
The Heat Shock Protein 60 and Pap1 Participate in the Sporothrix schenckii-Host Interaction


 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Peptidorhamnomannan Extraction and Fractioning
2.3. Peptidorhamnomannan Analysis by Liquid Chromatography-Mass Spectrometry
2.4. Peptidorhamnomannan Electrophoresis and Lectin Blot
2.5. Bioinformatic Analysis
2.6. RNA Isolation and Gene-Expression Analysis
2.7. Heterologous Expression of Sporothrix Schenckii HSP60 and PAP1 in Escherichia coli
2.8. Recombinant Protein Purification
2.9. Generation of Polyclonal Antibodies against Recombinant Hsp60 and Pap1
2.10. Analysis of Cell Adhesion to Extracellular Matrix Components
2.11. Infection Assays in Galleria mellonella
2.12. Analysis of the Galleria mellonella–Sporothrix schenckii Interaction
2.13. Statistical Analysis
3. Results
3.1. Polypeptide Composition of Peptidorhamnomannan
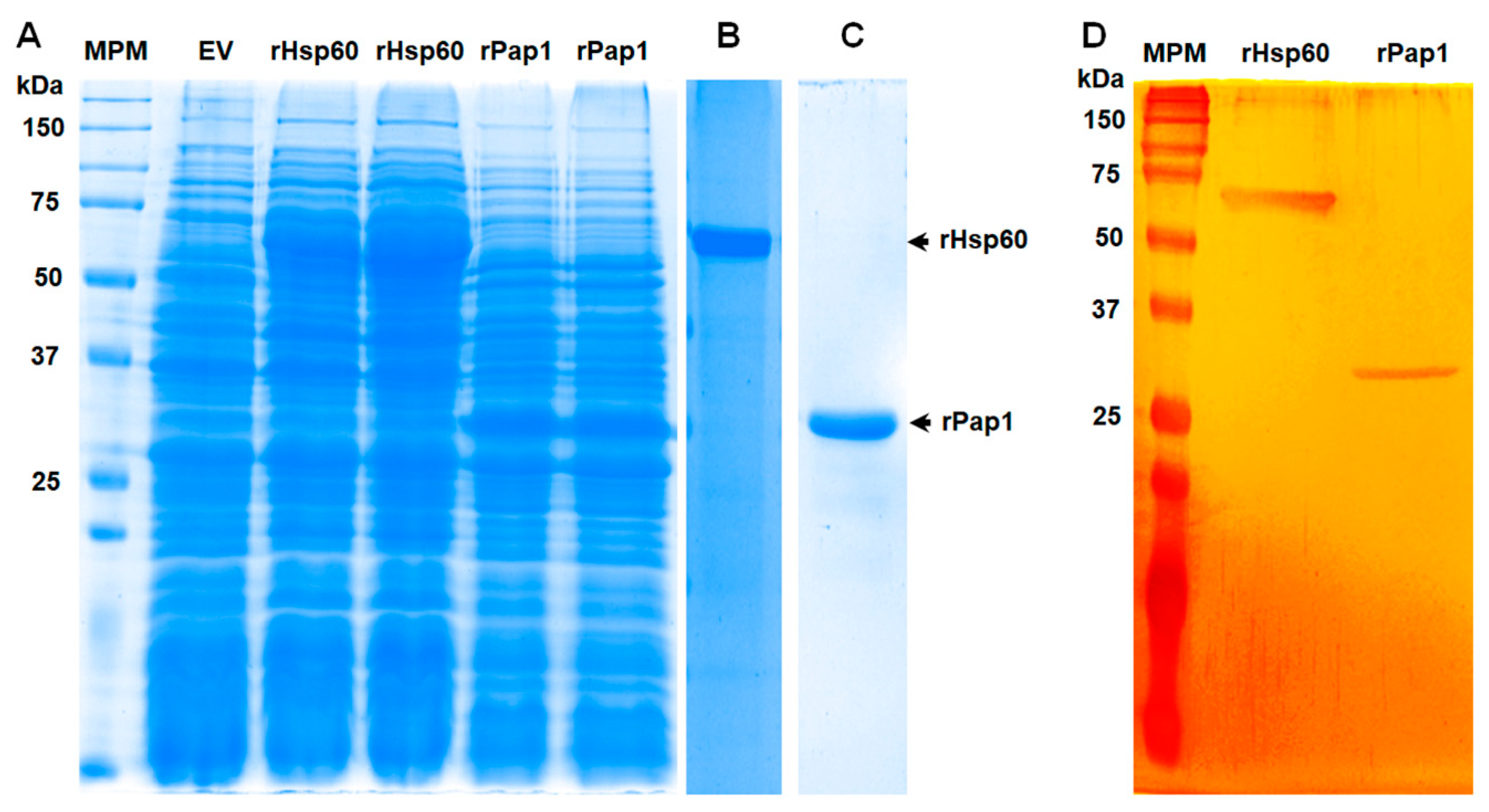
3.2. Production of Recombinant Hsp60 and Pap1
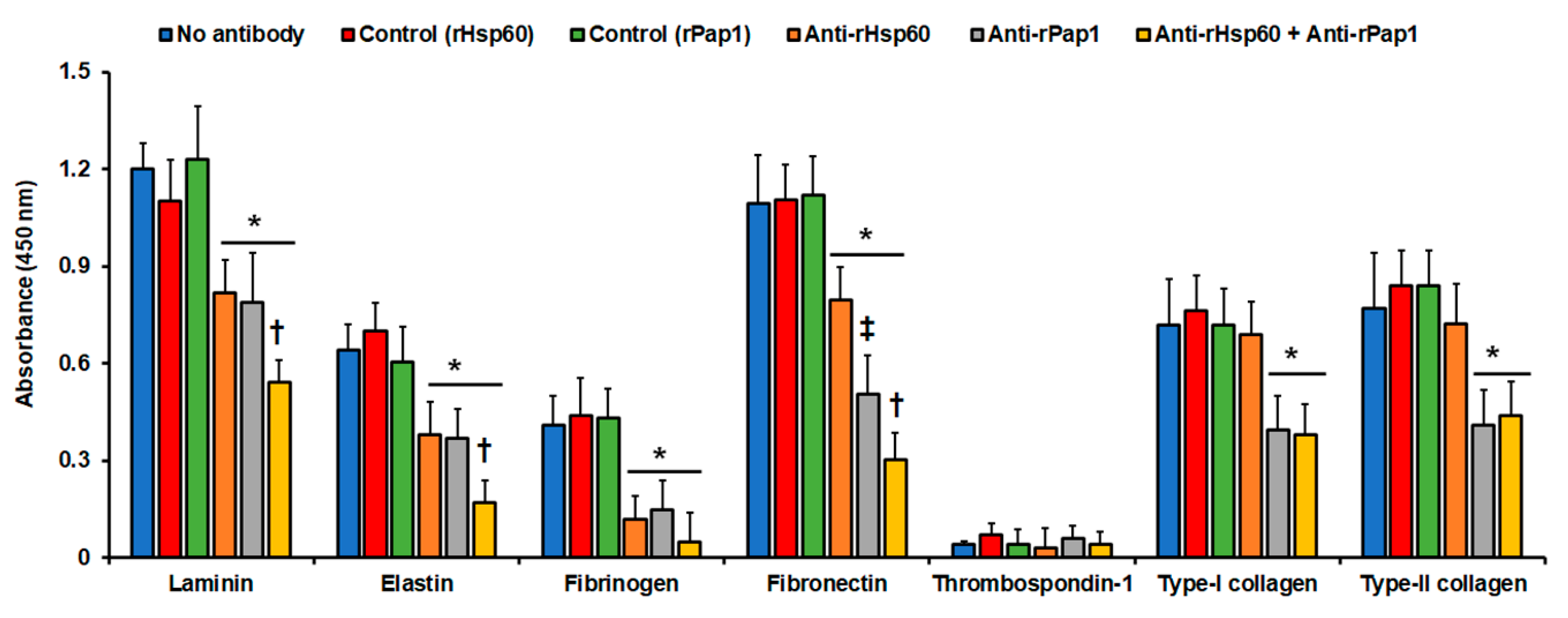
3.3. Hsp60 and Pap1 Participate in the Sporothrix schenckii Adhesion to Extracellular Matrix Proteins
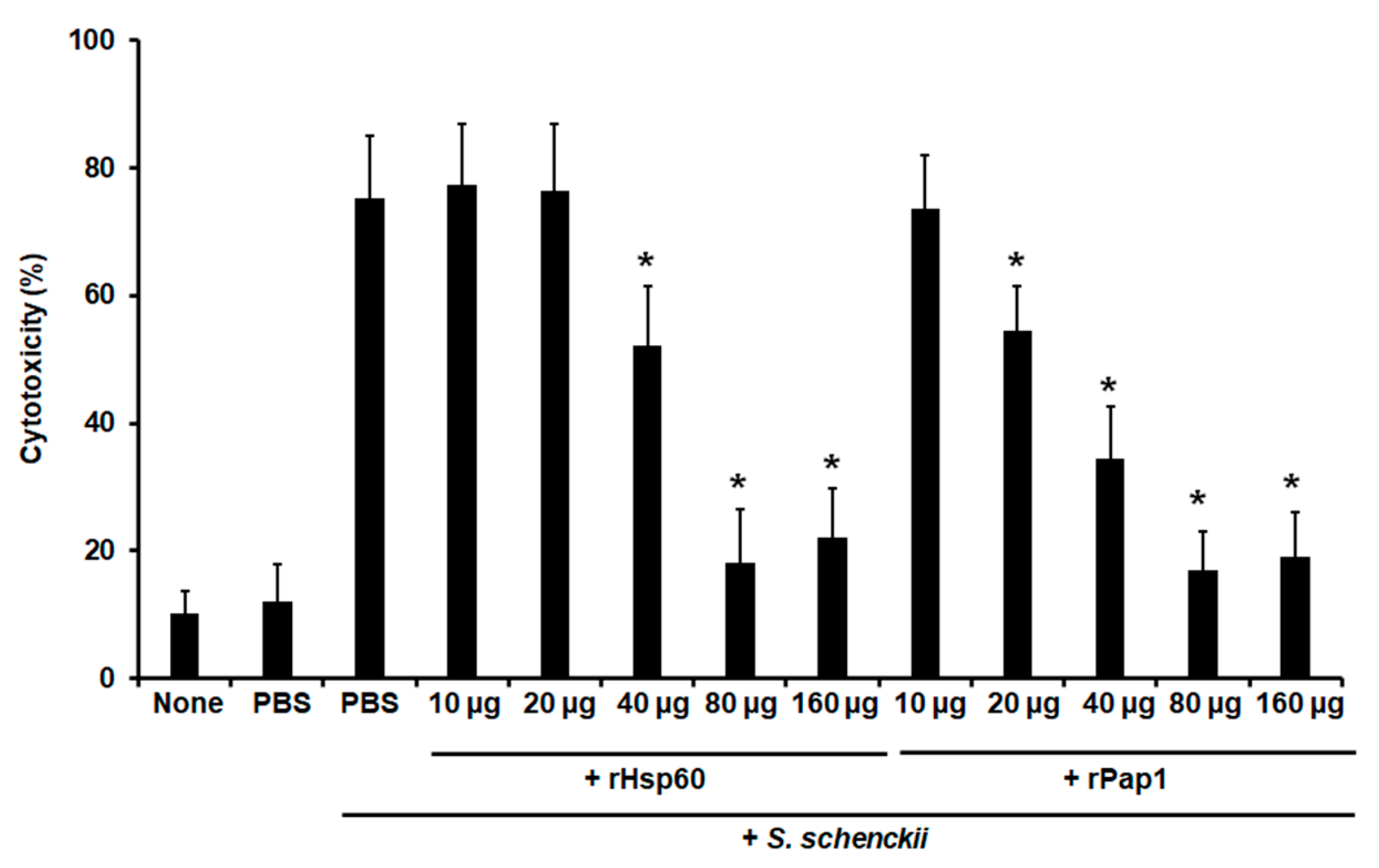
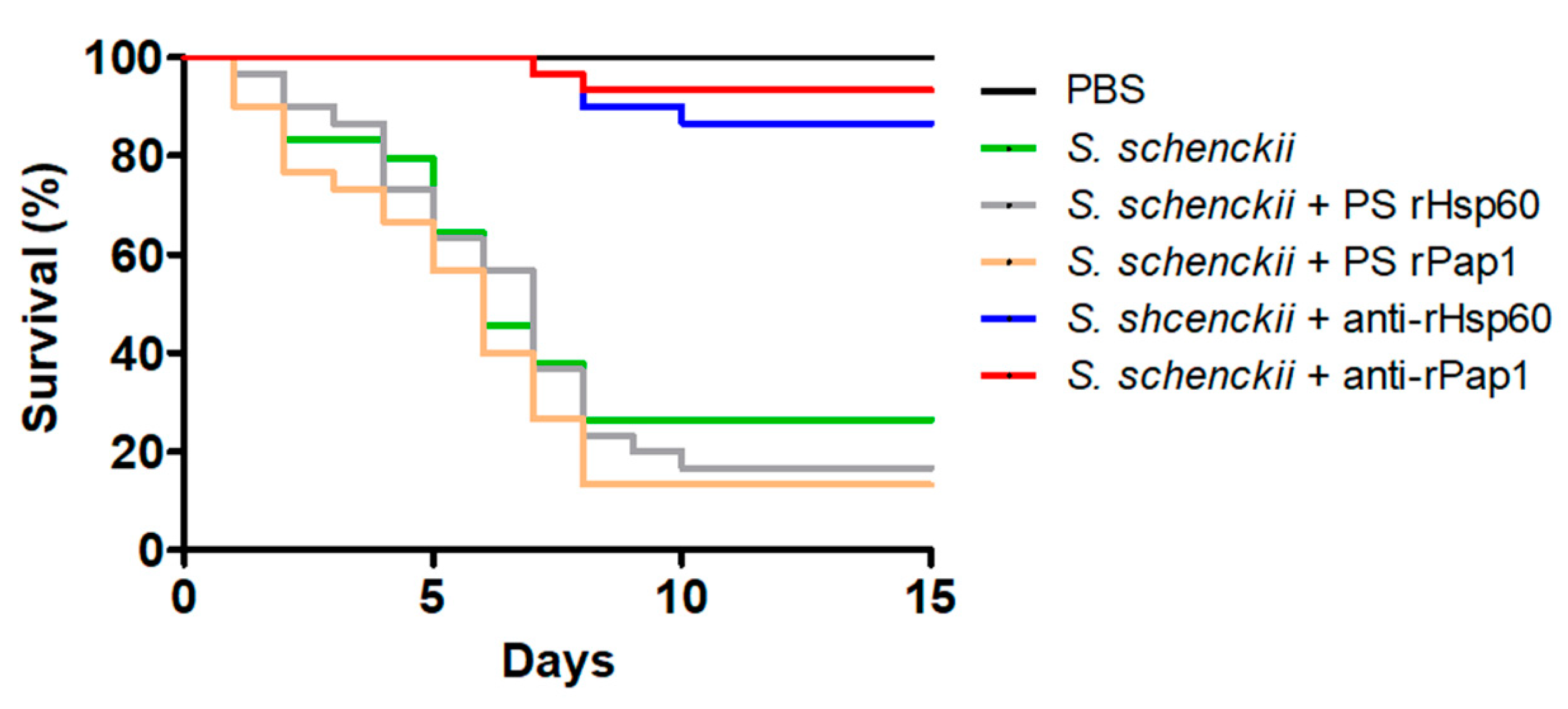
3.4. Hsp60 and Pap1 Contribute to Sporothrix schenckii Virulence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bonifaz, A.; Vázquez-González, D. Diagnosis and treatment of lymphocutaneous sporotrichosis: What are the options? Curr. Fungal Infect. Rep. 2013, 7, 252–259. [Google Scholar] [CrossRef]
- Bezerra, L.M.L.; Mora-Montes, H.M.; Zhang, Y.; Nino-Vega, G.; Rodrigues, A.M.; De Camargo, Z.P.; De Hoog, S. Sporotrichosis between 1898 and 2017: The evolution of knowledge on a changeable disease and on emerging etiological agents. Med. Mycol. 2018, 56, S126–S143. [Google Scholar] [CrossRef] [PubMed]
- De Beer, Z.; Duong, T.; Wingfield, M. The divorce of Sporothrix and Ophiostoma: Solution to a problematic relationship. Stud. Mycol. 2016, 83, 165–191. [Google Scholar] [CrossRef] [Green Version]
- López-Romero, E.; Reyes-Montes, M.D.R.; Pérez-Torres, A.; Ruiz-Baca, E.; Villagómez-Castro, J.C.; Mora-Montes, H.M.; Flores-Carreón, A.; Toriello, C. Sporothrix schenckii complex and sporotrichosis, an emerging health problem. Futur. Microbiol. 2011, 6, 85–102. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; De Hoog, G.S.; de Carmargo, Z.P. Sporothrix species causing outbreaks in animals and humans driven by animal—animal transmission. PLOS Pathog. 2016, 12, e1005638. [Google Scholar] [CrossRef]
- Etchecopaz, A.; Lanza, N.; Toscanini, M.A.; Devoto, T.B.; Pola, S.; Daneri, G.; Iovannitti, C.; Cuestas, M. Sporotrichosis caused by Sporothrix brasiliensis in Argentina: Case report, molecular identification and in vitro susceptibility pattern to antifungal drugs. J. Mycol. Med. 2020, 30, 100908. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N.E.; Clavijo-Giraldo, D.M.; Martínez-Duncker, I.; García-Carnero, L.C.; López-Ramírez, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Influences of the culturing media in the virulence and cell wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. J. Fungi 2020, 6, 323. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2014, 53, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Jiménez, D.F.; Pérez-García, L.A.; Martínez-Álvarez, J.A.; Mora-Montes, H.M. Role of the fungal cell wall in pathogenesis and antifungal resistance. Curr. Fungal Infect. Rep. 2012, 6, 275–282. [Google Scholar] [CrossRef]
- Hernández-Chávez, M.J.; Pérez-García, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Fungal strategies to evade the host immune recognition. J. Fungi 2017, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.; Latge, J.-P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 267–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Brown, G.; Kullberg, B.J.; Gow, N. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Genet. 2008, 6, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; De Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Previato, J.O.; Gorin, P.A.; Haskins, R.; Travassos, L.R. Soluble and insoluble glucans from different cell types of the human pathogen Sporothrix schenckii. Exp. Mycol. 1979, 3, 92–105. [Google Scholar] [CrossRef]
- Travassos, L.R.; Lloyd, K. Sporothrix schenckii and related species of Ceratocystis. Microbiol. Rev. 1980, 44, 683–721. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M.; Walker, L.A.; Nino-Vega, G.; Mora-Montes, H.M.; Neves, G.W.P.; Villalobos, H.; Barreto, L.; Garcia, K.; Franco, B.; Martínez-Álvarez, J.A.; et al. Cell walls of the dimorphic fungal pathogens Sporothrix schenckii and Sporothrix brasiliensis exhibit bilaminate structures and sloughing of extensive and intact layers. PLOS Negl. Trop. Dis. 2018, 12, e0006169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martinez-Duncker, I.; Bezerra, L.M.L.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Duno, H.L.; Barreto, L.A.; Alvarez-Aular, A.; Mora-Montes, H.M.; Lozoya-Pérez, N.E.; Franco, B.; Lopes-Bezerra, L.M.; Niño-Vega, G.A. Comparison of cell wall polysaccharide composition and structure between strains of Sporothrix schenckii and Sporothrix brasiliensis. Front. Microbiol. 2021, 12, 726958. [Google Scholar] [CrossRef]
- Lloyd, K.; Bitoon, M. Isolation and purification of a peptido-rhamnomannan from the yeast form of Sporothrix schenckii. Structural and immunochemical studies. J. Immunol. 1971, 107, 663–671. [Google Scholar]
- Bezerra, L.L.L.-B.L. Sporothrix schenckii Cell wall peptidorhamnomannans. Front. Microbiol. 2011, 2, 243. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Alves, L.M.; Fournet, B.; Degand, P. O-Glycosidically linked oligosaccharides from peptidorhamnomannans of Sporothrix schenckii. Glycoconj. J. 1992, 9, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Travassos, L.R.; de Sousa, W.; Mendonça-Previato, L.; Lloyd, K.O. Location and biochemical nature of surface components reacting with concanavalin A in different cell types of Sporothrix schenckii. Exp. Mycol. 1977, 1, 293–305. [Google Scholar] [CrossRef]
- Lima, O.C.; Bezerra, L.M.L. Identification of a concanavalin A-binding antigen of the cell surface of Sporothrix schenckii. Med. Mycol. 1997, 35, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, L.L.; Travassos, L.R.; Previato, J.O.; Previato, L.M. Novel antigenic determinants from peptidorhamnomannans of Sporothrix schenckii. Glycobiology 1994, 4, 281–288. [Google Scholar] [CrossRef]
- Penha, C.V.L.Y.; Bezerra, L.M.L. Concanavalin A-binding cell wall antigens of Sporothrix schenckii: A serological study. Med. Mycol. 2000, 38, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, K.O.; Travassos, L.R. Immunochemical studies on L-rhamno-D-mannans of Sporothrix schenckii and related fungi by use of rabbit and human antisera. Carbohydr. Res. 1975, 40, 89–97. [Google Scholar] [CrossRef]
- Bernardes-Engemann, A.R.; Costa, R.C.O.; Miguens, B.P.; Penha, C.V.L.; Neves, E.; Pereira, B.; Dias, C.M.P.; Mattos, M.; Gutierrez, M.C.; Schubach, A.; et al. Development of an enzyme-linked immunosorbent assay for the serodiagnosis of several clinical forms of sporotrichosis. Med. Mycol. 2005, 43, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Bernardes-Engemann, A.R.; Penha, C.V.L.Y.; Benvenuto, F.; Braga, J.U.; Barros, M.L.; Orofino-Costa, R.; Lopes-Bezerra, L.M. A comparative serological study of theSsCBF antigenic fraction isolated from three Sporothrix schenckii strains. Med. Mycol. 2009, 47, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.R.; de Sá, A.C.; Limongi, C.L.; Rozental, S.; Santos, A.L.; Barreto-Bergter, E. Involvement of peptidorhamnomannan in the interaction of Pseudallescheria boydii and HEp2 cells. Microbes Infect. 2004, 6, 1259–1267. [Google Scholar] [CrossRef]
- Tamez-Castrellón, A.K.; van der Beek, S.L.; López-Ramírez, L.A.; Martínez-Duncker, I.; Lozoya-Pérez, N.E.; van Sorge, N.M.; Mora-Montes, H.M. Disruption of protein rhamnosylation affects the Sporothrix schenckii-host interaction. Cell Surf. 2021, 7, 100058. [Google Scholar] [CrossRef]
- Figueiredo, C.C.; de Lima, O.C.; Carvalho, L.; Bezerra, L.M.L.; Morandi, V. The in vitro interaction of Sporothrix schenckii with human endothelial cells is modulated by cytokines and involves endothelial surface molecules. Microb. Pathog. 2004, 36, 177–188. [Google Scholar] [CrossRef]
- Gozalbo, D.; Gil-Navarro, I.; Azorín, I.; Renau-Piqueras, J.; Martiínez, J.P.; Gil, M.L. The Cell Wall-Associated Glyceraldehyde-3-Phosphate Dehydrogenase of Candida albicans Is Also a Fibronectin and Laminin Binding Protein. Infect. Immun. 1998, 66, 2052–2059. [Google Scholar] [CrossRef] [Green Version]
- Kozik, A.; Karkowska-Kuleta, J.; Zajac, D.; Bochenska, O.; Kedracka-Krok, S.; Jankowska, U.; Rapala-Kozik, M. Fibronectin-, vitronectin- and laminin-binding proteins at the cell walls of Candida parapsilosis and Candida tropicalis pathogenic yeasts. BMC Microbiol. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Karkowska-Kuleta, J.; Satała, D.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Moonlighting proteins are variably exposed at the cell surfaces of Candida glabrata, Candida parapsilosis and Candida tropicalis under certain growth conditions. BMC Microbiol. 2019, 19, 149. [Google Scholar] [CrossRef]
- Satala, D.; Karkowska-Kuleta, J.; Zelazna, A.; Rapala-Kozik, M.; Kozik, A. Moonlighting proteins at the candidal cell surface. Microorganisms 2020, 8, 1046. [Google Scholar] [CrossRef]
- Gomez, F.J.; Allendoerfer, R.; Deepe, G.S. Vaccination with recombinant heat shock protein 60 from Histoplasma capsulatum protects mice against pulmonary histoplasmosis. Infect. Immun. 1995, 63, 2587–2595. [Google Scholar] [CrossRef] [Green Version]
- Long, K.H.; Gomez, F.J.; Morris, R.E.; Newman, S.L. Identification of Heat Shock Protein 60 as the Ligand on Histoplasma capsulatum That Mediates Binding to CD18 Receptors on Human Macrophages. J. Immunol. 2003, 170, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, F.F.; Pitangui, N.S.; Peron, G.; Valim, C.X.R.; Mazucato, V.M.; Oliveira, A.F.; Da Silva, T.A.; Roque-Barreira, M.-C.; Panunto-Castelo, A. Expression of Hsp60 and its cell location in Paracoccidioides brasiliensis. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e29. [Google Scholar] [CrossRef]
- Silveira, C.P.; Piffer, A.C.; Kmetzsch, L.; Fonseca, F.L.; Soares, D.A.; Staats, C.; Rodrigues, M.; Schrank, A.; Vainstein, M. The heat shock protein (Hsp) 70 of Cryptococcus neoformans is associated with the fungal cell surface and influences the interaction between yeast and host cells. Fungal Genet. Biol. 2013, 60, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Amblee, V.; Jeffery, C.J. Physical features of intracellular proteins that moonlight on the cell surface. PLoS ONE 2015, 10, e0130575. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A.; Nosanchuk, J.D.; Williamson, P.; Rodrigues, M.L. Vesicular transport across the fungal cell wall. Trends Microbiol. 2009, 17, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Chávez, M.J.; González-Hernández, R.J.; Trujillo-Esquivel, J.E.; Hernández-Cervantes, A.; Mora-Montes, H.M. Chapter 9—The secretory pathway in the filamentous fungus Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 115–121. [Google Scholar]
- Teixeira, M.M.; de Almeida, L.G.; Kubitschek-Barreira, P.; Alves, F.L.; Kioshima, E.S.; Abadio, A.K.; Fernandes, L.; Derengowski, L.S.; Ferreira, K.S.; Souza, R.C.; et al. Comparative genomics of the major fungal agents of human and animal Sporotrichosis: Sporothrix schenckii and Sporothrix brasiliensis. BMC Genom. 2014, 15, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, K.O. Isolation, characterization, and partial structure of peptido galactomannans from the yeast form of Cladosporium werneckii. Biochemistry 1970, 9, 3446–3453. [Google Scholar] [CrossRef]
- Navarro-Arias, M.J.; Defosse, T.A.; Dementhon, K.; Csonka, K.; Mellado-Mojica, E.; Valério, A.D.; González-Hernández, R.J.; Courdavault, V.; Clastre, M.; Hernández, N.V.; et al. Disruption of protein mannosylation affects Candida guilliermondii cell wall, immune sensing, and virulence. Front. Microbiol. 2016, 7, 1951. [Google Scholar] [CrossRef]
- Díaz-Jiménez, D.F.; Mora-Montes, H.M.; Hernández-Cervantes, A.; Luna-Arias, J.P.; Gow, N.A.; Flores-Carreón, A. Biochemical characterization of recombinant Candida albicans mannosyltransferases Mnt1, Mnt2 and Mnt5 reveals new functions in O- and N-mannan biosynthesis. Biochem. Biophys. Res. Commun. 2012, 419, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Karasinski, J.; Wrobel, K.; Escobosa, A.R.C.; Konopka, A.; Bulska, E.; Wrobel, K. Allium cepa L. Response to Sodium Selenite (Se(IV)) Studied in Plant Roots by a LC-MS-Based Proteomic Approach. J. Agric. Food Chem. 2017, 65, 3995–4004. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Erdős, G.; Dosztányi, Z. Analyzing Protein Disorder with IUPred2A. Curr. Protoc. Bioinform. 2020, 70, e99. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Esquivel, E.; Martínez-Álvarez, J.A.; Clavijo-Giraldo, D.M.; Hernández, N.V.; Flores-Martínez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. The Sporothrix schenckii gene encoding for the ribosomal protein L6 has constitutive and stable expression and works as an endogenous control in gene expression analysis. Front. Microbiol. 2017, 8, 1676. [Google Scholar] [CrossRef]
- Robledo-Ortiz, C.I.; Flores-Carreón, A.; Hernández-Cervantes, A.; Álvarez-Vargas, A.; Lee, K.K.; Díaz-Jiménez, D.F.; Munro, C.A.; Cano-Canchola, C.; Mora-Montes, H.M. Isolation and functional characterization of Sporothrix schenckii ROT2, the encoding gene for the endoplasmic reticulum glucosidase II. Fungal Biol. 2012, 116, 910–918. [Google Scholar] [CrossRef]
- Trujillo-Esquivel, E.; Franco, B.; Flores-Martínez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. Purification of single-stranded cDNA based on RNA degradation treatment and adsorption chromatography. Nucleosides Nucleotides Nucleic Acids 2016, 35, 404–409. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hager, D.A.; Burgess, R.R. Elution of proteins from sodium dodecyl sulfate-polyacrylamide gels, removal of sodium dodecyl sulfate, and renaturation of enzymatic activity: Results with sigma subunit of Escherichia coli RNA polymerase, wheat germ DNA topoisomerase, and other enzymes. Anal. Biochem. 1980, 109, 76–86. [Google Scholar] [CrossRef]
- Martínez-Álvarez, J.A.; García-Carnero, L.C.; Kubitschek-Barreira, P.H.; Lozoya-Pérez, E.N.; Belmonte-Vázquez, J.L.; de Almeida, J.R.; Gómez-Infante, A.D.J.; Curty, N.; Villagómez-Castro, J.C.; Peña-Cabrera, E.; et al. Analysis of some immunogenic properties of the recombinant Sporothrix schenckii Gp70 expressed in Escherichia coli. Futur. Microbiol. 2019, 14, 397–410. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; Bader, O.; López-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Hube, B.; Gow, N.A.R.; Flores-Carreón, A. Kex2 protease converts the endoplasmic reticulum α1,2-mannosidase of Candida albicans into a soluble cytosolic form. Microbiology 2008, 154, 3782–3794. [Google Scholar] [CrossRef] [Green Version]
- Lima, O.; Figueiredo, C.; Pereira, B.; Coelho, M.; Morandi, V.; Lopes-Bezerra, L. Adhesion of the human pathogen Sporothrix schenckii to several extracellular matrix proteins. Braz. J. Med. Biol. Res. 1999, 32, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Clavijo-Giraldo, D.M.; Matínez-Alvarez, J.A.; Lopes-Bezerra, L.M.; Ponce-Noyola, P.; Franco, B.; Almeida, R.S.; Mora-Montes, H.M. Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis virulence in Galleria mellonella. J. Microbiol. Methods 2016, 122, 73–77. [Google Scholar] [CrossRef]
- Wu, G.; Yi, Y.; Lv, Y.; Li, M.; Wang, J.; Qiu, L. The lipopolysaccharide (LPS) of Photorhabdus luminescens TT01 can elicit dose- and time-dependent immune priming in Galleria mellonella larvae. J. Invertebr. Pathol. 2015, 127, 63–72. [Google Scholar] [CrossRef]
- Bidla, G.; Hauling, T.; Dushay, M.S.; Theopold, U. Activation of insect phenoloxidase after injury: Endogenous versus foreign elicitors. J. Innate Immun. 2009, 1, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; De Lucas, M.P.; Mesa-Arango, A.C.; Fusco-Almeida, A.M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M.J.; Zaragoza, O. Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef]
- Gómez-Gaviria, M.; Lozoya-Pérez, N.E.; Staniszewska, M.; Franco, B.; Niño-Vega, G.A.; Mora-Montes, H.M. Loss of Kex2 affects the Candida albicans cell wall and interaction with innate immune cells. J. Fungi 2020, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Horianopoulos, L.; Kronstad, J. Chaperone networks in fungal pathogens of humans. J. Fungi 2021, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.D.B.A.; Gomez, F.J.; Soares, C.M.D.A.; Deepe, G.S. Vaccination with heat shock protein 60 induces a protective immune response against experimental Paracoccidioides brasiliensis pulmonary infection. Infect. Immun. 2008, 76, 4214–4221. [Google Scholar] [CrossRef] [Green Version]
- Giosa, D.; Felice, M.R.; Giuffrè, L.; Cigliano, R.A.; Paytuví-Gallart, A.; Passo, C.L.; Barresi, C.; D’Alessandro, E.; Huang, H.; Criseo, G.; et al. Transcriptome-wide expression profiling of Sporothrix schenckii yeast and mycelial forms and the establishment of the Sporothrix Genome DataBase. Microb. Genom. 2020, 6, e000445. [Google Scholar] [CrossRef] [PubMed]
- Lozoya-Pérez, N.; Casas-Flores, S.; de Almeida, J.R.F.; Martínez-Álvarez, J.A.; López-Ramírez, L.A.; Jannuzzi, G.P.; Trujillo-Esquivel, E.; Estrada-Mata, E.; Almeida, S.R.; Franco, B.; et al. Silencing of OCH1 unveils the role of Sporothrix schenckii N-linked glycans during the host–fungus interaction. Infect. Drug Resist. 2018, 12, 67–85. [Google Scholar] [CrossRef] [Green Version]
- Lozoya-Pérez, N.E.; Casas-Flores, S.; Martínez-Álvarez, J.A.; López-Ramírez, L.A.; Lopes-Bezerra, L.M.; Franco, B.; Mora-Montes, H.M. Generation of Sporothrix schenckii mutants expressing the green fluorescent protein suitable for the study of host-fungus interactions. Fungal Biol. 2018, 122, 1023–1030. [Google Scholar] [CrossRef]
- García-Carnero, L.C.; Clavijo-Giraldo, D.M.; Gómez-Gaviria, M.; Lozoya-Pérez, N.E.; Tamez-Castrellón, A.K.; López-Ramírez, L.A.; Mora-Montes, H.M. Early Virulence Predictors during the Candida Species–Galleria mellonella Interaction. J. Fungi 2020, 6, 152. [Google Scholar] [CrossRef]
- Cohen, M.J.; Chirico, W.J.; Lipke, P.N. Through the back door: Unconventional protein secretion. Cell Surf. 2020, 6, 100045. [Google Scholar] [CrossRef]
- Gancedo, C.; Flores, C.-L.; Gancedo, J.M. The Expanding Landscape of Moonlighting Proteins in Yeasts. Microbiol. Mol. Biol. Rev. 2016, 80, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.N.; Solis, N.V.; Phan, Q.T.; Bajwa, J.S.; Kashleva, H.; Thompson, A.; Liu, Y.; Dongari-Bagtzoglou, A.; Edgerton, M.; Filler, S.G. Host Cell Invasion and Virulence Mediated by Candida albicans Ssa1. PLOS Pathog. 2010, 6, e1001181. [Google Scholar] [CrossRef] [Green Version]
- Jong, A.Y.; Chen, S.H.M.; Stins, M.F.; Kim, K.S.; Tuan, T.-L.; Huang, S.-H. Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J. Med. Microbiol. 2003, 52, 615–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Hoffmann, R.; Skerka, C.; Zipfel, P.F. Glycerol-3-phosphate dehydrogenase 2 is a novel factor H–, factor H–like protein 1–, and plasminogen-binding surface protein of Candida albicans. J. Infect. Dis. 2012, 207, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Crowe, J.D.; Sievwright, I.K.; Auld, G.C.; Moore, N.R.; Gow, N.A.R.; Booth, N.A. Candida albicans binds human plasminogen: Identification of eight plasminogen-binding proteins. Mol. Microbiol. 2003, 47, 1637–1651. [Google Scholar] [CrossRef] [PubMed]
- Poltermann, S.; Kunert, A.; von der Heide, M.; Eck, R.; Hartmann, A.; Zipfel, P.F. Gpm1p Is a Factor H-, FHL-1-, and Plasminogen-binding Surface Protein of Candida albicans. J. Biol. Chem. 2007, 282, 37537–37544. [Google Scholar] [CrossRef] [Green Version]
- Stie, J.; Bruni, G.; Fox, D. Surface-associated plasminogen binding of Cryptococcus neoformans promotes extracellular matrix invasion. PLoS ONE 2009, 4, e5780. [Google Scholar] [CrossRef] [Green Version]
- Félix-Contreras, C.; Alba-Fierro, C.A.; Ríos-Castro, E.; Luna-Martínez, F.; Cuéllar-Cruz, M.; Ruiz-Baca, E. Proteomic analysis of Sporothrix schenckii cell wall reveals proteins involved in oxidative stress response induced by menadione. Microb. Pathog. 2020, 141, 103987. [Google Scholar] [CrossRef]
- Ruiz-Baca, E.; Leyva-Sánchez, H.; Calderón-Barraza, B.; Esquivel-Naranjo, U.; López-Romero, E.; López-Rodríguez, A.; Cuéllar-Cruz, M. Identification of proteins in Sporothrix schenckii sensu stricto in response to oxidative stress induced by hydrogen peroxide. Rev. Iberoam. Micol. 2019, 36, 17–23. [Google Scholar] [CrossRef]
- Tiwari, S.; Thakur, R.; Shankar, J. Role of heat-shock proteins in cellular function and in the biology of fungi. Biotechnol. Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zügel, U.; Kaufmann, S.H.E. Role of heat shock proteins in protection from and pathogenesis of infectious diseases. Clin. Microbiol. Rev. 1999, 12, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Cleare, L.G.; Zamith-Miranda, D.; Nosanchuk, J.D. Heat shock proteins in Histoplasma and Paracoccidioides. Clin. Vaccine Immunol. 2017, 24, e00221-17. [Google Scholar] [CrossRef] [Green Version]
- Izacc, S.M.S.; Gomez, F.J.; Jesuino, R.S.A.; Fonseca, C.A.; Felipe, M.S.S.; Deepe, G.S.; Soares, C.M.A. Molecular cloning, characterization and expression of the heat shock protein 60 gene from the human pathogenic fungus Paracoccidioides brasiliensis. Med. Mycol. 2001, 39, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Thomaz, L.; Nosanchuk, J.D.; Rossi, D.C.; Travassos, L.R.; Taborda, C.P. Monoclonal antibodies to heat shock protein 60 induce a protective immune response against experimental Paracoccidioides lutzii. Microbes Infect. 2014, 16, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Raggam, R.B.; Salzer, H.J.F.; Marth, E.; Heiling, B.; Paulitsch, A.H.; Buzina, W. Molecular detection and characterisation of fungal heat shock protein 60. Mycoses 2011, 54, e394–e399. [Google Scholar] [CrossRef] [PubMed]
- Wallmann, A.; Kesten, C. Common functions of disordered proteins across evolutionary distant organisms. Int. J. Mol. Sci. 2020, 21, 2105. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.; Sullivan, W.J.S., Jr.; Radivojac, P.; Dunker, A.K.; Uversky, V.N. Intrinsic disorder in pathogenic and non-pathogenic microbes: Discovering and analyzing the unfoldomes of early-branching eukaryotes. Mol. BioSyst. 2008, 4, 328–340. [Google Scholar] [CrossRef]
- Jeffery, C.J. Why study moonlighting proteins? Front. Genet. 2015, 6, 211. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, F.F.; De Oliveira, L.L.; Landgraf, T.; Peron, G.; Costa, M.V.; Coelho-Castelo, A.A.M.; Bonato, V.; Barreira, M.C.R.; Panunto-Castelo, A. detrimental effect of fungal 60-kda heat shock protein on experimental Paracoccidioides brasiliensis infection. PLoS ONE 2016, 11, e0162486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Baca, E.; Toriello, C.; Pérez-Torres, A.; Sabanero-López, M.; Villagómez-Castro, J.C.; López-Romero, E. Isolation and some properties of a glycoprotein of 70 kDa (Gp70) from the cell wall of Sporothrix schenckii involved in fungal adherence to dermal extracellular matrix. Med. Mycol. 2009, 47, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.A.C.; De Castro, R.A.; Nascimento, R.C.; Tronchin, G.; Torres, A.P.; Lazéra, M.; Almeida, S.; Bouchara, J.-P.; Penha, C.V.L.Y.; Bezerra, L.M.L. Cell surface expression of adhesins for fibronectin correlates with virulence in Sporothrix schenckii. Microbiology 2009, 155, 3730–3738. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.A.; Kubitschek-Barreira, P.H.; Teixeira, P.A.C.; Sanches, G.; Teixeira, M.M.; Quintella, L.P.; Almeida, S.R.; Costa, R.O.; de Carmargo, Z.P.; Felipe, M.S.S.; et al. Differences in cell morphometry, cell wall topography and Gp70 expression correlate with the virulence of Sporothrix brasiliensis clinical isolates. PLoS ONE 2013, 8, e75656. [Google Scholar] [CrossRef] [Green Version]
- Lima, O.C.; Bouchara, J.-P.; Renier, G.; Marot-Leblond, A.; Chabasse, D.; Lopes-Bezerra, L.M. Immunofluorescence and flow cytometry analysis of fibronectin and laminin binding to Sporothrix schenckii yeast cells and conidia. Microb. Pathog. 2004, 37, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Lima, O.C.; Figueiredo, C.C.; Previato, J.O.; Mendonça-Previato, L.; Morandi, V.; Bezerra, L.M.L. Involvement of fungal cell wall components in adhesion of Sporothrix schenckii to human fibronectin. Infect. Immun. 2001, 69, 6874–6880. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Garduño, J.; Rodriguez, M.D.C.; Hernández-Martínez, S.; Martinez-Barnetche, J.; Alvarado-Delgado, A.; Izquierdo, J.; Herrera-Ortiz, A.; Moreno-García, M.; Velazquez-Meza, M.E.; Valverde, V.; et al. Plasmodium berghei induced priming in Anopheles albimanus independently of bacterial co-infection. Dev. Comp. Immunol. 2015, 52, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Xu, L.; Yi, Y. Galleria mellonella larvae are capable of sensing the extent of priming agent and mounting proportionatal cellular and humoral immune responses. Immunol. Lett. 2016, 174, 45–52. [Google Scholar] [CrossRef]
- Taszłow, P.; Vertyporokh, L.; Wojda, I. Humoral immune response of Galleria mellonella after repeated infection with Bacillus thuringiensis. J. Invertebr. Pathol. 2017, 149, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Jiang, R.; Wang, Y.; Zhu, M.; Zhang, X.; Dong, S.; Shi, H.; Wang, L. Recombinant phage elicits protective immune response against systemic S. globosa infection in mouse model. Sci. Rep. 2017, 7, 42024. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, J.R.F.; Jannuzzi, G.P.; Kaihami, G.H.; Breda, L.; Ferreira, K.S.; De Almeida, S.R. An immunoproteomic approach revealing peptides from Sporothrix brasiliensis that induce a cellular immune response in subcutaneous sporotrichosis. Sci. Rep. 2018, 8, 4192. [Google Scholar] [CrossRef]
- Guimaraes, A.; Frases, S.; Gomez, F.J.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Monoclonal antibodies to heat shock protein 60 alter the pathogenesis of Histoplasma capsulatum. Infect. Immun. 2009, 77, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- D’Áurea Moura, Á.N.; Garforth, S.J.; da Silva, L.B.R.; Woodley, D.; Barbalho, F.V.; Quiroz, J.A.; Almo, S.C.; Lai, J.R.; Nosanchuk, J.D.; Taborda, C.P. Generation and activity evaluation of a mouse-human immunoglobulin g1 chimeric antibody to Histoplasma capsulatum HSP60. Preprints 2021, 2021, 040101. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Gene | Coverage (%) | q-Value | Andromeda Score |
|---|---|---|---|---|
| Chaperonin GroEL-like protein/heat shock protein 60 | SPSK_01586 | 48.9 | 0 | 323 |
| Heat shock 70 kDa protein 1/8 | SPSK_08625 | 47.6 | 0 | 323 |
| Uncharacterized protein | SPSK_04236 | 22.2 | 0 | 323 |
| Uncharacterized protein | SPSK_00848 | 21.3 | 0 | 323 |
| Uncharacterized protein | SPSK_05930 | 19.8 | 0 | 323 |
| Glyceraldehyde-3-phosphate dehydrogenase | SPSK_00294 | 38.3 | 0 | 322 |
| ATP synthase subunit beta | SPSK_01537 | 54.7 | 0 | 307 |
| Elongation factor 1-alpha | SPSK_06026 | 33.5 | 0 | 291 |
| GPI-anchored cell wall beta-1,3-endoglucanase EglC | SPSK_01694 | 21.2 | 0 | 260 |
| Aldehyde dehydrogenase (NAD+) | SPSK_00262 | 28.4 | 0 | 244 |
| Large subunit ribosomal protein LP2 | SPSK_03751 | 34.2 | 0 | 218 |
| Uncharacterized protein | SPSK_01041 | 23.4 | 0 | 217 |
| AMPK1_CBM domain-containing protein | SPSK_08400 | 27.6 | 0 | 197 |
| Molecular chaperone DnaK | SPSK_03148 | 22.9 | 0 | 187 |
| Glucose-repressible protein | SPSK_01322 | 64.8 | 0 | 182 |
| Peptidyl-prolyl cis-trans isomerase | SPSK_01963 | 24.2 | 0 | 172 |
| Uncharacterized protein | SPSK_02764 | 50.3 | 0 | 158 |
| Large subunit ribosomal protein LP1 | SPSK_05488 | 43.6 | 0 | 149 |
| Sphingolipid long chain base-responsive protein | SPSK_05417 | 21.7 | 0 | 116 |
| Acetyltransferase component of pyruvate dehydrogenase complex | SPSK_07465 | 14.7 | 0 | 115 |
| Mismatched base pair and cruciform DNA recognition protein | SPSK_05604 | 28.3 | 0 | 113 |
| Aconitate hydratase, mitochondrial | SPSK_00414 | 13.8 | 0 | 110 |
| Uncharacterized protein | SPSK_06559 | 32.6 | 0 | 110 |
| Cytochrome c oxidase subunit 5b | SPSK_03566 | 29.4 | 0 | 110 |
| 2-phosphoglycerate dehydratase | SPSK_03292 | 20.5 | 0 | 106 |
| Actin beta/gamma 1 | SPSK_00108 | 15 | 0 | 104 |
| Inoculated with | Hemocytes (×106) mL−1 a | Phenoloxidase b | Melanin c |
|---|---|---|---|
| None | 3.3 ± 0.4 | 0.5 ± 0.3 | 0.7 ± 0.4 |
| PBS d | 3.1 ± 0.7 | 0.8 ± 0.5 | 1.1 ± 0.2 |
| 10 µg rHsp60 | 2.9 ± 0.6 | 0.7 ± 0.4 | 1.3 ± 0.5 |
| 20 µg rHsp60 | 3.2 ± 0.5 | 1.0 ± 0.3 | 1.0 ± 0.3 |
| 40 µg rHsp60 | 6.9 ± 0.8 * | 1.9 ± 0.6 * | 2.8 ± 0.4 * |
| 80 µg rHsp60 | 9.8 ± 0.4 * † | 2.7 ± 0.6 * † | 4.2 ± 0.8 * † |
| 160 µg rHsp60 | 10.1 ± 0.7 * † | 3.1 ± 0.7 * † | 3.8 ± 0.4 * † |
| 10 µg rPap1 | 3.3 ± 0.8 | 0.9 ± 0.2 | 1.1 ± 0.2 |
| 20 µg rPap1 | 5.3 ± 0.6 * | 1.6 ± 0.4 * | 2.1 ± 0.4 * |
| 40 µg rPap1 | 8.2 ± 0.5 * † | 2.4 ± 0.4 * † | 3.2 ± 0.4 * † |
| 80 µg rPap1 | 10.2 ± 0.6 * † | 3.4 ± 0.3 * † | 4.3 ± 0.6 * † |
| 160 µg rPap1 | 9.8 ± 0.6 * † | 3.3 ± 0.5 * † | 4.0 ± 0.5 * † |
| Inoculated with | Colony-Forming Units (×105) a | Hemocytes (×106) mL−1 a | Phenoloxidase b | Melanin c |
|---|---|---|---|---|
| None | 0.0 ± 0.0 | 3.6 ± 0.2 | 0.4 ± 0.4 | 0.5 ± 0.3 |
| PBS d | 0.0 ± 0.0 | 3.3 ± 0.3 | 0.9 ± 0.3 | 1.2 ± 0.4 |
| PBS + S. schenckii | 2.4 ± 0.4 | 8.0 ± 0.6 | 3.2 ± 0.6 | 3.5 ± 0.6 |
| 10 µg rHsp60 + S. schenckii | 2.7 ± 0.3 | 7.6 ± 0.5 | 3.7 ± 0.3 | 3.3 ± 0.3 |
| 20 µg rHsp60 + S. schenckii | 2.3 ± 0.4 | 8.1 ± 0.3 | 3.2 ± 0.5 | 3.3 ± 0.3 |
| 40 µg rHsp60 S. schenckii | 1.3 ± 0.2 * | 7.2 ± 0.5 | 2.6 ± 0.5 | 3.0 ± 0.2 |
| 80 µg rHsp60 + S. schenckii | 0.4 ± 0.2 * | 10.5 ± 0.5 * | 4.4 ± 0.4 * | 4.4 ± 0.3 * |
| 160 µg rHsp6 + S. schenckii | 0.2 ± 0.2 * | 10.3 ± 0.3 * | 4.6 ± 0.2 * | 4.6 ± 0.5 * |
| 10 µg rPap1 + S. schenckii | 2.7 ± 0.5 | 7.3 ± 0.7 | 2.9 ± 0.5 | 3.1 ± 0.5 |
| 20 µg rPap1 + S. schenckii | 1.9 ± 0.3 * | 7.7 ± 0.6 | 3.3 ± 0.3 | 3.2 ± 0.5 |
| 40 µg rPap1 + S. schenckii | 1.1 ± 0.2 * | 10.2 ± 0.3 * | 4.4 ± 0.3 * | 4.1 ± 0.3 * |
| 80 µg rPap1 + S. schenckii | 0.3 ± 0.2 * | 10.6 ± 0.4 * | 4.6 ± 0.5 * | 4.7 ± 0.5 * |
| 160 µg rPap1 + S. schenckii | 0.2 ± 0.1 * | 10.8 ± 0.8 * | 4.5 ± 0.3 * | 4.5 ± 0.4 * |
| Inoculated with | Colony-Forming Units (×105) a | Hemocytes (×106) mL−1 a | Phenoloxidase b | Melanin c |
|---|---|---|---|---|
| None | 0.0 ± 0.0 | 3.8 ± 0.4 | 0.3 ± 0.2 | 0.4 ± 0.2 |
| PBS d | 0.0 ± 0.0 | 3.9 ± 0.5 | 0.7 ± 0.5 | 0.9 ± 0.6 |
| S. schenckii | 2.3 ± 0.5 | 8.0 ± 0.3 | 2.9 ± 0.4 | 3.9 ± 0.3 |
| S. schenckii + PS rHsp60 | 2.8 ± 0.6 | 8.2 ± 0.4 | 3.1 ± 0.5 | 3.7 ± 0.6 |
| S. schenckii + PS rPap1 | 2.6 ± 0.6 | 7.8 ± 0.3 | 3.4 ± 0.3 | 3.9 ± 0.5 |
| S. schenckii + anti-rHsp60 | 0.9 ± 0.5 * | 4.3 ± 0.6 * | 0.7 ± 0.2 * | 0.8 ± 0.5 * |
| S. schenckii + anti-rPap1 | 0.5 ± 0.4 * | 3.5 ± 0.7 * | 0.6 ± 0.6 * | 0.5 ± 0.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Carnero, L.C.; Salinas-Marín, R.; Lozoya-Pérez, N.E.; Wrobel, K.; Wrobel, K.; Martínez-Duncker, I.; Niño-Vega, G.A.; Mora-Montes, H.M. The Heat Shock Protein 60 and Pap1 Participate in the Sporothrix schenckii-Host Interaction. J. Fungi 2021, 7, 960. https://doi.org/10.3390/jof7110960
García-Carnero LC, Salinas-Marín R, Lozoya-Pérez NE, Wrobel K, Wrobel K, Martínez-Duncker I, Niño-Vega GA, Mora-Montes HM. The Heat Shock Protein 60 and Pap1 Participate in the Sporothrix schenckii-Host Interaction. Journal of Fungi. 2021; 7(11):960. https://doi.org/10.3390/jof7110960
Chicago/Turabian StyleGarcía-Carnero, Laura C., Roberta Salinas-Marín, Nancy E. Lozoya-Pérez, Katarzyna Wrobel, Kazimierz Wrobel, Iván Martínez-Duncker, Gustavo A. Niño-Vega, and Héctor M. Mora-Montes. 2021. "The Heat Shock Protein 60 and Pap1 Participate in the Sporothrix schenckii-Host Interaction" Journal of Fungi 7, no. 11: 960. https://doi.org/10.3390/jof7110960
APA StyleGarcía-Carnero, L. C., Salinas-Marín, R., Lozoya-Pérez, N. E., Wrobel, K., Wrobel, K., Martínez-Duncker, I., Niño-Vega, G. A., & Mora-Montes, H. M. (2021). The Heat Shock Protein 60 and Pap1 Participate in the Sporothrix schenckii-Host Interaction. Journal of Fungi, 7(11), 960. https://doi.org/10.3390/jof7110960