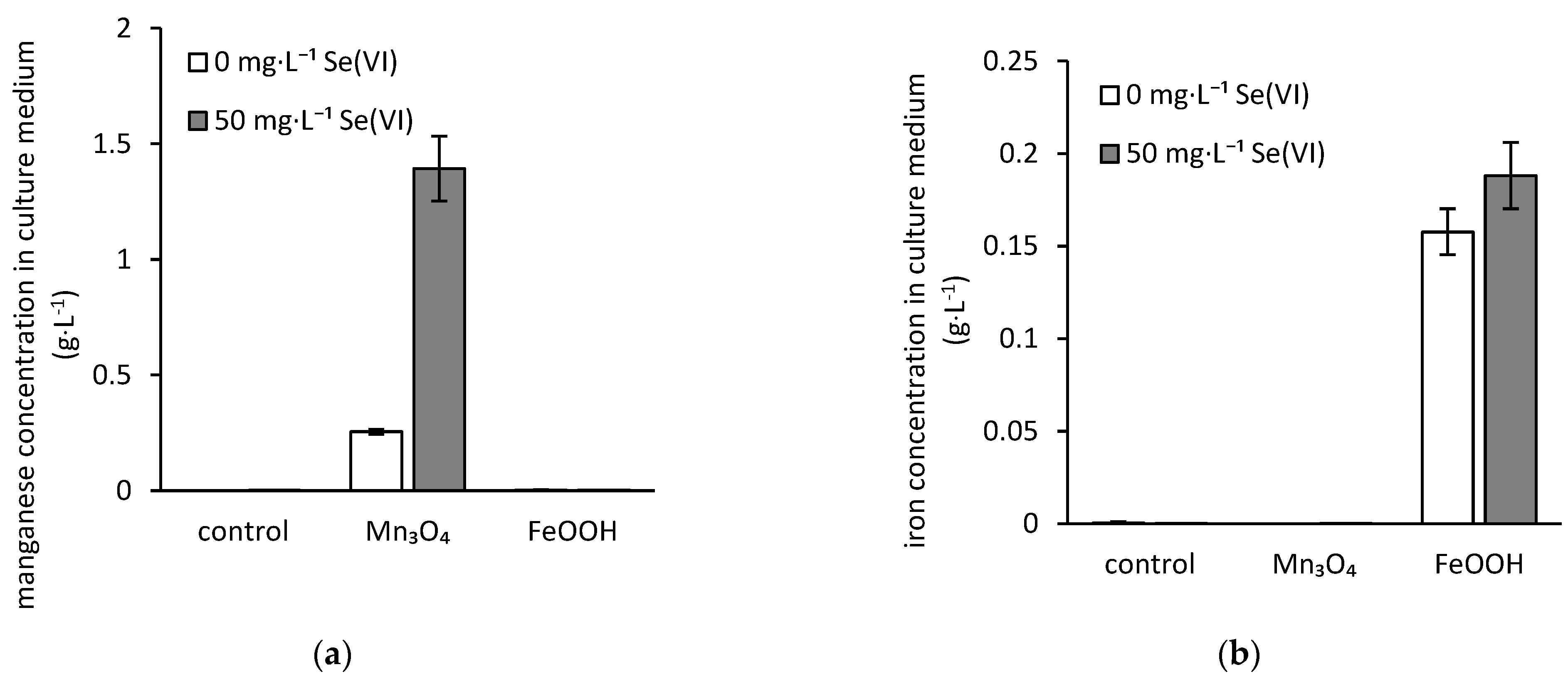
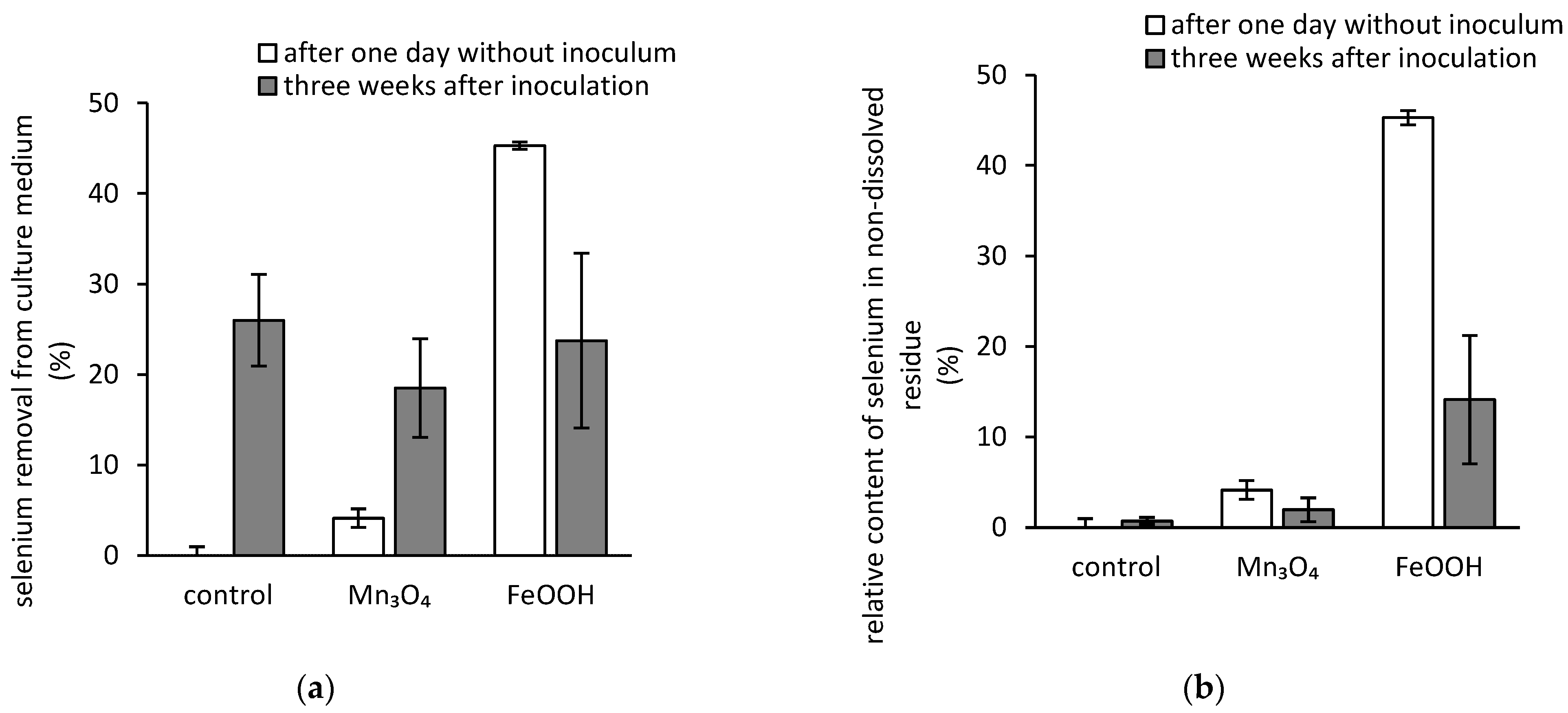
3.3. Bioextraction of Iron, Manganese, and Selenium
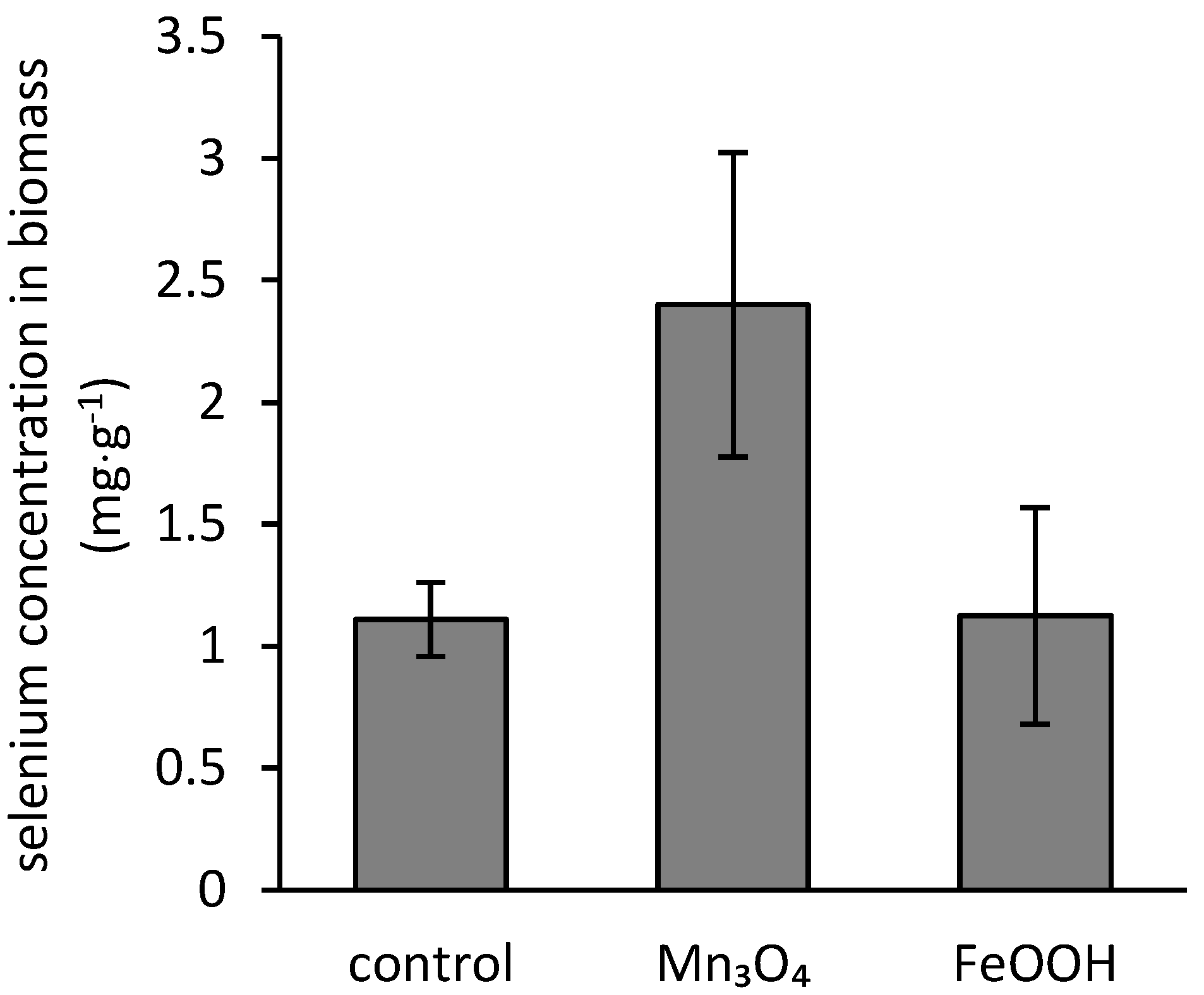
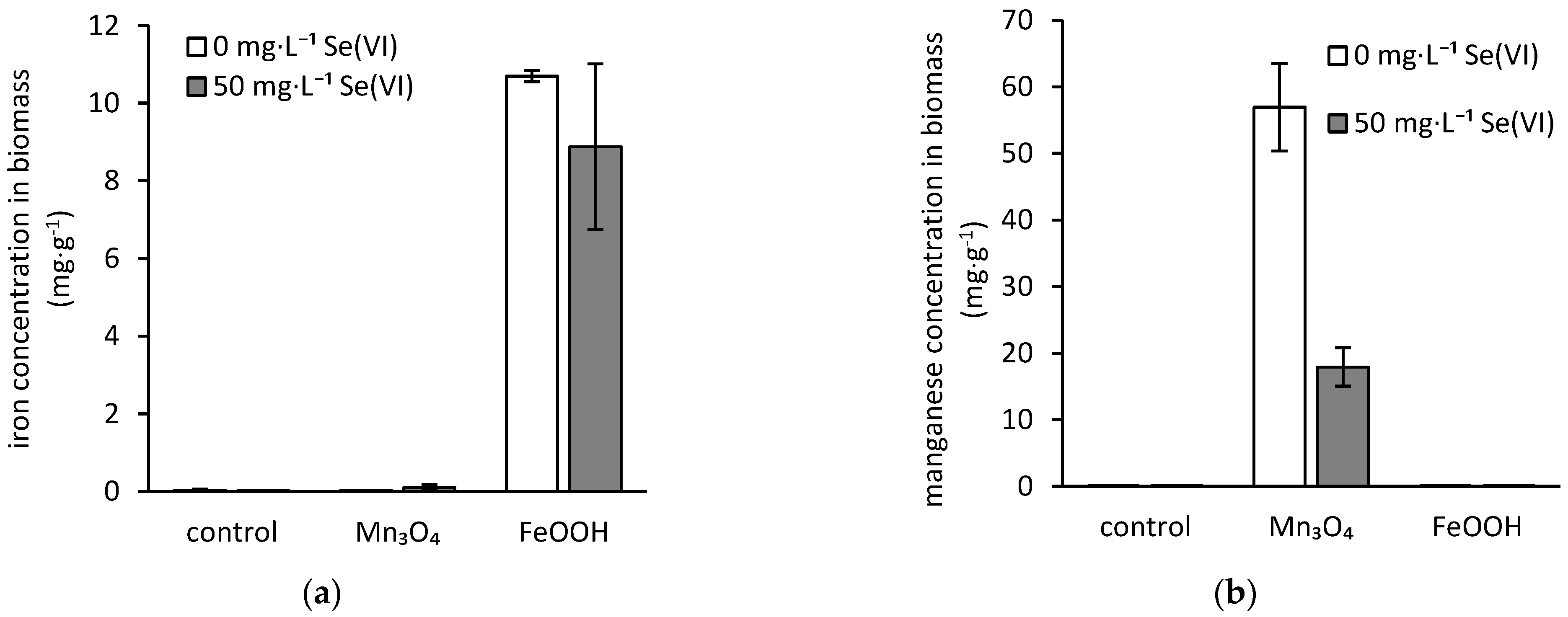
3.4. Bioaccumulation of Iron, Manganese, and Selenium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, C.; Yu, Z.-G.; Zeng, G.-M.; Jiang, M.; Yang, Z.-Z.; Cui, F.; Zhu, M.-Y.; Shen, L.-Q.; Hu, L. Effects of sediment geochemical properties on heavy metal bioavailability. Environ. Int. 2014, 73, 270–281. [Google Scholar] [CrossRef]
- Lee, G.; Bigham, J.M.; Faure, G. Removal of trace metals by coprecipitation with Fe, Al and Mn from natural waters contaminated with acid mine drainage in the Ducktown Mining District, Tennessee. Appl. Geochem. 2002, 17, 569–581. [Google Scholar] [CrossRef]
- Sipos, P.; Choi, C.; Németh, T.; Szalai, Z.; Póka, T. Relationship between iron and trace metal fractionation in soils. Chem. Speciat. Bioavailab. 2014, 26, 21–30. [Google Scholar] [CrossRef]
- Post, J.E. Manganese oxide minerals: Crystal structures and economic and environmental significance. Proc. Natl. Acad. Sci. USA 1999, 96, 3447–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korobova, E.M.; Ryzhenko, B.N.; Cherkasova, E.V.; Sedykh, E.M.; Korsakova, N.V.; Danilova, V.N.; Khushvakhtova, S.D.; Berezkin, V.Y. Iodine and selenium speciation in natural waters and their concentrating at landscape-geochemical barriers. Geochem. Int. 2014, 52, 500–514. [Google Scholar] [CrossRef]
- Dudová, J.; Bujdoš, M. Study of selenium sorption on iron oxide hydroxides. Chem. Listy 2015, 109, 770–774. [Google Scholar]
- Hagarová, I.; Nemček, L. Selenium in Blood Serum of Healthy European Population. Chem. Listy 2020, 114, 329–335. [Google Scholar]
- Hagarová, I.; Žemberyová, M. Determination of selenium in blood serum of children by electrothermal atomic absorption spectrometry. Chem. Listy 2005, 99, 34–39. [Google Scholar]
- Lenz, M.; Lens, P.N.L. The essential toxin: The changing perception of selenium in environmental sciences. Sci. Total Environ. 2009, 407, 3620–3633. [Google Scholar] [CrossRef]
- He, Y.; Xiang, Y.; Zhou, Y.; Yang, Y.; Zhang, J.; Huang, H.; Shang, C.; Luo, L.; Gao, J.; Tang, L. Selenium contamination, consequences and remediation techniques in water and soils: A review. Environ. Res. 2018, 164, 288–301. [Google Scholar] [CrossRef]
- Hagarová, I.; Žemberyová, M.; Bajčan, D. Sequential and single step extraction procedures used for fractionation of selenium in soil samples. Chem. Pap. 2005, 59, 93–98. [Google Scholar]
- Durán, P.; Acuña, J.J.; Jorquera, M.A.; Azcón, R.; Borie, F.; Cornejo, P.; Mora, M.L. Enhanced selenium content in wheat grain by co-inoculation of selenobacteria and arbuscular mycorrhizal fungi: A preliminary study as a potential Se biofortification strategy. J. Cereal Sci. 2013, 57, 275–280. [Google Scholar] [CrossRef]
- Perkins, W.T. Extreme selenium and tellurium contamination in soils—An eighty year-old industrial legacy surrounding a Ni refinery in the Swansea Valley. Sci. Total Environ. 2011, 412–413, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Schuler, C.A.; Anthony, R.G.; Ohlendorf, H.M. Selenium in wetlands and waterfowl foods at Kesterson Reservoir, California, 1984. Arch. Environ. Contam. Toxicol. 1990, 19, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Börsig, N.; Scheinost, A.C.; Shaw, S.; Schild, D.; Neumann, T. Uptake mechanisms of selenium oxyanions during the ferrihydrite-hematite recrystallization. Geochim. Cosmochim. Acta 2017, 206, 236–253. [Google Scholar] [CrossRef] [Green Version]
- Čanecká, L.; Bujdoš, M.; Boriová, K. Hydroxides and their potential utilization in reducing the content of some undesirable elements in the natural environment by sorption mechanisms. Chem. Listy 2015, 109, 105–108. [Google Scholar]
- Li, Z.; Yuan, Y.; Ma, L.; Zhang, Y.; Jiang, H.; He, J.; Hu, Y.; Yuan, S.; Ginder-Vogel, M.; Tu, S. Simultaneous Kinetics of Selenite Oxidation and Sorption on δ-MnO(2) in Stirred-Flow Reactors. Int. J. Environ. Res. Public Health 2021, 18, 2902. [Google Scholar] [CrossRef]
- Mayanna, S.; Peacock, C.L.; Schäffner, F.; Grawunder, A.; Merten, D.; Kothe, E.; Büchel, G. Biogenic precipitation of manganese oxides and enrichment of heavy metals at acidic soil pH. Chem. Geol. 2015, 402, 6–17. [Google Scholar] [CrossRef]
- Farkas, B.; Kolenčík, M.; Hain, M.; Dobročka, E.; Kratošová, G.; Bujdoš, M.; Feng, H.; Deng, Y.; Yu, Q.; Illa, R.; et al. Aspergillus niger decreases bioavailability of arsenic(V) via biotransformation of manganese oxide into biogenic oxalate minerals. J. Fungi 2020, 6, 270. [Google Scholar] [CrossRef]
- Miglierini, M.B.; Dekan, J.; Urík, M.; Cesnek, M.; Kmječ, T.; Matúš, P. Fungal-induced modification of spontaneously precipitated ochreous sediments from drainage of abandoned antimony mine. Chemosphere 2021, 269, 128733. [Google Scholar] [CrossRef]
- Duborská, E.; Szabó, K.; Bujdoš, M.; Vojtková, H.; Littera, P.; Dobročka, E.; Kim, H.; Urík, M. Assessment of Aspergillus niger strain’s suitability for arsenate-contaminated water treatment and adsorbent recycling via bioextraction in a laboratory-scale experiment. Microorganisms 2020, 8, 1668. [Google Scholar] [CrossRef] [PubMed]
- Milová-Žiaková, B.; Urík, M.; Boriová, K.; Bujdoš, M.; Kolenčík, M.; Mikušová, P.; Takáčová, A.; Matúš, P. Fungal solubilization of manganese oxide and its significance for antimony mobility. Int. Biodeterior. Biodegrad. 2016, 114, 157–163. [Google Scholar] [CrossRef]
- Urík, M.; Polák, F.; Bujdoš, M.; Miglierini, M.B.; Milová-Žiaková, B.; Farkas, B.; Goneková, Z.; Vojtková, H.; Matúš, P. Antimony leaching from antimony-bearing ferric oxyhydroxides by filamentous fungi and biotransformation of ferric substrate. Sci. Total Environ. 2019, 664, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, B.J.; Loeppert, R.H. Methyl arsenic adsorption and desorption behavior on iron oxides. Environ. Sci. Technol. 2005, 39, 2120–2127. [Google Scholar] [CrossRef]
- Urík, M.; Hlodák, M.; Mikušová, P.; Matúš, P. Potential of microscopic fungi isolated from mercury contaminated soils to accumulate and volatilize mercury(II). Water Air Soil Pollut. 2014, 225, 2219. [Google Scholar] [CrossRef]
- Matulová, M.; Urík, M.; Bujdoš, M.; Duborská, E.; Cesnek, M.; Miglierini, M.B. Selenite sorption onto goethite: Isotherm and ion-competitive studies, and effect of pH on sorption kinetics. Chem. Pap. 2019, 73, 2975–2985. [Google Scholar] [CrossRef]
- Matulová, M.; Bujdoš, M.; Miglierini, M.B.; Mitróová, Z.; Kubovčíková, M.; Urík, M. The effects of selenate on goethite synthesis and selenate sorption kinetics onto a goethite surface—A three-step process with an unexpected desorption phase. Chem. Geol. 2020, 556, 119852. [Google Scholar] [CrossRef]
- Marjanovic, V.; Peric-Grujic, A.; Ristic, M.; Marinkovic, A.; Markovic, R.; Onjia, A.; Sljivic-Ivanovic, M. Selenate adsorption from water using the hydrous iron oxide-impregnated hybrid polymer. Metals 2020, 10, 1630. [Google Scholar] [CrossRef]
- Zelmanov, G.; Semiat, R. Selenium removal from water and its recovery using iron (Fe3+) oxide/hydroxide-based nanoparticles sol (NanoFe) as an adsorbent. Sep. Purif. Technol. 2013, 103, 167–172. [Google Scholar] [CrossRef]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [Green Version]
- Odoni, D.I.; van Gaal, M.P.; Schonewille, T.; Tamayo-Ramos, J.A.; Martins dos Santos, V.A.P.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger Secretes Citrate to Increase Iron Bioavailability. Front. Microbiol. 2017, 8, 1424. [Google Scholar] [CrossRef]
- Urík, M.; Polák, F.; Bujdoš, M.; Pifková, I.; Kořenková, L.; Littera, P.; Matúš, P. Aluminium Leaching by Heterotrophic Microorganism Aspergillus niger: An Acidic Leaching? Arab. J. Sci. Eng. 2018, 43, 2369–2374. [Google Scholar] [CrossRef]
- Osman, Y.; Gebreil, A.; Mowafy, A.M.; Anan, T.I.; Hamed, S.M. Characterization of Aspergillus niger siderophore that mediates bioleaching of rare earth elements from phosphorites. World J. Microbiol. Biotechnol. 2019, 35, 93. [Google Scholar] [CrossRef] [PubMed]
- Odoni, D.I.; Vazquez-Vilar, M.; van Gaal, M.P.; Schonewille, T.; Santos, V.A.P.M.D.; Tamayo-Ramos, J.A.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger citrate exporter revealed by comparison of two alternative citrate producing conditions. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.R.; Lehmann, L.; Nielsen, J. Systemic analysis of the response of Aspergillus niger to ambient pH. Genome Biol. 2009, 10, R47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taxiarchou, M.; Panias, D.; Douni, I.; Paspaliaris, I.; Kontopoulos, A. Dissolution of hematite in acidic oxalate solutions. Hydrometallurgy 1997, 44, 287–299. [Google Scholar] [CrossRef]
- Polák, F.; Urík, M.; Bujdoš, M.; Matúš, P. Aspergillus niger enhances oxalate production as a response to phosphate deficiency induced by aluminium(III). J. Inorg. Biochem. 2020, 204, 110961. [Google Scholar] [CrossRef]
- Polák, F.; Urík, M.; Bujdoš, M.; Uhlík, P.; Matúš, P. Evaluation of aluminium mobilization from its soil mineral pools by simultaneous effect of Aspergillus strains’ acidic and chelating exometabolites. J. Inorg. Biochem. 2018, 181, 162–168. [Google Scholar] [CrossRef]
- Valix, M.; Usai, F.; Malik, R. Fungal bio-leaching of low grade laterite ores. Miner. Eng. 2001, 14, 197–203. [Google Scholar] [CrossRef]
- Urík, M.; Boriová, K.; Bujdoš, M.; Matúš, P. Fungal Selenium(VI) Accumulation and Biotransformation—Filamentous Fungi in Selenate Contaminated Aqueous Media Remediation. CLEAN—Soil Air Water 2016, 44, 610–614. [Google Scholar] [CrossRef]
- Fink, J.R.; Inda, A.V.; Tiecher, T.; Barrón, V. Iron oxides and organic matter on soil phosphorus availability. Cienc. Agrotecnol. 2016, 40, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Dusengemungu, L.; Kasali, G.; Gwanama, C.; Mubemba, B. Overview of fungal bioleaching of metals. Environ. Adv. 2021, 5, 100083. [Google Scholar] [CrossRef]
- Urík, M.; Farkas, B.; Miglierini, M.B.; Bujdoš, M.; Mitróová, Z.; Kim, H.; Matúš, P. Mobilisation of hazardous elements from arsenic-rich mine drainage ochres by three Aspergillus species. J. Hazard. Mater. 2021, 409, 124938. [Google Scholar] [CrossRef]
- Sabuda, M.C.; Rosenfeld, C.E.; DeJournett, T.D.; Schroeder, K.; Wuolo-Journey, K.; Santelli, C.M. Fungal Bioremediation of Selenium-Contaminated Industrial and Municipal Wastewaters. Front. Microbiol. 2020, 11, 2105. [Google Scholar] [CrossRef]
- Weissman, G.S.; Trelease, S.F. Influence of sulfur on the toxicity of selenium to Aspergillus. Am. J. Bot. 1955, 42, 489–495. [Google Scholar] [CrossRef]
- Casado, M.; Anawar, H.M.; Garcia-Sanchez, A.; Regina, I.S. Arsenic Bioavailability in Polluted Mining Soils and Uptake by Tolerant Plants (El Cabaco mine, Spain). Bull. Environ. Contam. Toxicol. 2007, 79, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.E.; Foster, A.L.; Ostergren, J.D. Mineral surfaces and bioavailability of heavy metals: A molecular-scale perspective. Proc. Natl. Acad. Sci. USA 1999, 96, 3388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Loi, G.; Otero-Gonzalez, L.; Laing, G.D.; Ferrer, I.; Lens, P.N.L. Selenate and selenite uptake, accumulation and toxicity in Lemna minuta. Water. Sci. Technol. 2020, 81, 1852–1862. [Google Scholar] [CrossRef] [PubMed]
- Balistrieri, L.S.; Chao, T.T. Adsorption of selenium by amorphous iron oxyhydroxide and manganese dioxide. Geochim. Cosmochim. Acta 1990, 54, 739–751. [Google Scholar] [CrossRef]
- Tang, Y.; Zeiner, C.A.; Santelli, C.M.; Hansel, C.M. Fungal oxidative dissolution of the Mn(II)-bearing mineral rhodochrosite and the role of metabolites in manganese oxide formation. Environ. Microbiol. 2013, 15, 1063–1077. [Google Scholar] [CrossRef]
- Wei, Z.; Hillier, S.; Gadd, G.M. Biotransformation of manganese oxides by fungi: Solubilization and production of manganese oxalate biominerals. Environ. Microbiol. 2012, 14, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Godunov, E.B.; Artamonova, I.V.; Gorichev, I.G.; Lainer, Y.A. Influence of oxalic acid on the dissolution kinetics of manganese oxide. Russ. Metall. 2012, 2012, 935–941. [Google Scholar] [CrossRef]
- Babich, H.; Stotzky, G. Manganese toxicity to fungi: Influence of pH. Bull. Environ. Contam. Toxicol. 1981, 27, 474–480. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farkas, B.; Vojtková, H.; Bujdoš, M.; Kolenčík, M.; Šebesta, M.; Matulová, M.; Duborská, E.; Danko, M.; Kim, H.; Kučová, K.; et al. Fungal Mobilization of Selenium in the Presence of Hausmannite and Ferric Oxyhydroxides. J. Fungi 2021, 7, 810. https://doi.org/10.3390/jof7100810
Farkas B, Vojtková H, Bujdoš M, Kolenčík M, Šebesta M, Matulová M, Duborská E, Danko M, Kim H, Kučová K, et al. Fungal Mobilization of Selenium in the Presence of Hausmannite and Ferric Oxyhydroxides. Journal of Fungi. 2021; 7(10):810. https://doi.org/10.3390/jof7100810
Chicago/Turabian StyleFarkas, Bence, Hana Vojtková, Marek Bujdoš, Marek Kolenčík, Martin Šebesta, Michaela Matulová, Eva Duborská, Martin Danko, Hyunjung Kim, Kateřina Kučová, and et al. 2021. "Fungal Mobilization of Selenium in the Presence of Hausmannite and Ferric Oxyhydroxides" Journal of Fungi 7, no. 10: 810. https://doi.org/10.3390/jof7100810
APA StyleFarkas, B., Vojtková, H., Bujdoš, M., Kolenčík, M., Šebesta, M., Matulová, M., Duborská, E., Danko, M., Kim, H., Kučová, K., Kisová, Z., Matúš, P., & Urík, M. (2021). Fungal Mobilization of Selenium in the Presence of Hausmannite and Ferric Oxyhydroxides. Journal of Fungi, 7(10), 810. https://doi.org/10.3390/jof7100810