
Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi


 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Spectrum of Activity
3. Indication
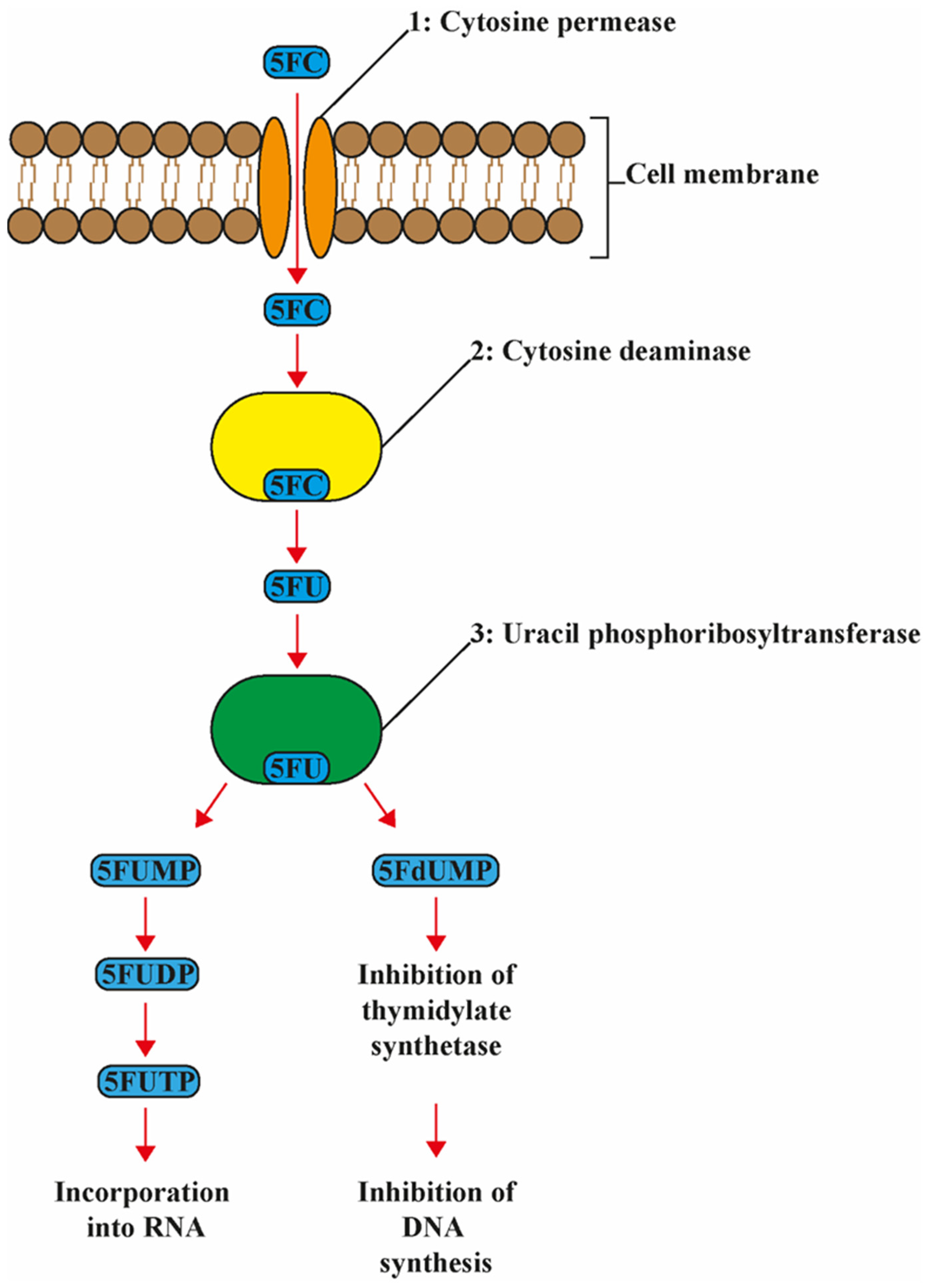
4. Mode of Action
5. Pharmacokinetics
6. Toxicity
7. 5-FC Resistance
7.1. Saccharomyces cerevisiae
7.2. Candida
7.2.1. Candida albicans
7.2.2. Candida dubliniensis
7.2.3. Candida lusitaniae
7.2.4. Candida glabrata
7.2.5. Candida tropicalis
7.2.6. Candida auris
7.3. Cryptococcus
7.4. Aspergillus
7.5. Dermatophytes
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scorzoni, L.; de Paula, E.S.A.C.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Kanafani, Z.A.; Perfect, J.R. Antimicrobial resistance: Resistance to antifungal agents: Mechanisms and clinical impact. Clin. Infect. Dis. 2008, 46, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.; Schalekamp, S.; van der Velden, W.; Kuiper, E.J.; van Paassen, J.; van der Hoven, B.; van der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermes, A.; Guchelaar, H.J.; Dankert, J. Flucytosine: A review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 2000, 46, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Ghannoum, M.A. Flucytosine Treatment and Resistance Mechanisms. In Antimicrobial Drug Resistance; Mayers, D.L., Sobel, J.D., Ouellette, M., Kaye, K.S., Marchaim, D., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 407–413. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J. Clinical practice guideline for the management of candidiasis: 2016 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourad, A.; Perfect, J.R. Present and Future Therapy of Cryptococcus Infections. J. Fungi 2018, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Messer, S.A.; Boyken, L.; Huynh, H.; Hollis, R.J.; Diekema, D.J. In vitro activities of 5-fluorocytosine against 8803 clinical isolates of Candida spp.: Global assessment of primary resistance using National Committee for Clinical Laboratory Standards susceptibility testing methods. Antimicrob. Agents Chemother. 2002, 46, 3518–3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Lopez, A.; Zaragoza, O.; Dos Anjos Martins, M.; Melhem, M.C.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. In vitro susceptibility of Cryptococcus gattii clinical isolates. Clin. Microbiol. Infect. 2008, 14, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Borman, A.M.; Muller, J.; Walsh-Quantick, J.; Szekely, A.; Patterson, Z.; Palmer, M.D.; Fraser, M.; Johnson, E.M. MIC distributions for amphotericin B, fluconazole, itraconazole, voriconazole, flucytosine and anidulafungin and 35 uncommon pathogenic yeast species from the UK determined using the CLSI broth microdilution method. J. Antimicrob. Chemother. 2020, 75, 1194–1205. [Google Scholar] [CrossRef]
- Johnson, M.D.; Perfect, J.R. Use of Antifungal Combination Therapy: Agents, Order, and Timing. Curr. Fungal Infect. Rep. 2010, 4, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molloy, S.F.; Kanyama, C.; Heyderman, R.S.; Loyse, A.; Kouanfack, C.; Chanda, D.; Mfinanga, S.; Temfack, E.; Lakhi, S.; Lesikari, S. Antifungal combinations for treatment of cryptococcal meningitis in Africa. N. Engl. J. Med. 2018, 378, 1004–1017. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Diagnosis, Prevention, and Management of Cryptococcal Disease in HIV-Infected Adults, Adolescents and Children, March 2018: Supplement to the 2016 Consolidated Guidelines of the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Stone, N.R.; Rhodes, J.; Fisher, M.C.; Mfinanga, S.; Kivuyo, S.; Rugemalila, J.; Segal, E.S.; Needleman, L.; Molloy, S.F.; Kwon-Chung, J. Dynamic ploidy changes drive fluconazole resistance in human cryptococcal meningitis. J. Clin. Investig. 2019, 129, 999–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwer, A.E.; van Kan, H.J.; Johnson, E.; Rajanuwong, A.; Teparrukkul, P.; Wuthiekanun, V.; Chierakul, W.; Day, N.; Harrison, T.S. Oral versus intravenous flucytosine in patients with human immunodeficiency virus-associated cryptococcal meningitis. Antimicrob. Agents Chemother. 2007, 51, 1038–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneale, M.; Bartholomew, J.S.; Davies, E.; Denning, D.W. Global access to antifungal therapy and its variable cost. J. Antimicrob. Chemother. 2016, 71, 3599–3606. [Google Scholar] [CrossRef] [PubMed]
- Cutler, R.E.; Blair, A.D.; Kelly, M.R. Flucytosine kinetics in subjects with normal and impaired renal function. Clin. Pharmacol. Ther. 1978, 24, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Block, E.R.; Bennett, J.E. Pharmacological studies with 5-fluorocytosine. Antimicrob. Agents Chemother. 1972, 1, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Daneshmend, T.K.; Warnock, D.W. Clinical pharmacokinetics of systemic antifungal drugs. Clin. Pharmacokinet. 1983, 8, 17–42. [Google Scholar] [CrossRef]
- Schonebeck, J.; Polak, A.; Fernex, M.; Scholer, H.J. Pharmacokinetic studies on the oral antimycotic agent 5-fluorocytosine in individuals with normal and impaired kidney function. Chemotherapy 1973, 18, 321–336. [Google Scholar] [CrossRef]
- Block, E.R.; Bennett, J.E.; Livoti, L.G.; Klein, W.J., Jr.; MacGregor, R.R.; Henderson, L. Flucytosine and amphotericin B: Hemodialysis effects on the plasma concentration and clearance. Studies in man. Ann. Intern. Med. 1974, 80, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Vermes, A.; van Der Sijs, H.; Guchelaar, H.J. Flucytosine: Correlation between toxicity and pharmacokinetic parameters. Chemotherapy 2000, 46, 86–94. [Google Scholar] [CrossRef]
- Stamm, A.M.; Diasio, R.B.; Dismukes, W.E.; Shadomy, S.; Cloud, G.A.; Bowles, C.A.; Karam, G.H.; Espinel-Ingroff, A. Toxicity of amphotericin B plus flucytosine in 194 patients with cryptococcal meningitis. Am. J. Med. 1987, 83, 236–242. [Google Scholar] [CrossRef]
- Bennett, J.E.; Dismukes, W.E.; Duma, R.J.; Medoff, G.; Sande, M.A.; Gallis, H.; Leonard, J.; Fields, B.T.; Bradshaw, M.; Haywood, H.; et al. A comparison of amphotericin B alone and combined with flucytosine in the treatment of cryptoccal meningitis. N. Engl. J. Med. 1979, 301, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, C.A.; Frame, P.T. Bone marrow toxicity associated with 5-fluorocytosine therapy. Antimicrob. Agents Chemother. 1977, 11, 244–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nett, J.E.; Andes, D.R. Antifungal agents: Spectrum of activity, pharmacology, and clinical indications. Infect. Dis. Clin. 2016, 30, 51–83. [Google Scholar] [CrossRef]
- Whelan, W.L. The genetic basis of resistance to 5-fluorocytosine in Candida species and Cryptococcus neoformans. Crit. Rev. Microbiol. 1987, 15, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2014, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Kern, L.; de Montigny, J.; Lacroute, F.; Jund, R. Regulation of the pyrimidine salvage pathway by the FUR1 gene product of Saccharomyces cerevisiae. Curr. Genet. 1991, 19, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, A.R.; Dodgson, K.J.; Pujol, C.; Pfaller, M.A.; Soll, D.R. Clade-specific flucytosine resistance is due to a single nucleotide change in the FUR1 gene of Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 2223–2227. [Google Scholar] [CrossRef] [Green Version]
- Hope, W.W.; Tabernero, L.; Denning, D.W.; Anderson, M.J. Molecular mechanisms of primary resistance to flucytosine in Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 4377–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Khan, Z.U.; Joseph, L.; Asadzadeh, M.; Theyyathel, A. Genotypic heterogeneity and molecular basis of 5-flucytosine resistance among Candida dubliniensis isolates recovered from clinical specimens in Kuwait. Med. Mycol. 2012, 50, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Al Mosaid, A.; Sullivan, D.J.; Polacheck, I.; Shaheen, F.A.; Soliman, O.; Al Hedaithy, S.; Al Thawad, S.; Kabadaya, M.; Coleman, D.C. Novel 5-flucytosine-resistant clade of Candida dubliniensis from Saudi Arabia and Egypt identified by Cd25 fingerprinting. J. Clin. Microbiol. 2005, 43, 4026–4036. [Google Scholar] [CrossRef] [Green Version]
- McManus, B.A.; Moran, G.P.; Higgins, J.A.; Sullivan, D.J.; Coleman, D.C. A Ser29Leu substitution in the cytosine deaminase Fca1p is responsible for clade-specific flucytosine resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 2009, 53, 4678–4685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florent, M.; Noel, T.; Ruprich-Robert, G.; Da Silva, B.; Fitton-Ouhabi, V.; Chastin, C.; Papon, N.; Chapeland-Leclerc, F. Nonsense and missense mutations in FCY2 and FCY1 genes are responsible for flucytosine resistance and flucytosine-fluconazole cross-resistance in clinical isolates of Candida lusitaniae. Antimicrob. Agents Chemother. 2009, 53, 2982–2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, A.; Asner, S.A.; Trachsel, E.; Kelly, S.; Parker, J.; Sanglard, D. Comparative Genomics for the Elucidation of Multidrug Resistance in Candida lusitaniae. mBio 2019, 10, e02512-19. [Google Scholar] [CrossRef] [Green Version]
- Chapeland-Leclerc, F.; Hennequin, C.; Papon, N.; Noel, T.; Girard, A.; Socie, G.; Ribaud, P.; Lacroix, C. Acquisition of flucytosine, azole, and caspofungin resistance in Candida glabrata bloodstream isolates serially obtained from a hematopoietic stem cell transplant recipient. Antimicrob. Agents Chemother. 2010, 54, 1360–1362. [Google Scholar] [CrossRef] [Green Version]
- Edlind, T.D.; Katiyar, S.K. Mutational analysis of flucytosine resistance in Candida glabrata. Antimicrob. Agents Chemother. 2010, 54, 4733–4738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandeputte, P.; Pineau, L.; Larcher, G.; Noel, T.; Brethes, D.; Chabasse, D.; Bouchara, J.P. Molecular mechanisms of resistance to 5-fluorocytosine in laboratory mutants of Candida glabrata. Mycopathologia 2011, 171, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desnos-Ollivier, M.; Bretagne, S.; Bernede, C.; Robert, V.; Raoux, D.; Chachaty, E.; Forget, E.; Lacroix, C.; Dromer, F.; Yeasts, G. Clonal population of flucytosine-resistant Candida tropicalis from blood cultures, Paris, France. Emerg. Infect. Dis. 2008, 14, 557–565. [Google Scholar] [CrossRef]
- Chen, Y.N.; Lo, H.J.; Wu, C.C.; Ko, H.C.; Chang, T.P.; Yang, Y.L. Loss of heterozygosity of FCY2 leading to the development of flucytosine resistance in Candida tropicalis. Antimicrob. Agents Chemother. 2011, 55, 2506–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, J.; Abdolrasouli, A.; Farrer, R.A.; Cuomo, C.A.; Aanensen, D.M.; Armstrong-James, D.; Fisher, M.C.; Schelenz, S. Genomic epidemiology of the UK outbreak of the emerging human fungal pathogen Candida auris. Emerg. Microbes Infect. 2018, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Billmyre, R.B.; Applen Clancey, S.; Li, L.X.; Doering, T.L.; Heitman, J. 5-fluorocytosine resistance is associated with hypermutation and alterations in capsule biosynthesis in Cryptococcus. Nat. Commun. 2020, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jund, R.; Lacroute, F. Genetic and physiological aspects of resistance to 5-fluoropyrimidines in Saccharomyces cerevisiae. J. Bacteriol. 1970, 102, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevallier, M.R.; Jund, R.; Lacroute, F. Characterization of cytosine permeation in Saccharomyces cerevisiae. J. Bacteriol. 1975, 122, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Paluszynski, J.P.; Klassen, R.; Rohe, M.; Meinhardt, F. Various cytosine/adenine permease homologues are involved in the toxicity of 5-fluorocytosine in Saccharomyces cerevisiae. Yeast 2006, 23, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, Y.; Zhou, Y.; Zhao, Y.; Zhou, Y.; Cheng, J. Expression profiling of the response of Saccharomyces cerevisiae to 5-fluorocytosine using a DNA microarray. Int. J. Antimicrob. Agents 2002, 20, 444–450. [Google Scholar] [CrossRef]
- Cuenca-Estrella, M.; Diaz-Guerra, T.M.; Mellado, E.; Rodriguez-Tudela, J.L. Flucytosine primary resistance in Candida species and Cryptococcus neoformans. Eur. J. Clin. Microbiol. Infect. Dis. 2001, 20, 276–279. [Google Scholar] [CrossRef]
- Barchiesi, F.; Arzeni, D.; Caselli, F.; Scalise, G. Primary resistance to flucytosine among clinical isolates of Candida spp. J. Antimicrob. Chemother. 2000, 45, 408–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, P.; Dumas, C.; Joly, J. A study of 666 strains of Candida albicans: Correlation between serotype and susceptibility to 5-fluorocytosine. J. Infect. Dis. 1979, 139, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Drouhet, E.; Mercier-Soucy, L.; Montplaisir, S. [Sensitivity and resistance of pathogenic yeasts to 5-fluoropyrimidines. I.—Relation between the phenotypes of resistance to 5-fluorocytosine, the serotype of Candida albicans and the ecology of various species of Candida of human origin (author’s transl.)]. Ann. Microbiol. 1975, 126B, 25–39. [Google Scholar]
- Stiller, R.L.; Bennett, J.E.; Scholer, H.J.; Wall, M.; Polak, A.; Stevens, D.A. Susceptibility to 5-fluorocytosine and prevalence of serotype in 402 Candida albicans isolates from the United States. Antimicrob. Agents Chemother. 1982, 22, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Poulain, D.; Hopwood, V.; Vernes, A. Antigenic variability of Candida albicans. Crit. Rev. Microbiol. 1985, 12, 223–270. [Google Scholar] [CrossRef]
- Mercure, S.; Montplaisir, S.; Lemay, G. Correlation between the presence of a self-splicing intron in the 25S rDNA of C. albicans and strains susceptibility to 5-fluorocytosine. Nucleic Acids Res. 1993, 21, 6020–6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujol, C.; Pfaller, M.A.; Soll, D.R. Flucytosine resistance is restricted to a single genetic clade of Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 262–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, B.A.; Coleman, D.C.; Moran, G.; Pinjon, E.; Diogo, D.; Bougnoux, M.E.; Borecka-Melkusova, S.; Bujdakova, H.; Murphy, P.; d’Enfert, C.; et al. Multilocus sequence typing reveals that the population structure of Candida dubliniensis is significantly less divergent than that of Candida albicans. J. Clin. Microbiol. 2008, 46, 652–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, T.; Francois, F.; Paumard, P.; Chastin, C.; Brethes, D.; Villard, J. Flucytosine-fluconazole cross-resistance in purine-cytosine permease-deficient Candida lusitaniae clinical isolates: Indirect evidence of a fluconazole uptake transporter. Antimicrob. Agents Chemother. 2003, 47, 1275–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papon, N.; Noel, T.; Florent, M.; Gibot-Leclerc, S.; Jean, D.; Chastin, C.; Villard, J.; Chapeland-Leclerc, F. Molecular mechanism of flucytosine resistance in Candida lusitaniae: Contribution of the FCY2, FCY1, and FUR1 genes to 5-fluorouracil and fluconazole cross-resistance. Antimicrob. Agents Chemother. 2007, 51, 369–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, Y.; Chew, S.Y.; Than, L.T.L. Candida glabrata: Pathogenicity and Resistance Mechanisms for Adaptation and Survival. J. Fungi 2021, 7, 667. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Zhao, Y.; Perez, W.B.; Lockhart, S.R.; Sobel, J.D.; Farmakiotis, D.; Kontoyiannis, D.P.; Sanglard, D.; Taj-Aldeen, S.J.; Alexander, B.D.; et al. Prevalent mutator genotype identified in fungal pathogen Candida glabrata promotes multi-drug resistance. Nat. Commun. 2016, 7, 11128. [Google Scholar] [CrossRef] [Green Version]
- Normark, S.; Schonebeck, J. In vitro studies of 5-fluorocytosine resistance in Candida albicans and Torulopsis glabrata. Antimicrob. Agents Chemother. 1972, 2, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Fasoli, M.; Kerridge, D. Isolation and characterization of fluoropyrimidine-resistant mutants in two Candida species. Ann. N. Y. Acad. Sci. 1988, 544, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Steier, Z.; Vermitsky, J.P.; Toner, G.; Gygax, S.E.; Edlind, T.; Katiyar, S. Flucytosine antagonism of azole activity versus Candida glabrata: Role of transcription factor Pdr1 and multidrug transporter Cdr1. Antimicrob. Agents Chemother. 2013, 57, 5543–5547. [Google Scholar] [CrossRef] [Green Version]
- Lamoth, F.; Kontoyiannis, D.P. The Candida auris alert: Facts and perspectives. J. Infect. Dis. 2018, 217, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Frías-De-León, M.G.; Hernández-Castro, R.; Vite-Garín, T.; Arenas, R.; Bonifaz, A.; Castañón-Olivares, L.; Acosta-Altamirano, G.; Martínez-Herrera, E. Antifungal resistance in Candida auris: Molecular determinants. Antibiotics 2020, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene duplication associated with increased fluconazole tolerance in Candida auris cells of advanced generational age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, E.R.; Jennings, A.E.; Bennett, J.E. 5-fluorocytosine resistance in Cryptococcus neoformans. Antimicrob. Agents Chemother. 1973, 3, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanal Lamichhane, A.; Garraffo, H.M.; Cai, H.; Walter, P.J.; Kwon-Chung, K.J.; Chang, Y.C. A Novel Role of Fungal Type I Myosin in Regulating Membrane Properties and Its Association with d-Amino Acid Utilization in Cryptococcus gattii. mBio 2019, 10, e01867-19. [Google Scholar] [CrossRef] [Green Version]
- Vu, K.; Thompson, G.R., 3rd; Roe, C.C.; Sykes, J.E.; Dreibe, E.M.; Lockhart, S.R.; Meyer, W.; Engelthaler, D.M.; Gelli, A. Flucytosine resistance in Cryptococcus gattii is indirectly mediated by the FCY2-FCY1-FUR1 pathway. Med. Mycol. 2018, 56, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Song, M.H.; Lee, J.W.; Kim, M.S.; Yoon, J.K.; White, T.C.; Floyd, A.; Heitman, J.; Strain, A.K.; Nielsen, J.N.; Nielsen, K.; et al. A flucytosine-responsive Mbp1/Swi4-like protein, Mbs1, plays pleiotropic roles in antifungal drug resistance, stress response, and virulence of Cryptococcus neoformans. Eukaryot Cell 2012, 11, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.W.; Yang, D.H.; Maeng, S.; Lee, K.T.; So, Y.S.; Hong, J.; Choi, J.; Byun, H.J.; Kim, H.; Bang, S.; et al. Systematic functional profiling of transcription factor networks in Cryptococcus neoformans. Nat. Commun. 2015, 6, 6757. [Google Scholar] [CrossRef] [Green Version]
- Patterson, T.F.; Thompson, G.R., 3rd; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice guidelines for the diagnosis and management of aspergillosis: 2016 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef] [PubMed]
- Te Dorsthorst, D.T.; Mouton, J.W.; van den Beukel, C.J.; van der Lee, H.A.; Meis, J.F.; Verweij, P.E. Effect of pH on the in vitro activities of amphotericin B, itraconazole, and flucytosine against Aspergillus isolates. Antimicrob. Agents Chemother. 2004, 48, 3147–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Dorsthorst, D.T.; Verweij, P.E.; Meis, J.F.; Mouton, J.W. Efficacy and pharmacodynamics of flucytosine monotherapy in a nonneutropenic murine model of invasive aspergillosis. Antimicrob. Agents Chemother. 2005, 49, 4220–4226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, P.E.; Te Dorsthorst, D.T.; Janssen, W.H.; Meis, J.F.; Mouton, J.W. In vitro activities at pH 5.0 and pH 7.0 and in vivo efficacy of flucytosine against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2008, 52, 4483–4485. [Google Scholar] [CrossRef] [Green Version]
- Gsaller, F.; Furukawa, T.; Carr, P.D.; Rash, B.; Jochl, C.; Bertuzzi, M.; Bignell, E.M.; Bromley, M.J. Mechanistic Basis of pH-Dependent 5-Flucytosine Resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62, e02593-17. [Google Scholar] [CrossRef] [Green Version]
- Birstonas, L.; Dallemulle, A.; Lopez-Berges, M.S.; Jacobsen, I.D.; Offterdinger, M.; Abt, B.; Strassburger, M.; Bauer, I.; Schmidt, O.; Sarg, B.; et al. Multiplex Genetic Engineering Exploiting Pyrimidine Salvage Pathway-Based Endogenous Counterselectable Markers. mBio 2020, 11, e00230-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, R.; Katoh, T.; Nishioka, K. Isolation of dermatophytes from house dust on a medium containing gentamicin and flucytosine: Zur Isolierung von Dermatophyten aus Hausstaub auf einem Gentamicin-und Flucytosin-haltigen Medium. Mycoses 1995, 38, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Ansari, S.; Ilkit, M.; Rafati, H.; Hedayati, M.T.; Taghizadeh-Armaki, M.; Nasrollahi-Omran, A.; Tolooe, A.; Zhan, P.; Liao, W.; et al. In Vitro Antifungal Susceptibility Profiles of 12 Antifungal Drugs against 55 Trichophyton schoenleinii Isolates from Tinea Capitis Favosa Patients in Iran, Turkey, and China. Antimicrob. Agents Chemother. 2017, 61, e01753-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Bin, W.; Diao, Y.; Yang, J.; Liu, T.; Peng, J.; Jin, Q. Global transcriptional profiles of Trichophyton rubrum in response to Flucytosine. Sci. China Ser. C Life Sci. 2009, 52, 1173–1185. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Isolates | Gene | Gene Function | Mutation | Consequence | Reference |
|---|---|---|---|---|---|---|
| S. cerivisae | In vitro | FUR1 | Uracil phosphoribosyltransferase | R134S | Resistance to 5FU | [30] |
| C. albicans | Clinical | FUR1 | Uracil phosphoribosyltransferase | C301T ---> C101R | Disruption of dimerization of the enzyme | [31,32] |
| Clinical | FCA1 | Cytosine deaminase | G28D S29L | Defective enzyme | [32] | |
| FCY2 | Cytosine permease | A176G | Defective enzyme | [32] | ||
| C. dubliensis | Clinical | CdFCA1 | Cytosine deaminase | C86T ---> S29L | Disruption of the quaternary structure of the enzyme | [33,34,35] |
| C. lusitaniae | Clinical | FCY1 | Cytosine deaminase | T26C --->M9T | Defective enzyme | [36] |
| FCY2 | Cytosine permease | C505T--->E169Stop | Truncated cytosine permease | [36] | ||
| Clinical | MRR1 | Transcriptional activator | V668G | Upregulation of the multidrug transporter MFS7 | [37] | |
| C. glabrata | Clinical | FUR1 | Uracil phosphoribosyltransferase | G190D | Defective Fur1p | [38] |
| In vitro | FUR1 | Uracil phosphoribosyltransferase | I83K, D193G | Defective enzyme | [39,40] | |
| In vitro | FUR1 | Uracil phosphoribosyltransferase | ΔG73-V81 | Defective enzyme | [39,40] | |
| In vitro | FCY1 | Cytosine deaminase | A15D, G11Dand W148R | Defective enzyme | [39,40] | |
| In vitro | FCY2 | Cytosine permease | G246S and I384F | Defective enzyme | [39,40] | |
| FUR1 FCY1 FCY2J | Cytosine deaminase | G210D L136R T84L I384F | Defective enzyme | [39,40] | ||
| C. tropicalis | Clinical | URA3 | ODCase | K177E | Overabundance of UMP | [41] |
| FCY2 | Cytosine permease | G145T | Truncated protein | [42] | ||
| C. auris | Clinical | FUR1 | Uracil phosphoribosyltransferase | F211I | [43] | |
| C. deuterogattii | In vitro | FUR1 | Uracil phosphoribosyltransferase | 1134delT 1136delT 1440delA | Defective enzyme | [44] |
| FCY2 | Cytosine permease | W167Stop | Defective enzyme | |||
| UXS1 | UDP-glucuronate decarboxylase 1 | D306G Y217C 1520delT 1182insC | Accumulation of UDP glucuronic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delma, F.Z.; Al-Hatmi, A.M.S.; Brüggemann, R.J.M.; Melchers, W.J.G.; de Hoog, S.; Verweij, P.E.; Buil, J.B. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. J. Fungi 2021, 7, 909. https://doi.org/10.3390/jof7110909
Delma FZ, Al-Hatmi AMS, Brüggemann RJM, Melchers WJG, de Hoog S, Verweij PE, Buil JB. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. Journal of Fungi. 2021; 7(11):909. https://doi.org/10.3390/jof7110909
Chicago/Turabian StyleDelma, Fatima Zohra, Abdullah M. S. Al-Hatmi, Roger J. M. Brüggemann, Willem J. G. Melchers, Sybren de Hoog, Paul E. Verweij, and Jochem B. Buil. 2021. "Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi" Journal of Fungi 7, no. 11: 909. https://doi.org/10.3390/jof7110909
APA StyleDelma, F. Z., Al-Hatmi, A. M. S., Brüggemann, R. J. M., Melchers, W. J. G., de Hoog, S., Verweij, P. E., & Buil, J. B. (2021). Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. Journal of Fungi, 7(11), 909. https://doi.org/10.3390/jof7110909