
Morphology, Phylogeny, and Pathogenicity of Pestalotioid Species on Camellia oleifera in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Morphological and Cultural Characterization
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analyses
2.5. Pathogenicity Testing
3. Results
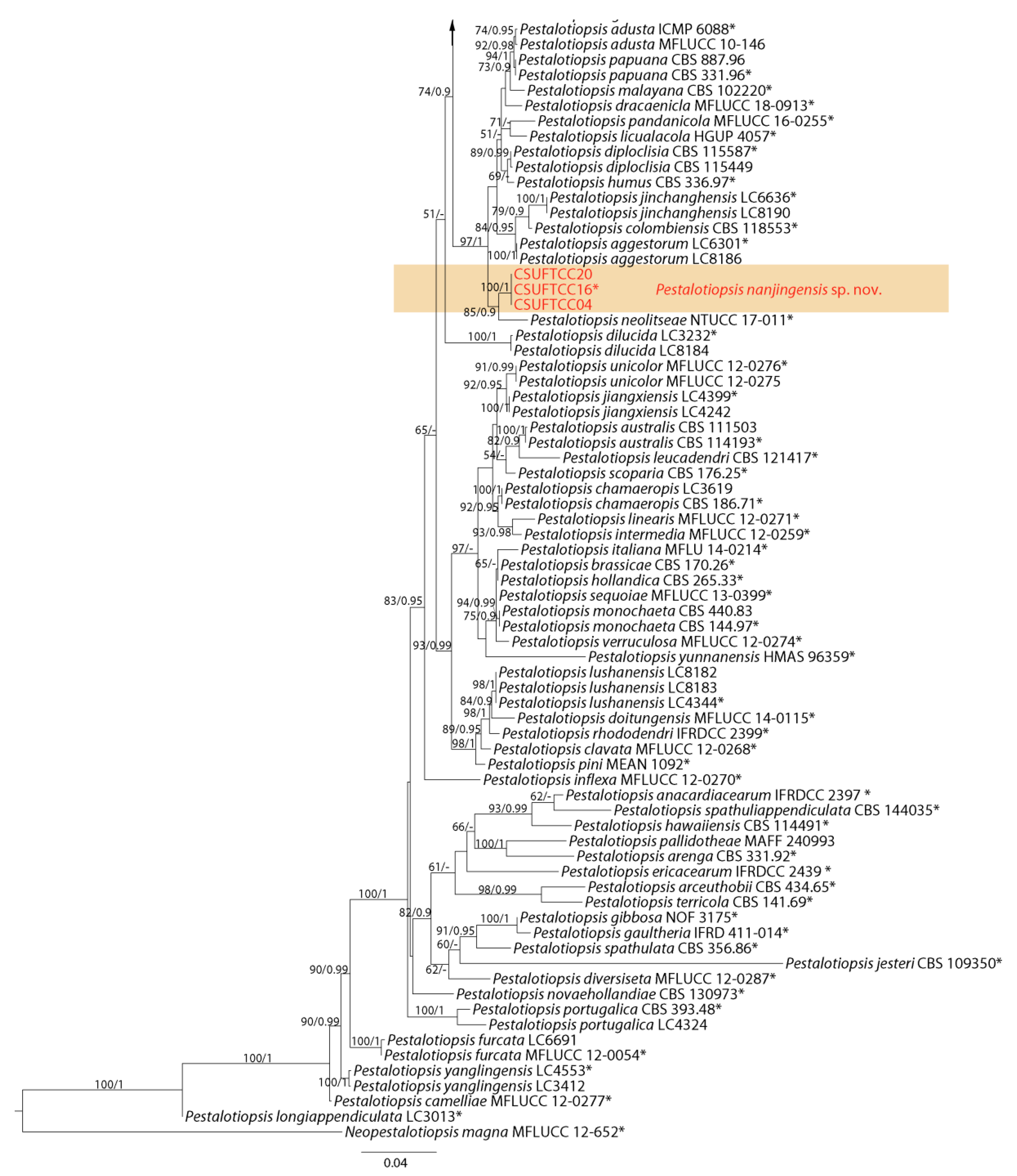
3.1. Phylogenetic Analyses
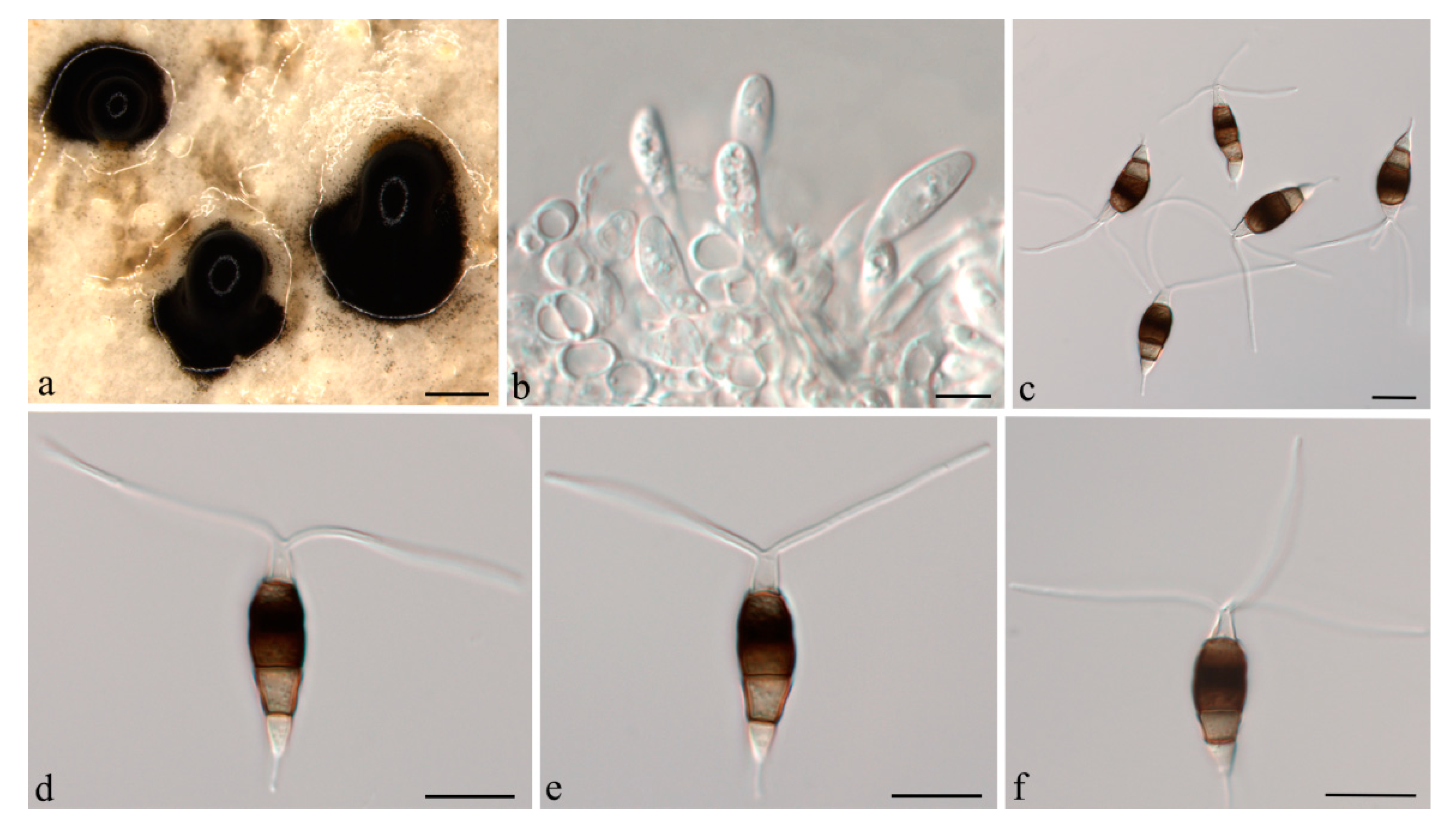
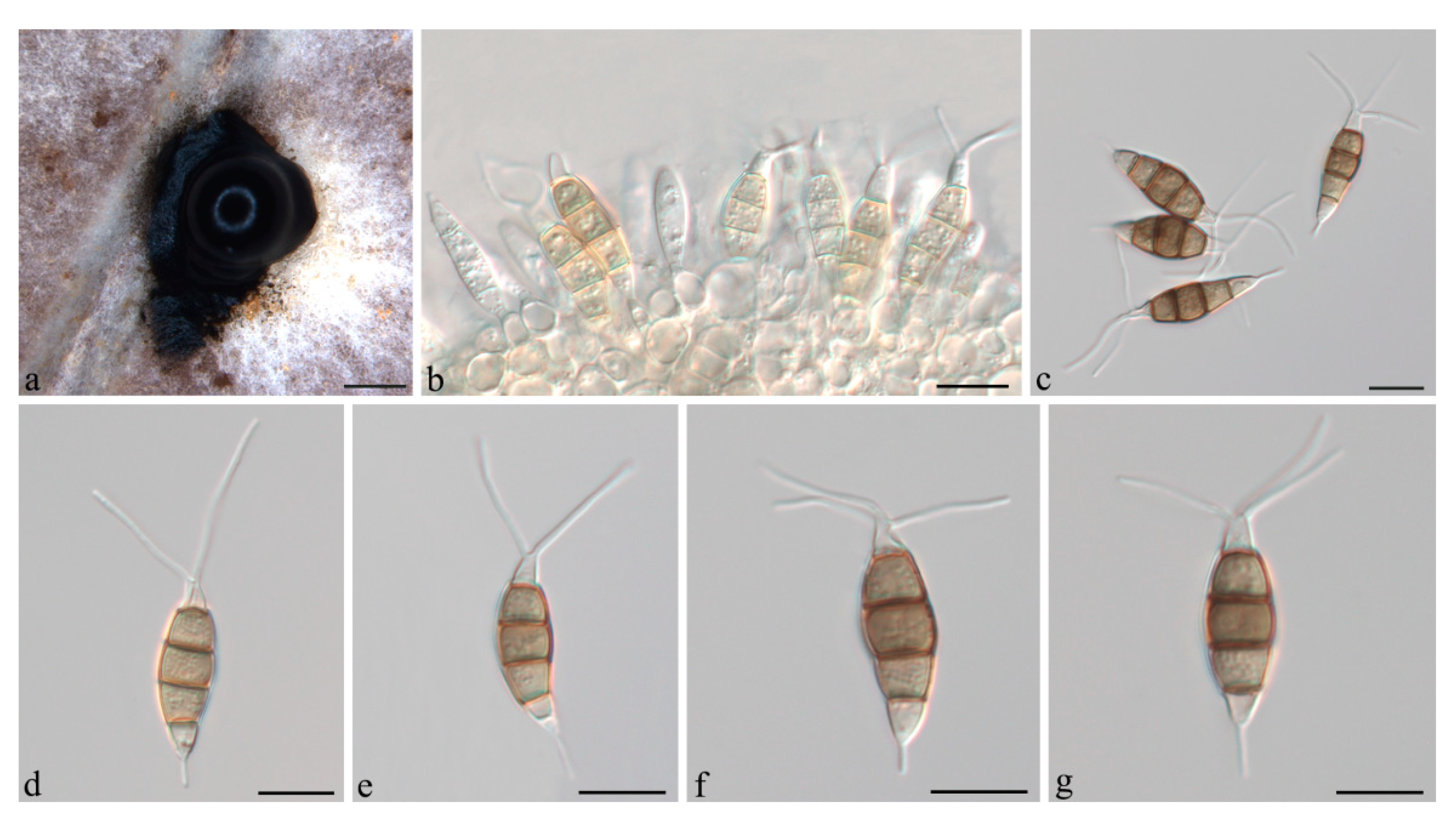
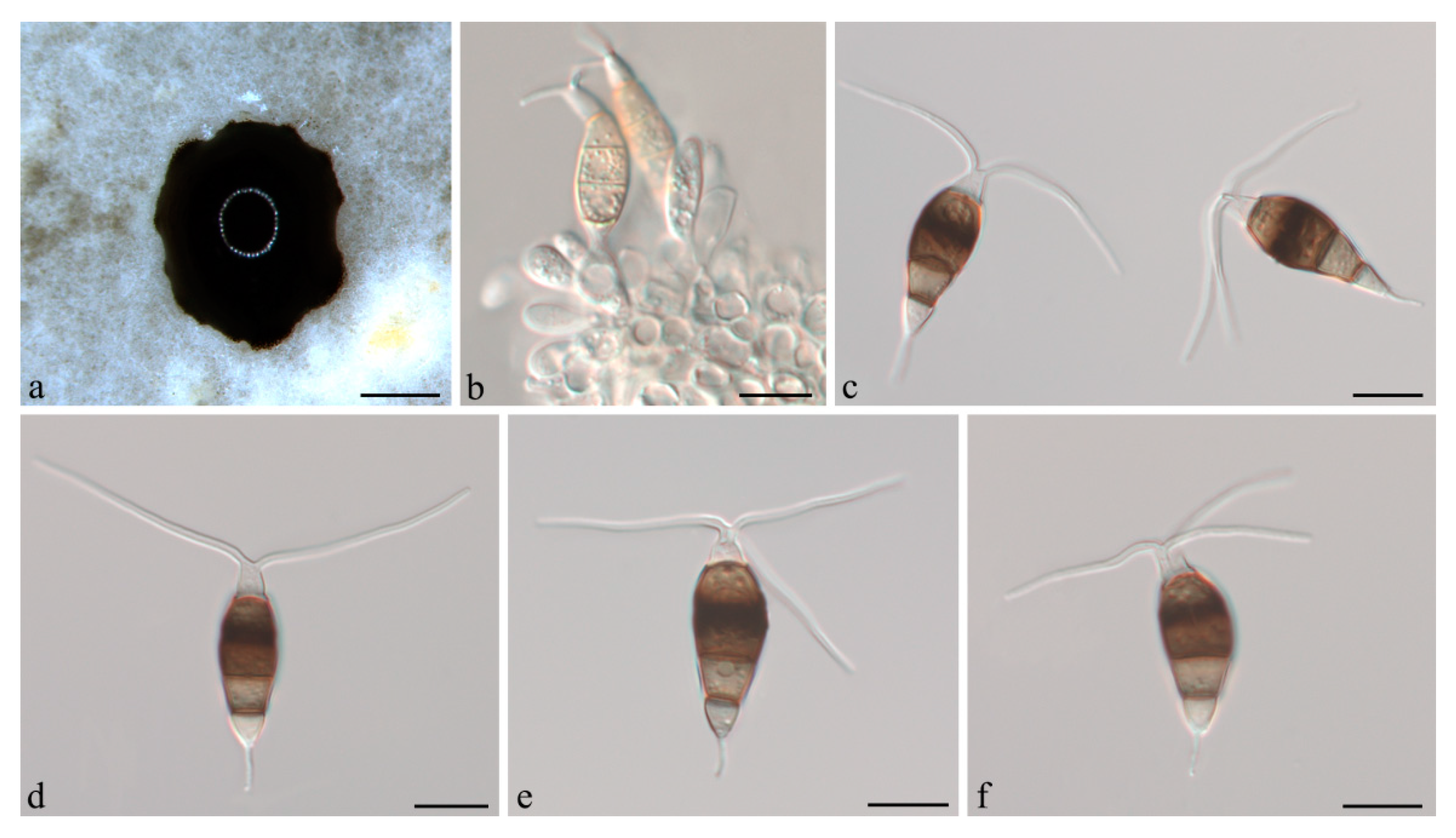
3.2. Taxonomy
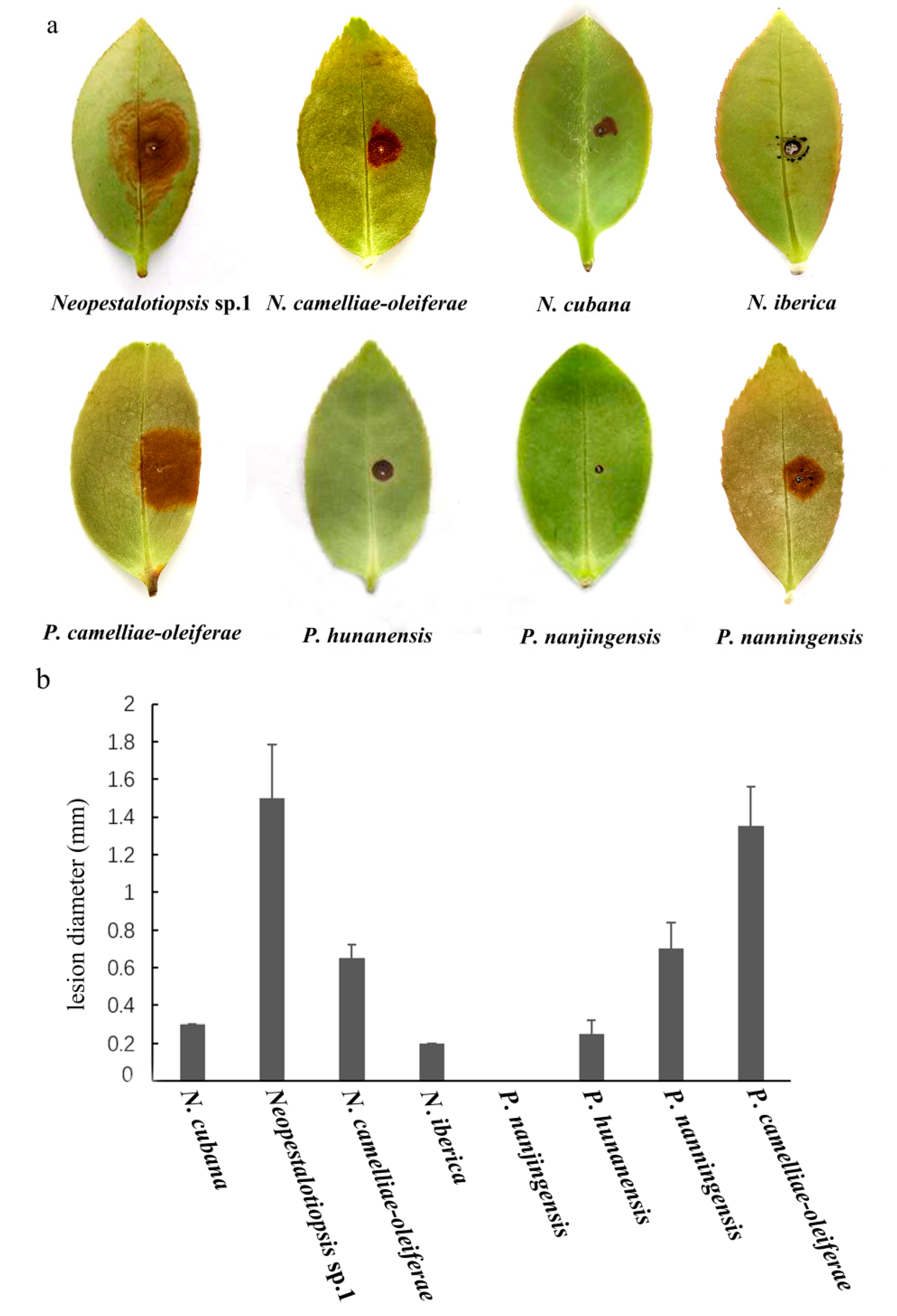
3.3. Pathogenicity Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhuang, R.L. Camellia oleifera, 2nd ed.; China Forestry Press: Beijing, China, 2008. [Google Scholar]
- Wang, W.J.; Chen, C.G.; Cheng, J. The medicinal active role of tea oil in health care. Food Nutr. China 2007, 9, 48–51. [Google Scholar]
- Wang, Y.; Chen, J.Y.; Xu, X.W.; Cheng, J.Y.; Zheng, L.; Huang, J.B.; Li, D.W. Identification and characterization of Colletotrichum species associated with anthracnose disease of Camellia oleifera in China. Plant Dis. 2020, 104, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wu, Y.; He, Z.; Li, M.; Zhu, K.; Gao, B. Diversity and antifungal activity of endophytic fungi associated with Camellia oleifera. Mycobiology 2018, 46, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Hou, C.L. Three new species of Diaporthe from China based on morphological characters and DNA sequence data analyses. Phytotaxa 2019, 422, 157–174. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Hyde, K.D. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef] [Green Version]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; Ekachai, C.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeewon, R.; Liew, E.C.Y.; Hyde, K.D. Phylogenetic relationships of Pestalotiopsis and allied genera inferred from ribosomal DNA sequences and morphological characters. Mol. Phylogenet. Evol. 2002, 25, 378–392. [Google Scholar] [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Simpson, J.A.; Hodgkiss, I.J.; Hyde, K.D. Phylogenetic significance of morphological characters in the taxonomy of Pestalotiopsis species. Mol. Phylogenet. Evol. 2003, 27, 372–383. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Laringnonl, P.; Hyde, K.D.; Al-Sady, A.; Liu, Z. Characterization of Neopestalotiopsis, Pestalotiopsis and Truncatella species associated with grapevine trunk diseases in France. Phytopathol. Mediterr. 2016, 55, 380–390. [Google Scholar]
- Liu, F.; Hou, L.W.; Raza, M.; Cai, L. Pestalotiopsis and allied genera from Camellia, with description of 11 new species from China. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozawa, S.; Yamaguchi, K.; Van Hop, D.; Phay, N.; Ando, K.; Watanabe, K. Identification of two new species and asexual morph from the genus Pseudopestalotiopsis. Mycoscience 2017, 58, 328–337. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Dis. 2017, 101, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Ariyawansa, H.A.; Hyde, K.D. Hyde. Additions to Pestalotiopsis in Taiwan. Mycosphere 2018, 9, 999–1013. [Google Scholar] [CrossRef]
- Jiang, N.; Bonthond, G.; Fan, X.L.; Tian, C.M. Neopestalotiopsis rosicola sp. nov. causing stem canker of Rosa chinensis in China. Mycotaxon 2018, 133, 271–283. [Google Scholar] [CrossRef]
- Tsai, I.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Ariyawansa, H.A. Molecular phylogeny, morphology and pathogenicity of Pseudopestalotiopsis species of Ixora in Taiwan. Mycol. Prog. 2018, 17, 941–952. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Microfungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Jayawardena, R.S.; Chen, Y.; Wen, T.C.; Meepol, W.; Hyde, K.D. Morphological and phylogenetic characterization of novel pestalotioid species associated with mangroves in Thailand. Mycosphere 2019, 10, 531–578. [Google Scholar] [CrossRef]
- Diogo, E.; Gonçalves, C.I.; Silva, A.C.; Valente, C.; Bragança, H.; Phillips, A.J. Five new species of Neopestalotiopsis associated with diseased Eucalyptus spp. in Portugal. Mycol. Prog. 2021, 20, 1441–1456. [Google Scholar] [CrossRef]
- Prasannath, K.; Shivas, R.G.; Galea, V.J.; Akinsanmi, O.A. Neopestalotiopsis species associated with flower diseases of Macadamia integrifolia in Australia. J. Fungi 2021, 7, 771. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Series 1999, 41, 95–98. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and Phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from Fagaceae Leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Cheewangkoon, R.; Gentekaki, E.; Maharachchikumbura, S.S.; Brahmanage, R.S.; Hyde, K.D. Neopestalotiopsis alpapicalis sp. nov. a new endophyte from tropical mangrove trees in Krabi Province (Thailand). Phytotaxa 2019, 393, 251–262. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Machado, A.R.; Firmino, A.L.; Rosado, A.W.C.; Souza, C.A.F.D.; Souza-Motta, C.M.D.; Freire, K.T.L.D.S.; Paiva, L.M.; Magalhães, O.M.C.; Pereira, O.L.; et al. Mycological diversity description I. Acta Bot. Bras. 2018, 32, 656–666. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Tibpromma, S.; Zhang, F.; Xu, J.; Chethana, K.W.T.; Karunarathna, S.C.; Mortimer, P.E. Neopestalotiopsis cavernicola sp. nov. from Gem Cave in Yunnan Province, China. Phytotaxa 2021, 512, 1–27. [Google Scholar] [CrossRef]
- Song, Y.; Geng, K.; Hyde, K.D.; Zhao, W.; Wei, J.G.; Kang, J.C.; Wang, Y. Two new species of Pestalotiopsis from Southern China. Phytotaxa 2013, 126, 22–30. [Google Scholar] [CrossRef]
- Ma, X.Y.; Maharachchikumbura, S.S.; Chen, B.W.; Hyde, K.D.; Mckenzie, E.H.; Chomnunti, P.; Kang, J.C. Endophytic pestalotiod taxa in Dendrobium orchids. Phytotaxa 2019, 419, 268–286. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal Planet description sheets: 371–399. Persoonia 2015, 35, 264. [Google Scholar] [CrossRef]
- Ul Haq, I.; Ijaz, S.; Khan, N.A. Genealogical concordance of phylogenetic species recognition-based delimitation of Neopestalotiopsis species associated with leaf spots and fruit canker disease affected guava plants. Pak. J. Agric. Sci. 2021, 58, 1301–1313. [Google Scholar]
- Freitas, E.F.S.; Da Silva, M.; Barros, M.V.P.; Kasuya, M.C.M. Neopestalotiopsis hadrolaeliae sp. nov., a new endophytic species from the roots of the endangered orchid Hadrolaelia jongheana in Brazil. Phytotaxa 2019, 416, 211–220. [Google Scholar] [CrossRef]
- Huanluek, N.; Jjayawardena, R.S.; Maharachchikumbura, S.S.N.; Harishchandra, D.L. Additions to pestalotioid fungi in Thailand: Neopestalotiopsis hydeana sp. nov. and Pestalotiopsis hydei sp. nov. Phytotaxa 2021, 479, 23–43. [Google Scholar] [CrossRef]
- Ayoubi, N.; Soleimani, M.J. Strawberry fruit rot caused by Neopestalotiopsis iranensis sp. nov., and N. mesopotamica. Curr. Microbiol. 2016, 72, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Maharachchikumbura, S.S.; Jiang, Y.L.; Hyde, K.D.; Wang, Y. Pestalotiopsis keteleeria sp. nov., isolated from Keteleeria pubescens in China. Chiang Mai J. Sci. 2014, 41, 885–893. [Google Scholar]
- Maharachchikumbura, S.S.; Guo, L.D.; Chukeatirote, E.; Hyde, K.D. Improving the backbone tree for the genus Pestalotiopsis; addition of P. steyaertii and P. magna sp. nov. Mycol. Prog. 2014, 13, 617–624. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef]
- Silvério, M.L.; Cavalcanti, M.A.Q.; Silva, G.A.; Oliveira, R.J.V.; Bezerra, J.L. A new epifoliar species of Neopestalotiopsis from Brazil. Agrotropica 2016, 28, 151–158. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Swart, L.; Denman, S.; Taylor, J.E.; Bezuidenhout, C.M.; Palm, M.E.; Marincowitz, S.; Groenewald, J.Z. Fungal pathogens of Proteaceae. Persoonia 2011, 27, 20–45. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zeng, X.Y.; Yuan, J.; Zhang, Q.; He, Y.K.; Wang, Y. Two new species of Neopestalotiopsis from southern China. Biodivers. Data J. 2021, 9, e70446. [Google Scholar] [CrossRef]
- Santos, J.; Hilário, S.; Pinto, G.; Alves, A. Diversity and pathogenicity of pestalotioid fungi associated with blueberry plants in Portugal, with description of three novel species of Neopestalotiopsis. Eur. J. Plant Pathol. 2021, 161, 1–17. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Liu, M.; Maharachchikumbura, S.S.N.; Zang, W.; Xing, Q.K.; Hyde, K.D.; Nilthong, S.; Li, X.; Yan, J. Neopestalotiopsis vitis sp. nov. causing grapevine leaf spot in China. Phytotaxa 2016, 258, 63–74. [Google Scholar] [CrossRef]
- Gu, M.; Hu, D.; Han, B.; Jiang, N.; Tian, C.M. Pestalotiopsis abietis sp. nov. from Abies fargesii in China. Phytotaxa 2021, 509, 93–105. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Silva, A.C.; Diogo, E.; Henriques, J.; Ramos, A.P.; Sandoval-Denis, M.; Crous, P.W.; Bragança, H. Pestalotiopsis pini sp. nov., an Emerging Pathogen on Stone Pine (Pinus pinea L.). Forests 2020, 11, 805. [Google Scholar] [CrossRef]
- Chaiwan, N.; Wanasinghe, D.N.; Mapook, A.; Jayawardena, R.S.; Norphanphoun, C.; Hyde, K.D. Novel species of Pestalotiopsis fungi on Dracaena from Thailand. Mycology 2020, 11, 306–315. [Google Scholar] [CrossRef]
- Zhang, Y.; Maharachchikumbura, S.S.; Tian, Q.; Hyde, K.D. Pestalotiopsis species on ornamental plants in Yunnan Province, China. Sydowia 2013, 65, 113–128. [Google Scholar]
- Maharachchikumbura, S.S.N.; Chukeatirote, E.; Guo, L.-D.; Crous, P.W.; McKenzie, E.H.C.; Hyde, K.D. Pestalotiopsis species associated with Camellia sinensis (tea). Mycotaxon 2013, 123, 47–61. [Google Scholar] [CrossRef]
- Watanabe, K.; Nozawa, S.; Hsiang, T.; Callan, B. The cup fungus Pestalopezia brunneopruinosa is Pestalotiopsis gibbosa and belongs to Sordariomycetes. PLoS ONE 2018, 13, e0197025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Liu, F.; Bonthond, G.; Groenewald, J.Z.; Cai, L.; Crous, P.W. Sporocadaeceae, a family of coelomycetous fungi with appendage-bearing conidia. Stud. Mycol. 2019, 92, 287–415. [Google Scholar] [CrossRef]
- Geng, K.; Zhang, B.; Hyde, K.D.; Kang, J.C.; Wang, Y. A new species of Pestalotiopsis from leaf spots of Licuala grandis from Hainan, China. Phytotaxa 2013, 88, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Motohashi, K.; Ono, Y. Description of Pestalotiopsis pallidotheae: A new species from Japan. Mycoscience 2010, 51, 182–188. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Nanayakkara, R.R.; Zhu, G.S.; Liu, Z.Y. Fungi from Asian Karst formations I. Pestalotiopsis photinicola sp. nov., causing leaf spots of Photinia serrulata. Mycosphere 2017, 8, 103–110. [Google Scholar]
- Tibpromma, S.; Mortimer, P.E.; Karunarathna, S.C.; Zhan, F.; Xu, J.; Promputtha, I.; Yan, K. Morphology and multi-gene phylogeny reveal Pestalotiopsis pinicola sp. nov. and a new host record of Cladosporium anthropophilum from edible pine (Pinus armandii) seeds in Yunnan province, China. Pathogens 2019, 8, 285. [Google Scholar] [CrossRef] [Green Version]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Zhang, Y.; Maharachchikumbura, S.S.; Mckenzie, E.H.; Hyde, K.D. A novel species of Pestalotiopsis causing leaf spots of Trachycarpus fortunei. Cryptogamie Mycol. 2012, 33, 311–318. [Google Scholar] [CrossRef]
- Wei, J.G.; Phan, C.K.; Wang, L.; Xu, T.; Luo, J.T.; Sun, X.; Guo, L.D. Pestalotiopsis yunnanensis sp. nov., an endophyte from Podocarpus macrophyllus (Podocarpaceae) based on morphology and ITS sequence data. Mycol. Prog. 2013, 12, 563–568. [Google Scholar] [CrossRef]
- Wei, J.G.; Xu, T.; Guo, L.D.; Liu, A.R.; Pan, X.H. Endophytic Pestalotiopsis species associated with plants of Podocarpaceae, Theaceae and Taxaceae in southern China. Fungal Divers. 2007, 24, 55–74. [Google Scholar]
- Sessa, L.; Abreo, E.; Lupo, S. Diversity of fungal latent pathogens and true endophytes associated with fruit trees in Uruguay. J. Phytopathol. 2018, 166, 633–647. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate | Host/Substrate | Location | GenBank Accessions Numbers | |||
|---|---|---|---|---|---|---|---|
| ITS | tub2 | tef-1α | References | ||||
| Neopestalotiopsis acrostichi | MFLUCC 17-1754 * | Acrostichum aureum | Thailand | MK764272 | MK764338 | MK764316 | [19] |
| MFLUCC 17-1755 | Acrostichum aureum | Thailand | MK764273 | MK764339 | MK764317 | [19] | |
| N. alpapicalis | MFLUCC 17-2544 * | Rhizophora mucronata | Thailand | MK357772 | MK463545 | MK463547 | [30] |
| MFLUCC 17-2545 | Symptomatic Rhizophora | Thailand | MK357773 | MK463546 | MK463548 | [30] | |
| N. aotearoa | CBS 367.54 * | Canvas | New Zealand | KM199369 | KM199454 | KM199526 | [6] |
| N. asiatica | MFLUCC 12-0286 * | Prunus dulcis | China | JX398983 | JX399018 | JX399049 | [8] |
| N. australis | CBS 114159 * | Telopea sp. | Australia | KM199348 | KM199432 | KM199537 | [8] |
| N. brachiata | MFLUCC 17-1555 * | Rhizophora apiculata | Thailand | MK764274 | MK764340 | MK764318 | [19] |
| N. brasiliensis | COAD 2166 * | Psidium guajava | Brazil | MG686469 | MG692400 | MG692402 | [31] |
| N. camelliae-oleiferae | CSUFTCC81 * | Camellia oleifera | China | OK493585 | OK562360 | OK507955 | This study |
| CSUFTCC82 | Camellia oleifera | China | OK493586 | OK562361 | OK507956 | This study | |
| N. cavernicola | KUMCC 20-0269 * | Cave | China | MW545802 | MW557596 | MW550735 | [32] |
| N. chiangmaiensis | MFLUCC 18-0113 * | Pandanus sp. | Thailand | NA | MH412725 | MH388404 | [18] |
| N. chrysea | MFLUCC 12-0261 * | Dead leaves | China | JX398985 | JX399020 | JX399051 | [6] |
| MFLUCC 12-0262 | Dead leaves | China | JX398986 | JX399021 | JX399052 | [6] | |
| N. clavispora | MFLUCC 12-0281 * | Magnolia sp. | China | JX398979 | JX399014 | JX399045 | [6] |
| MFLUCC 12-0280 | Magnolia sp. | China | JX398978 | JX399013 | JX399044 | [6] | |
| N. cocoës | MFLUCC 15-0152 * | Cocos nucifera | Thailand | NR 156312 | NA | KX789689 | [19] |
| N. coffeae-arabicae | HGUP4015 | Coffea arabica | China | KF412647 | KF412641 | KF412644 | [33] |
| HGUP4019 * | Coffea arabica | China | KF412649 | KF412643 | KF412646 | [33] | |
| N. cubana | CBS 600.96 * | Leaf litter | Cuba | KM199347 | KM199438 | KM199521 | [8] |
| CSUFTCC37 | Camellia oleifera | China | OK493583 | OK562358 | OK507953 | This study | |
| CSUFTCC42 | Camellia oleifera | China | OK493584 | OK562359 | OK507954 | This study | |
| N. dendrobii | MFLUCC 14-0106 * | Dendrobium cariniferum | Thailand | MK993571 | MK975835 | MK975829 | [34] |
| MFLUCC 14-0099 | Dendrobium cariniferum | Thailand | MK993570 | MK975834 | MK975828 | [34] | |
| N. drenthii | BRIP 72263a | Macadamia integrifolia | Australia | MZ303786 | MZ312679 | MZ344171 | [21] |
| BRIP 72264a * | Macadamia integrifolia | Australia | MZ303787 | MZ312680 | MZ344172 | [21] | |
| N. egyptiaca | CBS 1401628 | Mangifera indica | Egypt | KP943747 | KP943746 | KP943748 | [35] |
| N. ellipsospora | MFLUCC 12-02838 | Dead plant material | China | JX398980 | JX399016 | JX399047 | [6] |
| N. eucalyptorum | CBS 147684 * | Eucalyptus globulus | Portugal | MW794108 | MW802841 | MW805397 | [20] |
| N. eucalypticola | CBS 264.37 * | Eucalyptus globulus | NA | KM199376 | KM199431 | KM199551 | [8] |
| N. foedans | CGMCC 3.9123 * | Mangrove plant | China | JX398987 | JX399022 | JX399053 | [6] |
| CGMCC 3.9178 | Neodypsis decaryi | China | JX398989 | JX399024 | JX399055 | [6] | |
| N. formicarum | CBS 362.72 * | Dead ant | Cuba | KM199358 | KM199455 | KM199517 | [8] |
| CBS 115.83 | Plant debris | Cuba | KM199344 | KM199444 | KM199519 | [8] | |
| N. guajavae | FMBCC 11.1 * | Guava | Pakistan | MF783085 | MH460871 | MH460868 | [36] |
| N. guajavicola | FMBCC 11.4 * | Guava | Pakistan | MH209245 | MH460873 | MH460870 | [36] |
| N. hadrolaeliae | EHJ6a | Cattleya jongheana | Brazil | MK454709 | MK465120 | MK465122 | [37] |
| N. hispanica | CBS 147686 * | Eucalyptus globulus | Portugal | MW794107 | MW802840 | MW805399 | [20] |
| N. honoluluana | CBS 114495 * | Telopea sp. | USA | KM199364 | KM199457 | KM199548 | [8] |
| CBS 111535 | Telopea sp. | USA | KM199363 | KM199461 | KM199546 | [8] | |
| N. hydeana | MFLUCC 20-0132 * | Artocarpus heterophyllus | Thailand | MW266069 | MW251119 | MW251129 | [38] |
| N. iberica | CSUFTCC91 | Camellia oleifera | China | OK493587 | OK562362 | OK507957 | This study |
| CSUFTCC92 | Camellia oleifera | China | OK493588 | OK562363 | OK507958 | This study | |
| CSUFTCC93 | Camellia oleifera | China | OK493589 | OK562364 | OK507959 | This study | |
| CBS 147688 * | Eucalyptus globulus | Portugal | MW794111 | MW802844 | MW805402 | [20] | |
| N. iraniensis | CBS 137768 * | Fragaria ananassa | Iran | KM074048 | KM074057 | KM074051 | [39] |
| CBS 137767 | Fragaria ananassa | Iran | KM074045 | KM074056 | KM074053 | [39] | |
| N. javaensis | CBS 257.31 * | Cocos nucifera | Indonesia | KM199357 | KM199457 | KM199548 | [8] |
| N. keteleerie | MFLUCC 13-0915 * | Keteleeria pubescens | China | KJ503820 | KJ503821 | KJ503822 | [40] |
| N. longiappendiculata | CBS 147690 * | Eucalyptus globulus | Portugal | MW794110 | MW802845 | MW805404 | [20] |
| N. lusitanica | CBS 147692 * | Eucalyptus globulus | Portugal | MW794112 | MW802843 | MW805406 | [20] |
| N. macadamiae | BRIP 63737c * | Macadamia integrifolia | Australia | KX186604 | KX186654 | KX186629 | [14] |
| BRIP 63742a | Macadamia integrifolia | Australia | KX186599 | KX186657 | KX186627 | [14] | |
| N. maddoxii | BRIP 72266a * | Macadamia integrifolia | Australia | MZ303782 | MZ312675 | MZ344167 | [14] |
| N. magna | MFLUCC 12-0652 * | Pteridium sp. | France | KF582795 | KF582793 | KF582791 | [41] |
| N. mesopotamica | CBS 336.86 * | Pinus brutia | Iraq | KM199362 | KM199441 | KM199555 | [8] |
| CBS 299.74 | Eucalyptus sp. | Turkey | KM199361 | KM199435 | KM199541 | [8] | |
| N. musae | MFLUCC 15-0776 * | Musa sp. | Thailand | KX789683 | KX789686 | KX789685 | [19] |
| N. natalensis | CBS 138.41 * | Acacia mollissima | South Africa | KM199377 | KM199466 | KM199552 | [8] |
| N. nebuloides | BRIP 66617 * | Sporobolus elongatus | Australia | MK966338 | MK977632 | MK977633 | [42] |
| N. olumideae | BRIP 72273a * | Macadamia integrifolia | Australia | MZ303790 | MZ312683 | MZ344175 | [21] |
| N. pandanicola | KUMCC 17-0175 | Pandanus sp. | China | NA | MH412720 | MH388389 | [18] |
| N. pernambucana | URM7148-01 * | Vismia guianensis | Brazil | KJ792466 | NA | KU306739 | [43] |
| URM7148-02 | Vismia guianensis | Brazil | KJ792467 | NA | KU306740 | [43] | |
| N. perukae | FMBCC 11.3 * | Guava | Pakistan | MH209077 | MH460876 | MH523647 | [36] |
| N. petila | MFLUCC 17-1738 | Rhizophora mucronata | Thailand | MK764275 | MK764341 | MK764319 | [19] |
| MFLUCC 17-1737 * | Rhizophora mucronata | Thailand | MK764276 | MK764342 | MK764320 | [19] | |
| N. phangngaensis | MFLUCC 18-0119 * | Pandanus sp. | Thailand | MH388354 | MH412721 | MH388390 | [18] |
| N. piceana | CBS 254.32 | Cocos nucifera | Indonesia | KM199372 | KM199452 | KM199529 | [8] |
| CBS 394.48 * | Picea sp. | UK | KM199368 | KM199453 | KM199527 | [8] | |
| N. protearum | CBS 114178 * | Leucospermum cuneiforme cv. “Sunbird” | Zimbabwe | JN712498 | KM199463 | LT853201 | [44] |
| N. psidii | FMBCC 11.2 * | Guava | Pakistan | MF783082 | MH477870 | MH460874 | [36] |
| N. rhapidis | GUCC 21501 * | Rhododendron simsii | China | MW931620 | MW980441 | MW980442 | [45] |
| N. rhizophorae | MFLUCC 17-1550 * | Rhizophora mucronata | Thailand | MK764277 | MK764343 | MK764321 | [19] |
| MFLUCC 17-1551 | Rhizophora mucronata | Thailand | MK764278 | MK764344 | MK764322 | [19] | |
| N. rhododendri | GUCC 21504 * | Rhododendron simsii | China | MW979577 | MW980443 | MW980444 | [45] |
| GUCC 21505 | Rhododendron simsii | China | MW979576 | MW980445 | MW980446 | [45] | |
| N. rosae | CBS 101057 * | Rosa sp. | New Zealand | KM199359 | KM199429 | KM199523 | [8] |
| CBS 124745 | Paeonia suffruticosa | USA | KM199360 | KM199430 | KM199524 | [8] | |
| N. rosicola | CFCC 51992 * | Rosa chinensis | China | KY885239 | KY885245 | KY885243 | [15] |
| CFCC 51993 | Rosa chinensis | China | KY885240 | KY885246 | KY885244 | [15] | |
| N. samarangensis | CBS 115451 | Unidentified tree | China | KM199365 | KM199447 | KM199556 | [8] |
| N. saprophytica | MFLUCC 12-0282 * | Magnolia sp. | China | JX398982 | JX399017 | JX399048 | [8] |
| N. scalabiensis | MUM 21.34 * | Vaccinium corymbosum | Portugal | MW969748 | MW934611 | MW959100 | [46] |
| N. sichuanensis | CFCC 54338 * | Castanea mollissima | China | MW166231 | MW218524 | MW199750 | [16] |
| SM15-1C | Castanea mollissima | China | MW166232 | MW218525 | MW199751 | [16] | |
| N. sonneratae | MFLUCC 17-1745 * | Sonneronata alba | Thailand | MK764279 | MK764345 | MK764323 | [19] |
| MFLUCC 17-1744 | Sonneronata alba | Thailand | MK764280 | MK764346 | MK764324 | [19] | |
| Neopestalotiopsis sp.1 | CSUFTCC61 | Camellia oleifera | China | OK493590 | OK562365 | OK507960 | This study |
| CSUFTCC62 | Camellia oleifera | China | OK493591 | OK562366 | OK507961 | This study | |
| CSUFTCC63 | Camellia oleifera | China | OK493592 | OK562367 | OK507962 | This study | |
| N. steyaertii | IMI 192475 * | Eucalyptus viminalis | Australia | KF582796 | KF582794 | KF582792 | [8] |
| N. surinamensis | CBS 450.74 * | Soil under Elaeis guineensis | Suriname | KM199351 | KM199465 | KM199518 | [8] |
| N. thailandica | MFLUCC 17-1730 * | Rhizophora mucronata | Thailand | MK764281 | MK764347 | MK764325 | [19] |
| MFLUCC 17-1731 | Rhizophora mucronata | Thailand | MK764282 | MK764348 | MK764326 | [19] | |
| N. umbrinospora | MFLUCC 12-0285 * | Unidentified plant | China | JX398984 | JX399019 | JX399050 | [6] |
| N. vaccinii | MUM 21.36 * | Vaccinium corymbosum | Portugal | MW969747 | MW934610 | MW959099 | [46] |
| N. vacciniicola | MUM 21.35 * | Vaccinium corymbosum | Portugal | MW969751 | MW934614 | MW959103 | [46] |
| N. vheenae | BRIP 72293a * | Macadamia integrifolia | Australia | MZ303792 | MZ312685 | MZ344177 | [21] |
| N. vitis | MFLUCC 15-1265 * | Vitis vinifera cv. “Summer black” | China | KU140694 | KU140685 | KU140676 | [47] |
| MFLUCC 15-1270 | Vitis vinifera cv. “Kyoho” | China | KU140699 | KU140690 | KU140681 | [47] | |
| N. zakeelii | BRIP 72282a * | Macadamia integrifolia | Australia | MZ303789 | MZ312682 | MZ344174 | [21] |
| N. zimbabwana | CBS 111495 * | Leucospermum cunciforme | Zimbabwe | JX556231 | KM199456 | KM199545 | [8] |
| Pestalotiopsis abietis | CFCC 53011 * | Abies fargesii | China | MK397013 | MK622280 | MK622277 | [48] |
| CFCC 53012 | Abies fargesii | China | MK397014 | MK622281 | MK622278 | [48] | |
| CFCC 53013 | Abies fargesii | China | MK397015 | MK622282 | MK622279 | [48] | |
| P. adusta | ICMP 6088 * | Refrigerator door | Fiji | JX399006 | JX399037 | JX399070 | [6] |
| MFLUCC 10-146 | Syzygium sp. | Thailand | JX399007 | JX399038 | JX399071 | [6] | |
| P. aggestorum | LC6301 * | Camellia sinensis | China | KX895015 | KX895348 | KX895234 | [12] |
| LC8186 | Camellia sinensis | China | KY464140 | KY464160 | KY464150 | [12] | |
| P. anacardiacearum | IFRDCC 2397 * | Mangifera indica | China | KC247154 | KC247155 | KC247156 | [8] |
| P. arceuthobii | CBS 434.65 * | Arceuthobium campylopodum | USA | KM199341 | KM199427 | KM199516 | [8] |
| P. arenga | CBS 331.92 * | Arenga undulatifolia | Singapore | KM199340 | KM199426 | KM199515 | [8] |
| P. australasia | CBS 114126 * | Knightia sp. | New Zealand | KM199297 | KM199409 | KM199499 | [8] |
| CBS 114141 | Protea sp. | New South Wales | KM199298 | KM199410 | KM199501 | [8] | |
| P. australis | CBS 111503 | Protea neriifolia × susannae cv. “Pink Ice” | South Africa | KM199331 | KM199382 | KM199557 | [8] |
| CBS 114193 * | Grevillea sp. | New South Wales | KM199332 | KM199383 | KM199475 | [8] | |
| P. biciliata | CBS 124463 * | Platanus × hispanica | Slovakia | KM199308 | KM199399 | KM199505 | [8] |
| CBS 236.38 | Paeonia sp. | Italy | KM199309 | KM199401 | KM199506 | [8] | |
| P. brachiata | LC2998 * | Camellia sp. | China | KX894933 | KX895265 | KX895150 | [12] |
| LC8188 | Camellia sp. | China | KY464142 | KY464162 | KY464152 | [12] | |
| LC8189 | Camellia sp. | China | KY464143 | KY464163 | KY464153 | [12] | |
| P. brassicae | CBS 170.26 * | Brassica napus | New Zealand | KM199379 | NA | KM199558 | [8] |
| P. camelliae | MFLUCC 12-0277 * | Camellia japonica | China | JX399010 | JX399041 | JX399074 | [6] |
| P. camelliae-oleiferae | CSUFTCC08 * | Camellia oleifera | China | OK493593 | OK562368 | OK507963 | In this study |
| CSUFTCC09 | Camellia oleifera | China | OK493594 | OK562369 | OK507964 | In this study | |
| CSUFTCC10 | Camellia oleifera | China | OK493595 | OK562370 | OK507965 | In this study | |
| P. chamaeropis | CBS 186.71 * | Chamaerops humilis | Italy | KM199326 | KM199391 | KM199473 | [6] |
| LC3619 | Camellia sp. | China | KX894991 | KX895322 | KX895208 | [12] | |
| P. clavata | MFLUCC 12-0268 * | Buxus sp. | China | JX398990 | JX399025 | JX399056 | [6] |
| P. colombiensis | CBS 118553 * | Eucalyptus eurograndis | Colombia | KM199307 | KM199421 | KM199488 | [8] |
| P. digitalis | MFLU 14-0208 * | Digitalis purpurea | New Zealand | KP781879 | KP781883 | NA | [49] |
| P. dilucida | LC3232 * | Camellia sinensis | China | KX894961 | KX895293 | KX895178 | [12] |
| LC8184 | Camellia sinensis | China | KY464138 | KY464158 | KY464148 | [12] | |
| P. diploclisiae | CBS 115449 | Psychotria tutcheri | China | KM199314 | KM199416 | KM199485 | [8] |
| CBS 115587 * | Diploclisia glaucescens | China | KM199320 | KM199419 | KM199486 | [8] | |
| P. disseminata | CBS 118552 | Eucalyptus botryoides | New Zealand | MH553986 | MH554652 | MH554410 | [12] |
| CBS 143904 | Persea americana | New Zealand | MH554152 | MH554825 | MH554587 | [12] | |
| MEAN 1165 | Pinus pinea | Portugal | MT374687 | MT374712 | MT374699 | [50] | |
| MEAN 1166 | Pinus pinea | Portugal | MT374688 | MT374713 | MT374700 | [50] | |
| P. diversiseta | MFLUCC 12-0287 * | Rhododendron sp. | China | JX399009 | JX399040 | JX399073 | [6] |
| P. doitungensis | MFLUCC 14-0115 * | Dendrobium sp. | Thailand | MK993574 | MK975837 | MK975832 | [34] |
| P. dracaenicla | MFLUCC 18-0913 * | Dracaena sp. | Thailand | MN962731 | MN962733 | MN962732 | [51] |
| P. dracontomelonis | MFLU 14-0207 * | Dracontomelon dao | Thailand | NA | NA | KP781880 | [49] |
| P. ericacearum | IFRDCC 2439 * | Rhododendron delavayi | China | KC537807 | KC537821 | KC537814 | [52] |
| P. etonensis | BRIP 66615 * | Sporobolus jacquemontii | Australia | MK966339 | MK977634 | MK977635 | [42] |
| P. formosana | NTUCC 17-009 * | On dead grass | China | MH809381 | MH809385 | MH809389 | [15] |
| P. furcata | MFLUCC 12-0054 * | Camellia sinensis | Thailand | JQ683724 | JQ683708 | JQ683740 | [53] |
| LC6691 | Camellia sinensis | China | KX895030 | KX895363 | KX895248 | [12] | |
| P. gaultheria | IFRD 411-014 * | Gaultheria forrestii | China | KC537805 | KC537819 | KC537812 | [8] |
| P. gibbosa | NOF 3175 * | Gaultheria shallon | Canada | LC311589 | LC311590 | LC311591 | [54] |
| P. grevilleae | CBS 114127 * | Grevillea sp. | Australia | KM199300 | KM199407 | KM199504 | [8] |
| P. hawaiiensis | CBS 114491 * | Leucospermum sp. | Hawaii | KM199339 | KM199428 | KM199514 | [8] |
| P. hollandica | CBS 265.33 * | Sciadopitys verticillata | Netherlands | KM199328 | KM199388 | KM199481 | [8] |
| P. hispanica | CBS 115391 * | Protea cv. ‘Susara’ | Spain | MH553981 | MH554640 | MH554399 | [8] |
| P. humus | CBS 336.97 * | Soil | Papua New Guinea | KM199317 | KM199420 | KM199484 | [8] |
| P. hunanensis | CSUFTCC15 * | Camellia oleifera | China | OK493599 | OK562374 | OK507969 | In this study |
| CSUFTCC18 | Camellia oleifera | China | OK493600 | OK562375 | OK507970 | In this study | |
| CSUFTCC19 | Camellia oleifera | China | OK493601 | OK562376 | OK507971 | In this study | |
| P. inflexa | MFLUCC 12-0270 * | Unidentified tree | China | JX399008 | JX399039 | JX399072 | [6] |
| P. intermedia | MFLUCC 12-0259 * | Unidentified tree | China | JX398993 | JX399028 | JX399059 | [6] |
| P. italiana | MFLU 14-0214 * | Cupressus glabra | Italy | KP781878 | KP781882 | KP781881 | [49] |
| P. jesteri | CBS 109350 * | Fragraea bodenii | Papua New Guinea | KM199380 | KM199468 | KM199554 | [8] |
| P. jiangxiensis | LC4242 | Eurya sp. | China | KX895035 | KX895327 | KX895213 | [12] |
| LC4399 * | Camellia sp. | China | KX895009 | KX895341 | KX895227 | [12] | |
| P. jinchanghensis | LC6636 * | Camellia sinensis | China | KX895028 | KX895361 | KX895247 | [12] |
| LC8190 | Camellia sinensis | China | KY464144 | KY464164 | KY464154 | [12] | |
| P. kandelicola | NCYU 19-0355 * | Kandelia candel | China | MT560723 | MT563100 | MT563102 | [55] |
| P. kenyana | CBS 442.67 * | Coffea sp. | Kenya | KM199302 | KM199395 | KM199502 | [8] |
| LC6633 | Camellia sinensis | China | KX895027 | KX895360 | KX895246 | [8] | |
| P. knightiae | CBS 111963 | Knightia sp. | New Zealand | KM199311 | KM199406 | KM199495 | [8] |
| CBS 114138 * | Knightia sp. | New Zealand | KM199310 | KM199408 | KM199497 | [8] | |
| P. leucadendri | CBS 121417 * | Leucadendron sp. | South Africa | MH553987 | MH554654 | MH554412 | [56] |
| P. licualacola | HGUP 4057 * | Licuala grandis | China | KC492509 | KC481683 | KC481684 | [57] |
| P. linearis | MFLUCC 12-0271 * | Trachelospermum sp. | China | JX398992 | JX399027 | JX399058 | [6] |
| P. longiappendiculata | LC3013 * | Camellia sinensis | China | KX894939 | KX895271 | KX895156 | [12] |
| P. lushanensis | LC4344 * | Camellia sp. | China | KX895005 | KX895337 | KX895223 | [12] |
| LC8182 | Camellia sp. | China | KY464136 | KY464156 | KY464146 | [12] | |
| LC8183 | Camellia sp. | China | KY464137 | KY464157 | KY464147 | [12] | |
| P. macadamiae | BRIP 63738b * | Macadamia integrifolia | Australia | KX186588 | KX186680 | KX186621 | [14] |
| BRIP 63739a | Macadamia integrifolia | Australia | KX186589 | KX186681 | KX186622 | [14] | |
| BRIP 63739b | Macadamia integrifolia | Australia | KX186587 | KX186679 | KX186620 | [14] | |
| P. malayana | CBS 102220 * | Macaranga triloba | Malaysia | KM199306 | KM199411 | KM199482 | [8] |
| P. monochaeta | CBS 144.97 * | Quercus robur | Netherlands | KM199327 | KM199386 | KM199479 | [8] |
| CBS 440.83 | Taxus baccata | Netherlands | KM199329 | KM199387 | KM199480 | [8] | |
| P. nanjingensis | CSUFTCC16 * | Camellia oleifera | China | OK493602 | OK562377 | OK507972 | This study |
| CSUFTCC20 | Camellia oleifera | China | OK493603 | OK562378 | OK507973 | This study | |
| CSUFTCC04 | Camellia oleifera | China | OK493604 | OK562379 | OK507974 | This study | |
| P. nanningensis | CSUFTCC10 * | Camellia oleifera | China | OK493596 | OK562371 | OK507966 | This study |
| CSUFTCC11 | Camellia oleifera | China | OK493597 | OK562372 | OK507967 | This study | |
| CSUFTCC12 | Camellia oleifera | China | OK493598 | OK562373 | OK507968 | This study | |
| P. neolitseae | NTUCC 17-011 * | On leaf of Neolitsea villosa | Taiwan | MH809383 | MH809387 | MH809391 | [15] |
| P. novaehollandiae | CBS 130973 * | Banksia grandis | Australia | KM199337 | KM199425 | KM199511 | [8] |
| P. oryzae | CBS 111522 | Telopea sp. | USA | KM199294 | KM199394 | KM199493 | [8] |
| CBS 171.26 | NA | Italy | KM199304 | KM199397 | KM199494 | [8] | |
| CBS 353.69 * | Oryza sativa | Denmark | KM199299 | KM199398 | KM199496 | [8] | |
| P. pandanicola | MFLUCC 16-0255 * | Pandanus sp. | Thailand | MH388361 | MH412723 | MH388396 | [18] |
| P. papuana | CBS 331.96 * | Coastal soil | Papua New Guinea | KM199321 | KM199413 | KM199491 | [8] |
| CBS 887.96 | Cocos nucifera | Papua New Guinea | KM199318 | KM199415 | KM199492 | [8] | |
| P. pallidotheae | MAFF 240993 * | Pieris japonica | Japan | NR111022 | LC311584 | LC311585 | [58] |
| P. parva | CBS 265.37 * | Delonix regia | NA | KM199312 | KM199404 | KM199508 | [8] |
| CBS 278.35 | Leucothoe fontanesiana | NA | KM199313 | KM199405 | KM199509 | [8] | |
| P. photinicola | GZCC 16-0028 * | Photinia serrulata | China | KY092404 | KY047663 | KY047662 | [59] |
| P. portugalica | CBS 393.48 * | NA | Portugal | KM199335 | KM199422 | KM199510 | [8] |
| LC4324 | Camellia chekiangoleosa | China | KX895001 | KX895333 | KX895219 | [12] | |
| P. pini | MEAN 1092 * | Pinus pinea | Portugal | MT374680 | MT374705 | MT374693 | [50] |
| P. pinicola | KUMCC 19-0183 * | Pinus armandii | China | MN412636 | MN417507 | MN417509 | [60] |
| P. rhododendri | IFRDCC 2399 * | Rhododendron sinogrande | China | KC537804 | KC537818 | KC537811 | [52] |
| P. rhodomyrtus | HGUP4230 * | Rhodomyrtus tomentosa | China | KF412648 | KF412642 | KF412645 | [33] |
| LC4458 | Camellia sinensis | China | KX895010 | KX895342 | KX895228 | [12] | |
| P. rhizophorae | MFLUCC 17-0416 * | Rhizophora apiculata | Thailand | MK764283 | MK764349 | MK764327 | [19] |
| P. rosea | MFLUCC 12-0258 * | Pinus sp. | China | JX399005 | JX399036 | JX399069 | [6] |
| P. scoparia | CBS 176.25 * | Chamaecyparis sp. | NA | KM199330 | KM199393 | KM199478 | [8] |
| P. sequoiae | MFLUCC 13-0399 * | Sequoia sempervirens | Italy | KX572339 | NA | NA | [61] |
| P. spathulata | CBS 356.86 * | Gevuina avellana | Chile | KM199338 | KM199423 | KM199513 | [8] |
| P. spathuliappendiculata | CBS 144035 * | Phoenix canariensis | Australia | MH554172 | MH554845 | MH554607 | [56] |
| P. telopeae | CBS 114137 | Protea sp. | Australia | KM199301 | KM199469 | KM199559 | [8] |
| CBS 114161 * | Telopea sp. | Australia | KM199296 | KM199403 | KM199500 | [8] | |
| CBS 113606 | Telopea sp. | Australia | KM199295 | KM199402 | KM199498 | [8] | |
| P. terricola | CBS 141.69 * | Soil | Pacific Islands | MH554004 | MH554680 | MH554438 | [56] |
| P. thailandica | MFLUCC 17-1616 * | Rhizophora apiculata | Thailand | MK764285 | MK764351 | MK764329 | [19] |
| P. trachicarpicola | IFRDCC 2403 | Podocarpus macrophyllus | China | KC537809 | KC537823 | KC537816 | [52] |
| LC4523 | Camellia sinensis | China | KX895011 | KX895344 | KX895230 | [12] | |
| MFLUCC 12-0264 | Chrysophyllum sp. | China | JX399004 | JX399035 | JX399068 | [6] | |
| OP068 * | Trachycarpus fortunei | China | JQ845947 | JQ845945 | JQ845946 | [62] | |
| P. unicolor | MFLUCC 12-0276 * | Rhododendron sp. | China | JX398999 | JX399030 | NA | [6] |
| MFLUCC 12-0275 | unidentified tree | China | JX398998 | JX399029 | JX399063 | [6] | |
| P. verruculosa | MFLUCC 12-0274 * | Rhododendron sp. | China | JX398996 | NA | JX399061 | [6] |
| P. yanglingensis | LC4553 * | Camellia sinensis | China | KX895012 | KX895345 | KX895231 | [12] |
| LC3412 | Camellia sinensis | China | KX894980 | KX895312 | KX895197 | [12] | |
| P. yunnanensis | HMAS 96359 * | Podocarpus macrophyllus | China | AY373375 | NA | NA | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yang, Q.; Li, H. Morphology, Phylogeny, and Pathogenicity of Pestalotioid Species on Camellia oleifera in China. J. Fungi 2021, 7, 1080. https://doi.org/10.3390/jof7121080
Li L, Yang Q, Li H. Morphology, Phylogeny, and Pathogenicity of Pestalotioid Species on Camellia oleifera in China. Journal of Fungi. 2021; 7(12):1080. https://doi.org/10.3390/jof7121080
Chicago/Turabian StyleLi, Lingling, Qin Yang, and He Li. 2021. "Morphology, Phylogeny, and Pathogenicity of Pestalotioid Species on Camellia oleifera in China" Journal of Fungi 7, no. 12: 1080. https://doi.org/10.3390/jof7121080
APA StyleLi, L., Yang, Q., & Li, H. (2021). Morphology, Phylogeny, and Pathogenicity of Pestalotioid Species on Camellia oleifera in China. Journal of Fungi, 7(12), 1080. https://doi.org/10.3390/jof7121080