
Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives


Abstract
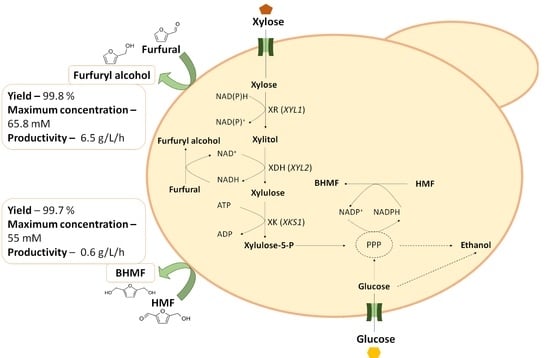
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Furfural and HMF-Reductive Enzymatic Activities
2.3. Whole-Cell Bioconversion of Furfural and HMF
2.4. Cell Viability Assay
2.5. Analytical Procedures
3. Results
3.1. Screening of Different K. marxianus Strains for Xylose Consumption and Tolerance to Lignocellulosic-Derived Inhibitors
3.2. Evaluation of K. marxianus Strains S9 and CBS6556 Tolerance in the Presence of Higher Inhibitory Loads
3.3. HMF and Furfural Reducing Activity for K. marxianus Strains S9 and CBS6556
Assessment of Furfural and HMF Derivatives Produced by K. marxianus Strains S9 and CBS6556
3.4. Whole-Cell Bioconversion of Furfural and HMF by K. marxianus Strain S9
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.M.; Morrissey, J.P. Kluyveromyces marxianus: A yeast emerging from its sister’s shadow. Fungal Biol. Rev. 2010, 24, 17–26. [Google Scholar] [CrossRef]
- Madeira, J.V., Jr.; Gombert, A.K. Towards high-temperature fuel ethanol production using Kluyveromyces marxianus: On the search for plug-in strains for the Brazilian sugarcane-based biorefinery. Biomass Bioenergy 2018, 119, 217–228. [Google Scholar] [CrossRef]
- Raimondi, S.; Zanni, E.; Amaretti, A.; Palleschi, C.; Uccelletti, D.; Rossi, M. Thermal adaptability of Kluyveromyces marxianus in recombinant protein production. Microb. Cell Factories 2013, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sa-Correia, I.; Domingues, L. Molecular and physiological basis of Saccharomyces cerevisiae tolerance to adverse lignocellulose-based process conditions. Appl. Microbiol. Biotechnol. 2018, 103, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Oliva, J.; Ballesteros, I.; Negro, M.J.; Manzanares, P.; Cabañas, A.; Ballesteros, M. Effect of Binary Combinations of Selected Toxic Compounds on Growth and Fermentation of Kluyveromyces marxianus. Biotechnol. Prog. 2004, 20, 715–720. [Google Scholar] [CrossRef]
- Nitiyon, S.; Keo-Oudone, C.; Murata, M.; Lertwattanasakul, N.; Limtong, S.; Kosaka, T.; Yamada, M. Efficient conversion of xylose to ethanol by stress-tolerant Kluyveromyces marxianus BUNL-21. SpringerPlus 2016, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, V.; Radecka, D.; Aerts, G.; Verstrepen, K.J.; Lievens, B.; Thevelein, J.M. Phenotypic landscape of non-conventional yeast species for different stress tolerance traits desirable in bioethanol fermentation. Biotechnol. Biofuels 2017, 10, 1–19. [Google Scholar] [CrossRef]
- Flores, G.; Biotechnology, C.I. Physiological Response to Furan Derivatives Stress by Kluyveromyces marxianus SLP1 in Ethanol Production. Rev. Mex. Ing. Quim. 2018, 17, 189–202. [Google Scholar] [CrossRef]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—the US Department of Energy’s “Top 10” revisited. Green Chem. 2010, 12, 539–554. [Google Scholar] [CrossRef]
- Lalanne, L.; Nyanhongo, G.S.; Guebitz, G.M.; Pellis, A. Biotechnological production and high potential of furan-based renewable monomers and polymers. Biotechnol. Adv. 2021, 48, 107707. [Google Scholar] [CrossRef]
- De María, P.D.; Guajardo, N. Biocatalytic Valorization of Furans: Opportunities for Inherently Unstable Substrates. ChemSusChem 2017, 10, 4123–4134. [Google Scholar] [CrossRef]
- Hu, L.; He, A.; Liu, X.; Xia, J.; Xu, J.; Zhou, S.; Xu, J. Biocatalytic Transformation of 5-Hydroxymethylfurfural into High-Value Derivatives: Recent Advances and Future Aspects. ACS Sustain. Chem. Eng. 2018, 6, 15915–15935. [Google Scholar] [CrossRef]
- Lin, B.; Tao, Y. Whole-cell biocatalysts by design. Microb. Cell Factories 2017, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Millán, G.G.; Sixta, H. Towards the Green Synthesis of Furfuryl Alcohol in A One-Pot System from Xylose: A Review. Catalysts 2020, 10, 1101. [Google Scholar] [CrossRef]
- Iroegbu, A.O.; Hlangothi, S.P. Furfuryl Alcohol a Versatile, Eco-Sustainable Compound in Perspective. Chem. Afr. 2019, 2, 223–239. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Woortman, A.J.J.; van Ekenstein, G.O.R.A.; Petrović, D.M.; Loos, K. Enzymatic Synthesis of Biobased Polyesters Using 2,5-Bis(hydroxymethyl)furan as the Building Block. Biomacromolecules 2014, 15, 2482–2493. [Google Scholar] [CrossRef]
- Howell, B.A.; Lazar, S.T. Biobased Plasticizers from Carbohydrate-Derived 2,5-Bis(hydroxymethyl)furan. Ind. Eng. Chem. Res. 2018, 58, 1222–1228. [Google Scholar] [CrossRef]
- Saikia, K.; Rathankumar, A.K.; Kumar, P.S.; Varjani, S.; Nizar, M.; Lenin, R.; George, J.; Vaidyanathan, V.K. Recent advances in biotransformation of 5-Hydroxymethylfurfural: Challenges and future aspects. J. Chem. Technol. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Cunha, J.T.; Soares, P.O.; Baptista, S.L.; Costa, C.E.; Domingues, L. Engineered Saccharomyces cerevisiae for lignocellulosic valorization: A review and perspectives on bioethanol production. Bioengineered 2020, 11, 883–903. [Google Scholar] [CrossRef]
- Baptista, S.L.; Costa, C.E.; Cunha, J.T.; Soares, P.O.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae for the production of top value chemicals from biorefinery carbohydrates. Biotechnol. Adv. 2021, 47, 107697. [Google Scholar] [CrossRef]
- Pereira, M.; Schwan, R.; Teixeira, J. Isolation, Screening, and Characterisation of Flocculating and Pectinase Producing Kluyveromyces Strains. Food Technol. Biotechnol. 1999, 37, 257–261. [Google Scholar]
- Costa, C.E.; Romaní, A.; Cunha, J.T.; Johansson, B.; Domingues, L. Integrated approach for selecting efficient Saccharomyces cerevisiae for industrial lignocellulosic fermentations: Importance of yeast chassis linked to process conditions. Bioresour. Technol. 2017, 227, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, A.; Gorwa-Grauslund, M.F.; Hahn-Hägerdal, B.; Lidén, G. Cofactor Dependence in Furan Reduction by Saccharomyces cerevisiae in Fermentation of Acid-Hydrolyzed Lignocellulose. Appl. Environ. Microbiol. 2005, 71, 7866–7871. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef]
- Lulu, L.; Ling, Z.; Dongmei, W.; Xiaolian, G.; Hisanori, T.; Hidehiko, K.; Jiong, H. Identification of a Xylitol Dehydrogenase Gene from Kluyveromyces marxianus NBRC1777. Mol. Biotechnol. 2012, 53, 159–169. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Wang, D.; Gao, X.; Hong, J. Identification of a xylose reductase gene in the xylose metabolic pathway of Kluyveromyces marxianus NBRC1777. J. Ind. Microbiol. Biotechnol. 2011, 38, 2001–2010. [Google Scholar] [CrossRef]
- Martynova, J.; Kokina, A.; Kibilds, J.; Liepins, J.; Scerbaka, R.; Vigants, A. Effects of acetate on Kluyveromyces marxianus DSM 5422 growth and metabolism. Appl. Microbiol. Biotechnol. 2016, 100, 4585–4594. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Mueller, M.; Eichling, S.; Banat, I. Fermentation of xylose by the thermotolerant yeast strains Kluyveromyces marxianus IMB2, IMB4, and IMB5 under anaerobic conditions. Process. Biochem. 2008, 43, 346–350. [Google Scholar] [CrossRef]
- Wang, D.; Wu, D.; Yang, X.; Hong, J. Transcriptomic analysis of thermotolerant yeast Kluyveromyces marxianus in multiple inhibitors tolerance. RSC Adv. 2018, 8, 14177–14192. [Google Scholar] [CrossRef] [Green Version]
- Kundiyana, D.K.; Wilkins, M.; Huhnke, R.; Banat, I. Effect of Furfural Addition on Xylose Utilization by Kluyveromyces marxianus IMB4 under Anaerobic and Microaerobic Conditions. Biol. Eng. 2009, 2, 3–15. [Google Scholar] [CrossRef]
- Samagaci, L.; Ouattara, H.G.; Goualie, B.G.; Niamke, S.L. Growth capacity of yeasts potential starter strains under cocoa fermentation stress conditions in Ivory Coast. Emir. J. Food Agric. 2014, 26, 861. [Google Scholar] [CrossRef]
- Pereira, F.B.; Romaní, A.; Ruiz, H.A.; Teixeira, J.A.; Domingues, L. Industrial robust yeast isolates with great potential for fermentation of lignocellulosic biomass. Bioresour. Technol. 2014, 161, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmqvist, E.; Grage, H.; Meinander, N.Q. Main and interaction effects of acetic acid, furfural, and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol. Bioeng. 1999, 63, 46–55. [Google Scholar] [CrossRef]
- Verho, R.; Londesborough, J.; Penttilä, M.; Richard, P. Engineering Redox Cofactor Regeneration for Improved Pentose Fermentation in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2003, 69, 5892–5897. [Google Scholar] [CrossRef] [Green Version]
- Tarrã o, N.; Becerra, M.; Cerdã¡n, M.E.; González-Siso, M.-I. Reoxidation of cytosolic NADPH in Kluyveromyces lactis. FEMS Yeast Res. 2006, 6, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.T.; Soares, P.O.; Romaní, A.; Thevelein, J.M.; Domingues, L. Xylose fermentation efficiency of industrial Saccharomyces cerevisiae yeast with separate or combined xylose reductase/xylitol dehydrogenase and xylose isomerase pathways. Biotechnol. Biofuels 2019, 12, 20. [Google Scholar] [CrossRef]
- Jeong, H.; Lee, D.-H.; Kim, S.H.; Kim, H.-J.; Lee, K.; Song, J.Y.; Kim, B.K.; Sung, B.H.; Park, J.C.; Sohn, J.H.; et al. Genome Sequence of the Thermotolerant Yeast Kluyveromyces marxianus var. marxianus KCTC 17555. Eukaryot. Cell 2012, 11, 1584–1585. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.-J.; Zhang, M.-L.; Ding, M.; Mai, Z.-M.; Wu, S.-X.; Du, Y.; Feng, J.-X. Alcohol dehydrogenases from Kluyveromyces marxianus: Heterologous expression in Escherichia coliand biochemical characterization. BMC Biotechnol. 2014, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Akita, H.; Watanabe, M.; Suzuki, T.; Nakashima, N.; Hoshino, T. Characterization of the Kluyveromyces marxianus strain DMB1 YGL157w gene product as a broad specificity NADPH-dependent aldehyde reductase. AMB Express 2015, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Horváth, I.S.; Franzén, C.J.; Taherzadeh, M.J.; Niklasson, C.; Lidén, G. Effects of Furfural on the Respiratory Metabolism of Saccharomyces cerevisiae in Glucose-Limited Chemostats. Appl. Environ. Microbiol. 2003, 69, 4076–4086. [Google Scholar] [CrossRef] [Green Version]
- Ask, M.; Bettiga, M.; Duraiswamy, V.R.; Olsson, L. Pulsed addition of HMF and furfural to batch-grown xylose-utilizing Saccharomyces cerevisiae results in different physiological responses in glucose and xylose consumption phase. Biotechnol. Biofuels 2013, 6, 181. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-M.; Zhang, X.-Y.; Li, N.; Xu, P.; Lou, W.-Y.; Zong, M.-H. Biocatalytic Reduction of HMF to 2,5-Bis(hydroxymethyl)furan by HMF-Tolerant Whole Cells. Chem. Sus. Chem 2016, 10, 372–378. [Google Scholar] [CrossRef]
- Xia, Z.-H.; Zong, M.-H.; Li, N. Catalytic synthesis of 2,5-bis(hydroxymethyl)furan from 5-hydroxymethylfurfual by recombinant Saccharomyces cerevisiae. Enzym. Microb. Technol. 2020, 134, 109491. [Google Scholar] [CrossRef]
- Xu, Z.-H.; Cheng, A.-D.; Xing, X.-P.; Zong, M.-H.; Bai, Y.-P.; Li, N. Improved synthesis of 2,5-bis(hydroxymethyl)furan from 5-hydroxymethylfurfural using acclimatized whole cells entrapped in calcium alginate. Bioresour. Technol. 2018, 262, 177–183. [Google Scholar] [CrossRef]
- Chang, S.; He, X.; Li, B.; Pan, X. Improved Bio-Synthesis of 2,5-bis(hydroxymethyl)furan by Burkholderia contaminans NJPI-15 With Co-substrate. Front. Chem. 2021, 9, 1–7. [Google Scholar] [CrossRef]
- He, Y.-C.; Jiang, C.-X.; Chong, G.-G.; Di, J.-H.; Ma, C.-L. Biological synthesis of 2,5-bis(hydroxymethyl)furan from biomass-derived 5-hydroxymethylfurfural by E. coli CCZU-K14 whole cells. Bioresour. Technol. 2018, 247, 1215–1220. [Google Scholar] [CrossRef]
- Chen, D.; Cang, R.; Zhang, Z.-D.; Huang, H.; Ji, X.-J. Efficient reduction of 5-hydroxymethylfurfural to 2, 5-bis (hydroxymethyl) furan by a fungal whole-cell biocatalyst. Mol. Catal. 2021, 500, 111341. [Google Scholar] [CrossRef]
- Millán, A.; Sala, N.; Torres, M.; Canela-Garayoa, R. Biocatalytic Transformation of 5-Hydroxymethylfurfural into 2,5-di(hydroxymethyl)furan by a Newly Isolated Fusarium striatum Strain. Catalysts 2021, 11, 216. [Google Scholar] [CrossRef]
- Mandalika, A.; Qin, L.; Sato, T.K.; Runge, T. Integrated biorefinery model based on production of furans using open-ended high yield processes. Green Chem. 2014, 16, 2480–2489. [Google Scholar] [CrossRef]
- Heer, D.; Sauer, U. Identification of furfural as a key toxin in lignocellulosic hydrolysates and evolution of a tolerant yeast strain. Microb. Biotechnol. 2008, 1, 497–506. [Google Scholar] [CrossRef]
- Taherzadeh, M.; Gustafsson, L.; Niklasson, C.; Lidén, G. Conversion of furfural in aerobic and anaerobic batch fermentation of glucose by Saccharomyces cerevisiae. J. Biosci. Bioeng. 1999, 87, 169–174. [Google Scholar] [CrossRef]
- Choi, S.Y.; Gong, G.; Park, H.-S.; Um, Y.; Sim, S.J.; Woo, H.M. Extreme furfural tolerance of a soil bacterium Enterobacter cloacae GGT036. J. Biotechnol. 2015, 193, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, Y.; Hori, Y.; Kudou, M.; Ishii, J.; Hasunuma, T.; Kondo, A. Detoxification of furfural in Corynebacterium glutamicum under aerobic and anaerobic conditions. Appl. Microbiol. Biotechnol. 2014, 98, 8675–8683. [Google Scholar] [CrossRef]
- Ran, H.; Zhang, J.; Gao, Q.; Lin, Z.; Bao, J. Analysis of biodegradation performance of furfural and 5-hydroxymethylfurfural by Amorphotheca resinae ZN1. Biotechnol. Biofuels 2014, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; He, J. Clostridium species strain BOH3 tolerates and transforms inhibitors from horticulture waste hydrolysates. Appl. Microbiol. Biotechnol. 2017, 101, 6289–6297. [Google Scholar] [CrossRef]
- Bu, C.-Y.; Yan, Y.-X.; Zou, L.-H.; Zheng, Z.-J.; Ouyang, J. One-pot biosynthesis of furfuryl alcohol and lactic acid via a glucose coupled biphasic system using single Bacillus coagulans NL01. Bioresour. Technol. 2020, 313, 123705. [Google Scholar] [CrossRef]
- Ishii, J.; Yoshimura, K.; Hasunuma, T.; Kondo, A. Reduction of furan derivatives by overexpressing NADH-dependent Adh1 improves ethanol fermentation using xylose as sole carbon source with Saccharomyces cerevisiae harboring XR–XDH pathway. Appl. Microbiol. Biotechnol. 2013, 97, 2597–2607. [Google Scholar] [CrossRef]
- Hasunuma, T.; Ismail, K.S.K.; Nambu, Y.; Kondo, A. Co-expression of TAL1 and ADH1 in recombinant xylose-fermenting Saccharomyces cerevisiae improves ethanol production from lignocellulosic hydrolysates in the presence of furfural. J. Biosci. Bioeng. 2014, 117, 165–169. [Google Scholar] [CrossRef]
- Kılmanoğlu, H.; Hoşoğlu, M.İ.; Güneşer, O.; Yüceer, Y.K. Optimization of pretreatment and enzymatic hydrolysis conditions of tomato pomace for production of alcohols and esters by Kluyveromyces marxianus. LWT 2020, 138, 110728. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baptista, M.; Cunha, J.T.; Domingues, L. Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives. J. Fungi 2021, 7, 1047. https://doi.org/10.3390/jof7121047
Baptista M, Cunha JT, Domingues L. Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives. Journal of Fungi. 2021; 7(12):1047. https://doi.org/10.3390/jof7121047
Chicago/Turabian StyleBaptista, Marlene, Joana T. Cunha, and Lucília Domingues. 2021. "Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives" Journal of Fungi 7, no. 12: 1047. https://doi.org/10.3390/jof7121047
APA StyleBaptista, M., Cunha, J. T., & Domingues, L. (2021). Establishment of Kluyveromyces marxianus as a Microbial Cell Factory for Lignocellulosic Processes: Production of High Value Furan Derivatives. Journal of Fungi, 7(12), 1047. https://doi.org/10.3390/jof7121047