Monocyte-Derived Dendritic Cells Can Revert In Vitro Antigen-Specific Cellular Anergy in Active Human Paracoccidioidomycosis

 , and
, and 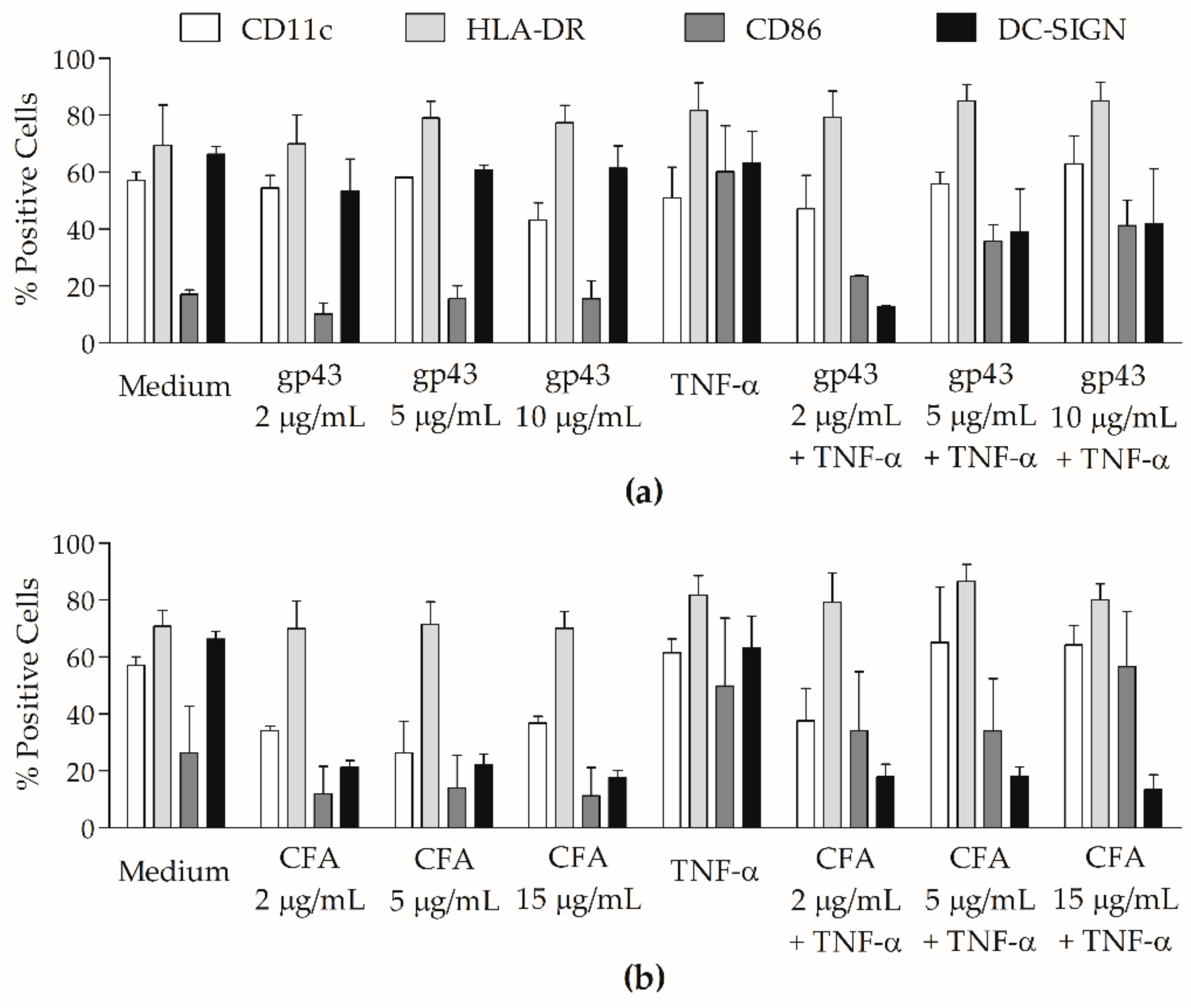{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Paracoccidioidomycosis (PCM) Patients and Control Subjects
2.2. P. brasiliensis Antigens
2.3. Monocyte-Derived Dendritic Cells (MoDCs) Generation and Stimulation
2.4. Flow Cytometric Analyses
2.5. Antigen Presentation Assays
2.6. Cytokine Analyses
2.7. Statistical Analyses
3. Results
3.1. P. brasiliensis Antigens Modulate Surface Molecule Expression on MoDCs from PCM Patients
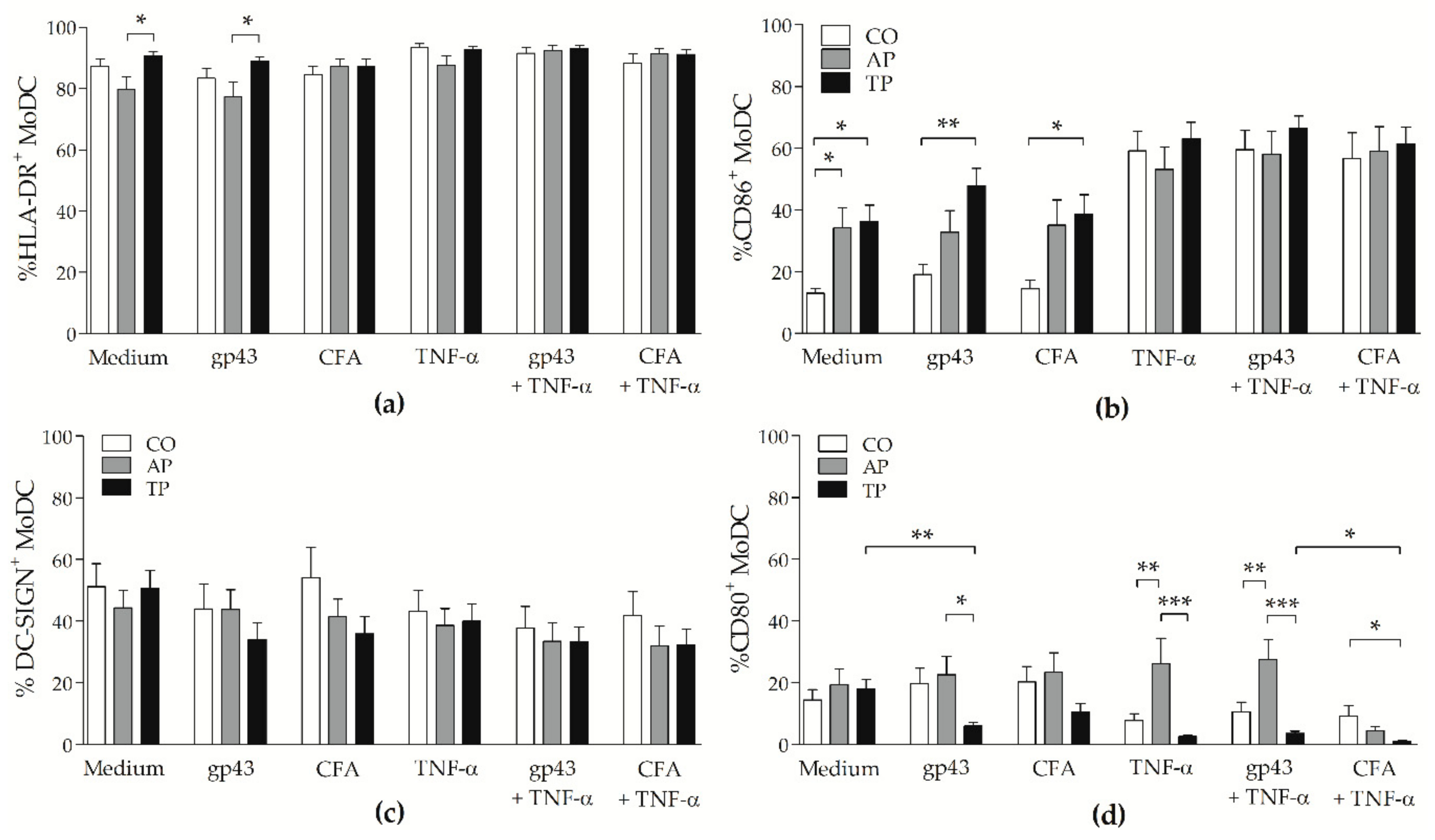
3.2. Patients with Active PCM Have More CD80+ Cells and CD86+ Cells Are Augmented after Antifungal Treatment
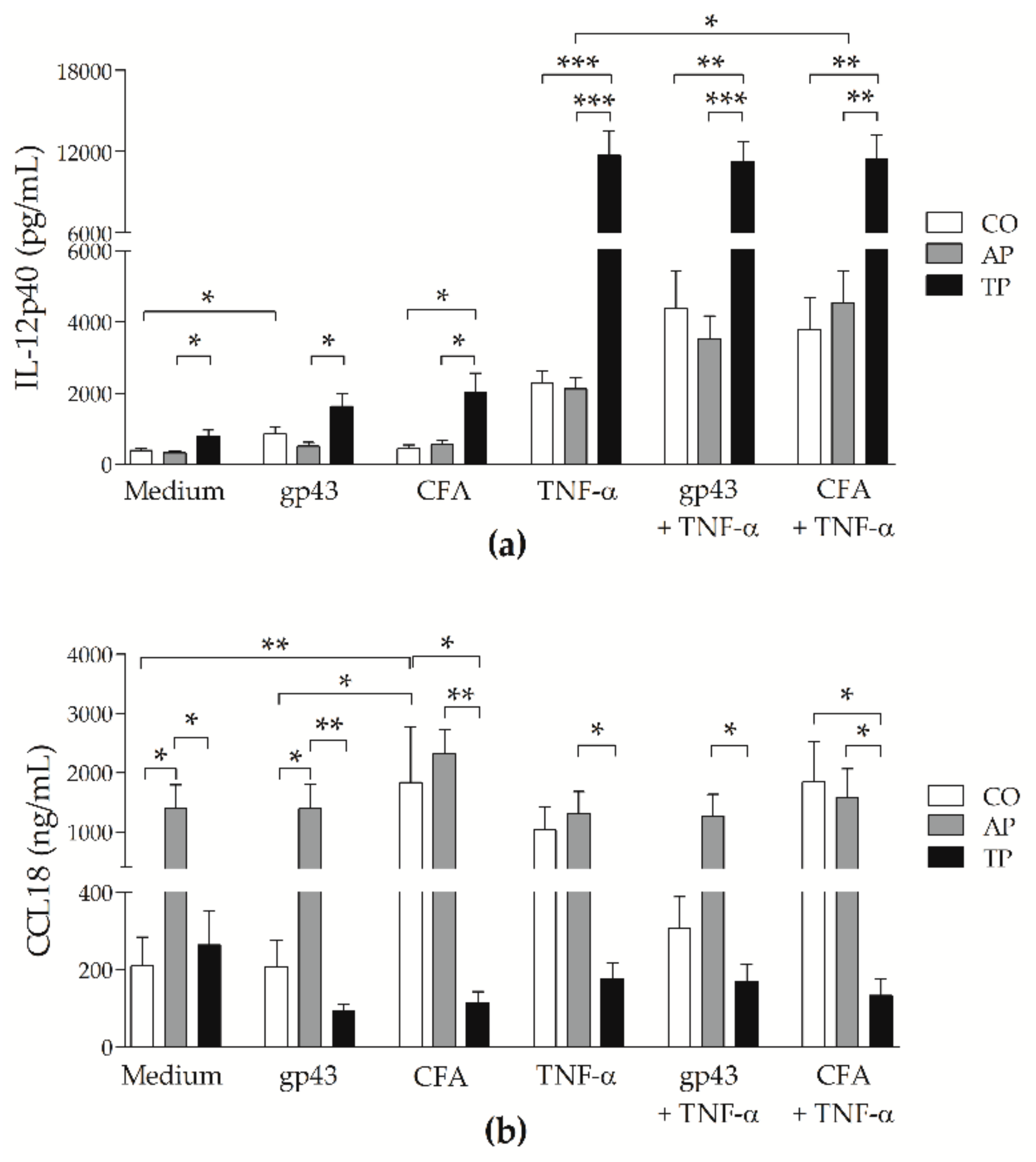
3.3. Patients’ MoDCs Secrete More CCL18 during Active PCM and More IL-12p40 after Antifungal Treatment
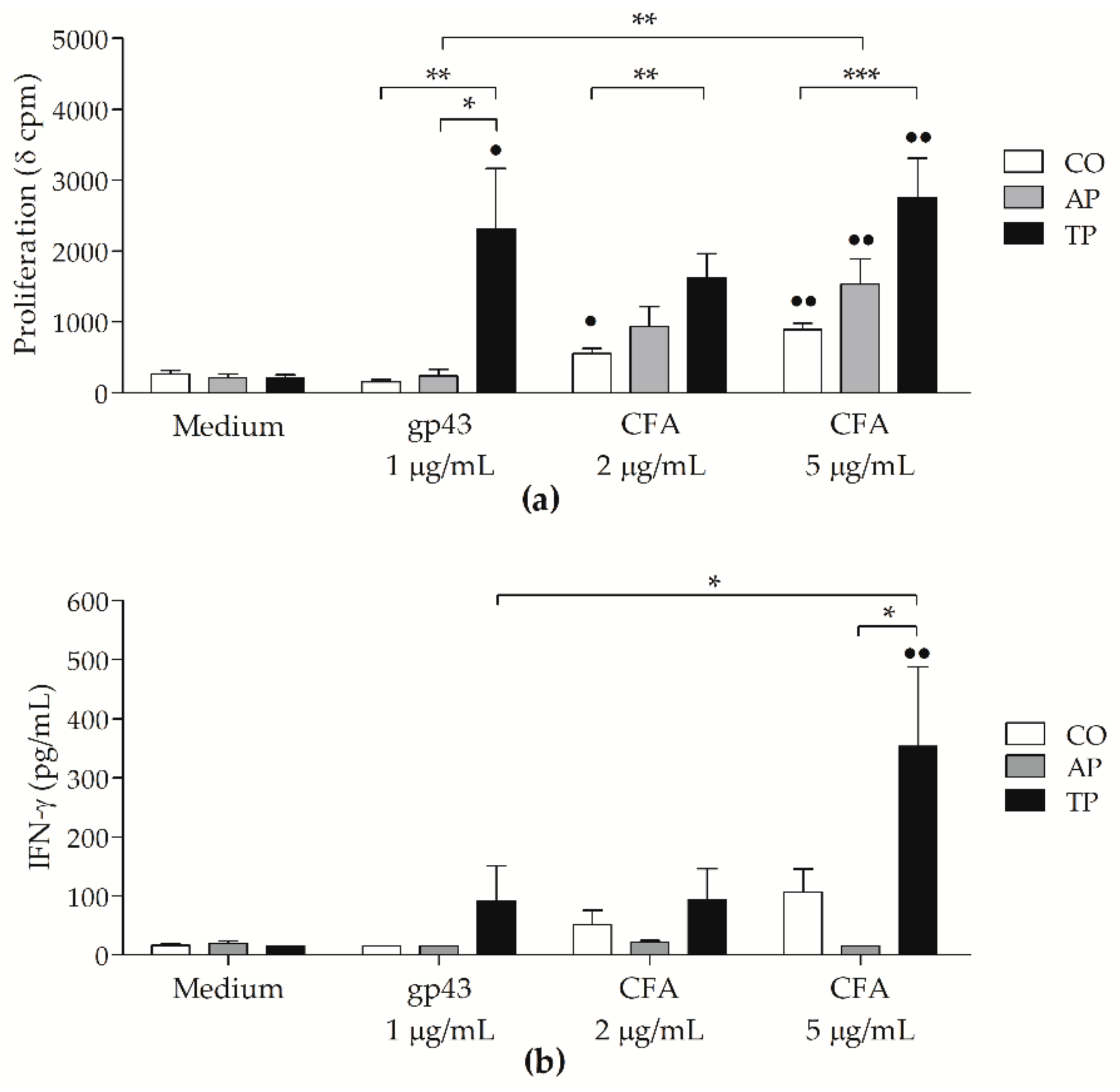
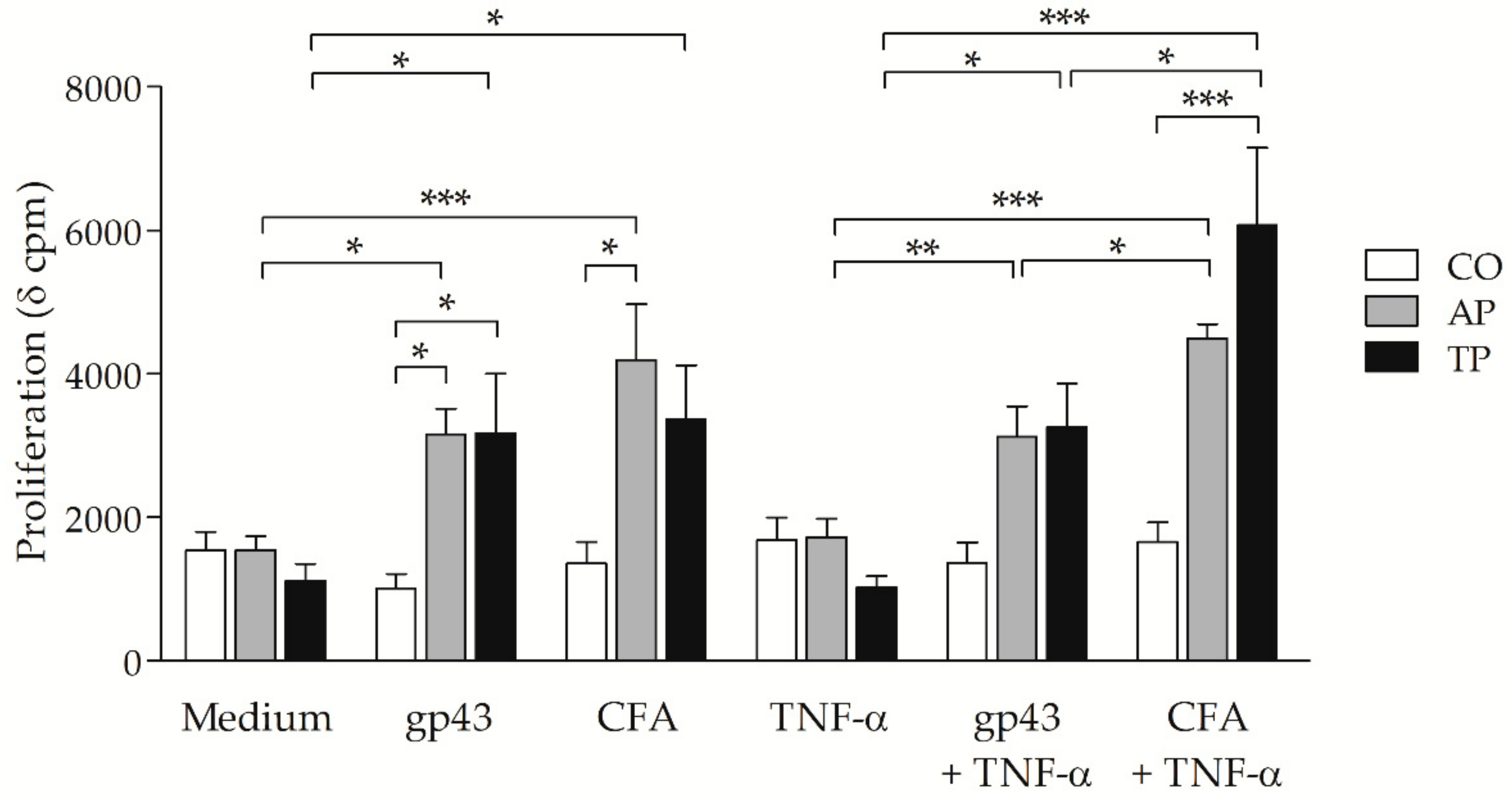
3.4. Cell-Free Antigen (CFA) Induces Proliferation on Peripheral Blood Mononuclear Cells (PBMCs) from Patients with Active PCM But Not Stronger than That from Treated Patients
3.5. P. brasiliensis Antigens-Stimulated MoDCs from PCM Patients Can Induce Strong Proliferative Response
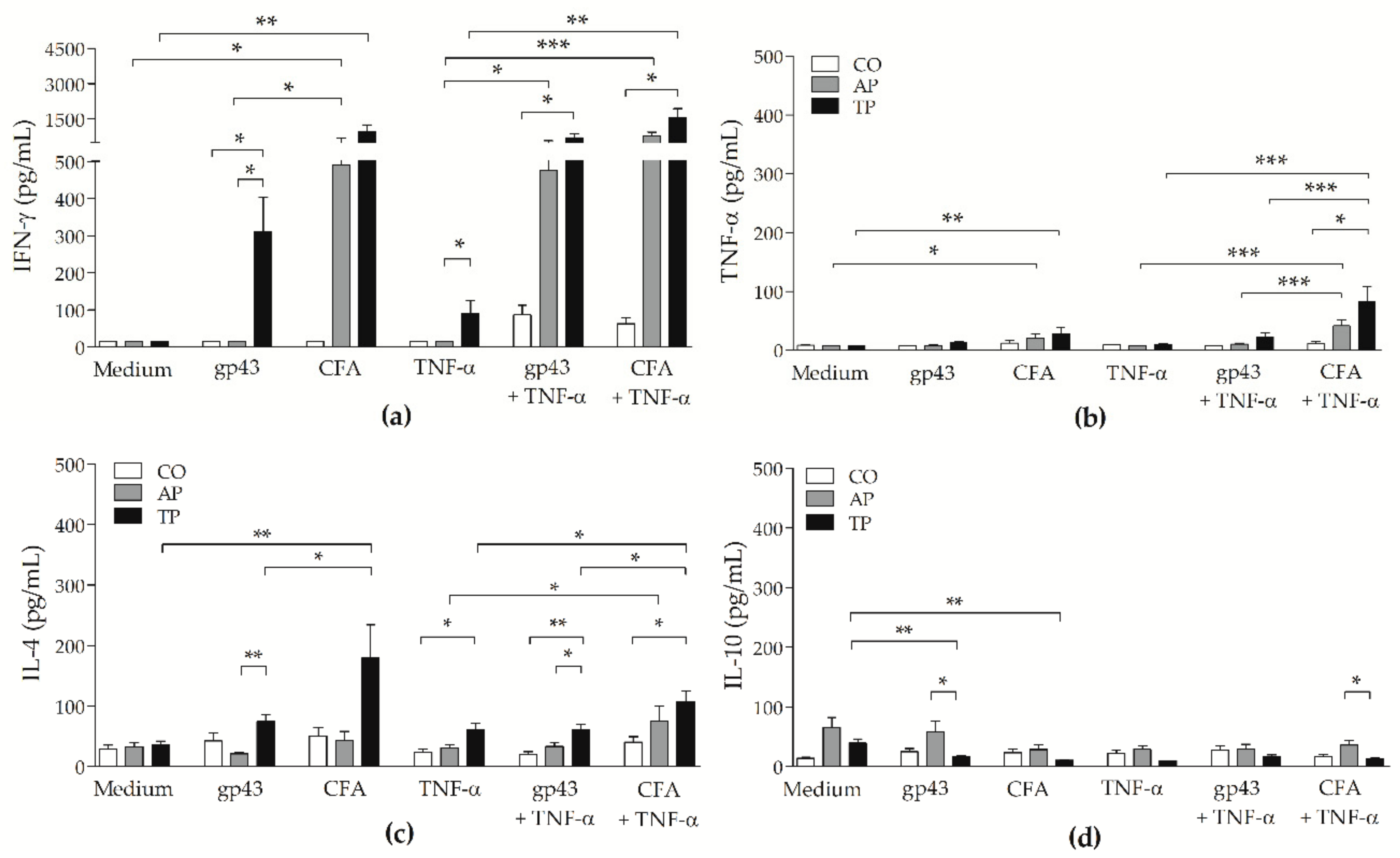
3.6. CFA-Stimulated MoDCs from PCM Patients Up-Regulated IFN-γ and TNF-α Secretion by Lymphocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coutinho, Z.F.; Wanke, B.; Travassos, C.; Oliveira, R.M.; Xavier, D.R.; Coimbra, C.E.A., Jr. Hospital morbidity due to paracoccidioidomycosis in Brazil (1998–2006). Trop. Med. Int. Health 2015, 20, 673–680. [Google Scholar] [CrossRef]
- De Melo Teixeira, M.; Theodoro, R.C.; De Oliveira, F.F.M.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Theodoro, R.C.; Teixeira, M.D.M.; Felipe, M.S.S.; Paduan, K.D.S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus Paracoccidioides: Species Recognition and Biogeographic Aspects. PLoS ONE 2012, 7, e37694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.F.; Lacaz, L.C.; Restrepo-Moreno, A.; Del Negro, G. Paracoccidioidomycosis; CRC Press: Boca Raton, FL, USA, 1994; ISBN 0-8493-4868-4. [Google Scholar]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Shikanai-Yasuda, M.A.; Felipe, M.S.S. Paracoccidioidomycosis: Eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Kashino, S.S.; Fazioli, R.A.; Cafalli-Favati, C.; Meloni-Bruneri, L.H.; Vaz, C.A.C.; Burger, E.; Singer, L.M.; Calich, V.L.G. Resistance to Paracoccidioides brasiliensis Infection Is Linked to a Preferential Th1 Immune Response, Whereas Susceptibility is Associated with Absence of IFN-gamma Production. J. Interf. Cytokine Res. 2000, 20, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Calich, V.L.G.; Mamoni, R.L.; Loures, F.V. Regulatory T cells in paracoccidioidomycosis. Virulence 2019, 10, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.J.; Mamoni, R.L.; Musatti, C.C.; Papaiordanou, P.M.; Blotta, M.H.S. Cytokines and lymphocyte proliferation in juvenile and adult forms of paracoccidioidomycosis: Comparison with infected and non-infected controls. Microbes Infect. 2002, 4, 139–144. [Google Scholar] [CrossRef]
- De Castro, L.F.; Ferreira, M.C.; Da Silva, R.M.; de Souza Lima Blotta, M.H.; Longhi, L.N.A.; Mamoni, R.L. Characterization of the immune response in human paracoccidioidomycosis. J. Infect. 2013, 67, 470–485. [Google Scholar] [CrossRef]
- Benard, G.; Mendes-Gianinni, M.J.; Juvenale, M.; Miranda, E.T.; Duarte, A.J.S. Immunosuppression in paracoccidioidomycosis: T cell hyporesponsiveness to two Paracoccidioides brasiliensis glycoproteins that elicit strong humoral immune response. J. Infect. Dis. 1997, 175, 1263–1267. [Google Scholar] [CrossRef] [Green Version]
- Karhawi, A.S.K.; Colombo, A.L.; Salomão, R. Production of IFN-γ is impaired in patients with paracoccidioidomycosis during active disease and is restored after clinical remission. Med. Mycol. 2000, 38, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Shortman, K.; Liu, Y.-J. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002, 2, 151–161. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, S.R.; Lopes, J. The low efficiency of dendritic cells and macrophages from mice susceptible to Paracoccidioides brasiliensis in inducing a Th1 response. Braz. J. Med. Biol. Res. 2001, 34, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calich, V.L.; Singer-Vermes, L.M.; Siqueira, A.M.; Burger, E. Susceptibility and resistance of inbred mice to Paracoccidioides brasiliensis. Br. J. Exp. Pathol. 1985, 66, 585–594. [Google Scholar] [PubMed]
- Ferreira, K.S.; Lopes, J.D.; De Almeida, S.R. Down-regulation of dendritic cell activation induced by Paracoccidioides brasiliensis. Immunol. Lett. 2004, 94, 107–114. [Google Scholar] [CrossRef]
- Ferreira, K.; De Almeida, S.R. Immunization of susceptible mice with gp43-pulsed dendritic cells induce an increase of pulmonary Paracoccidioidomycosis. Immunol. Lett. 2006, 103, 121–126. [Google Scholar] [CrossRef]
- Giménez, M.F.; Tausk, F.; Gigli, I. Langerhans’ Cells in Paracoccidioidomycosis. Arch. Dermatol. 1987, 123, 479–481. [Google Scholar] [CrossRef]
- Pagliari, C.; Sotto, M.N. Correlation of factor XIIIa + dermal dendrocytes with paracoccidioidomycosis skin lesions. Med. Mycol. 2002, 40, 407–410. [Google Scholar] [CrossRef]
- Da Silva, W.L.F.; Pagliari, C.; Duarte, M.I.S.; Sotto, M.N. Paracoccidioides brasiliensis interacts with dermal dendritic cells and keratinocytes in human skin and oral mucosa lesions. Med. Mycol. 2016, 54, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Pagliari, C.; Kanashiro-Galo, L.; Silva, A.A.D.L.; Barboza, T.C.; Criado, P.R.; Duarte, M.I.S.; Brito, A.C.D.; Xavier, M.B.; Unger, D.; Oliveira, C.M.M.; et al. Plasmacytoid dendritic cells in cutaneous lesions of patients with chromoblastomycosis, lacaziosis, and paracoccidioidomycosis: A comparative analysis. Med. Mycol. 2014, 52, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.K.; Bachiega, T.F.; Rodrigues, D.R.; Golim, M.D.A.; Dias-Melicio, L.A.; Balderramas, H.D.A.; Kaneno, R.; Soares, A.M.V.C. Paracoccidioides brasiliensis Interferes on Dendritic Cells Maturation by Inhibiting PGE2 Production. PLoS ONE 2015, 10, e0120948. [Google Scholar] [CrossRef]
- De Castro, L.F.; Longhi, L.N.A.; Paião, M.R.; Justo-Júnior, A.D.S.; De Jesus, M.B.; Blotta, M.H.D.S.L.; Mamoni, R.L. NLRP3 inflammasome is involved in the recognition of Paracoccidioides brasiliensis by human dendritic cells and in the induction of Th17 cells. J. Infect. 2018, 77, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Preite, N.W.; Feriotti, C.; De Lima, D.S.; Da Silva, B.B.; Condino-Neto, A.; Pontillo, A.; Calich, V.L.G.; Loures, F.V. The Syk-Coupled C-Type Lectin Receptors Dectin-2 and Dectin-3 Are Involved in Paracoccidioides brasiliensis Recognition by Human Plasmacytoid Dendritic Cells. Front. Immunol. 2018, 9, 464. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.O.; Favali, C.; Soares, N.C.; Tavares, N.M.; Jerônimo, M.S.; Junior, P.H.V.; Marina, C.L.; Santos, C.; Brodskyn, C.; Bocca, A.L. New Role of P. brasiliensis α-Glucan: Differentiation of Non-conventional Dendritic Cells. Front. Microbiol. 2019, 10, 2445. [Google Scholar] [CrossRef] [PubMed]
- Venturini, J.; Cavalcante, R.D.S.; Golim, M.A.; Marchetti, C.M.; Azevedo, P.Z.D.; Amorim, B.C.; De Arruda, M.S.P.; Mendes, R.P. Phenotypic and functional evaluations of peripheral blood monocytes from chronic-form paracoccidioidomycosis patients before and after treatment. BMC Infect. Dis. 2014, 14, 552. [Google Scholar] [CrossRef] [Green Version]
- Sato, P.; Oshiro, T.M.; Diogo, C.L.; Passos, É.C.; Shikanai-Yasuda, M.A. Characterization of Monocyte-Derived Dendritic Cells from Patients with Active and Treated Paracoccidioidomycosis. Scand. J. Immunol. 2011, 74, 609–618. [Google Scholar] [CrossRef]
- De Camargo, Z.; Unterkircher, C.; Campoy, S.P.; Travassos, L.R. Production of Paracoccidioides brasiliensis exoantigens for immunodiffusion tests. J. Clin. Microbiol. 1988, 26, 2147–2151. [Google Scholar] [CrossRef] [Green Version]
- Camargo, Z.P.; Taborda, C.P.; Rodrigues, E.G.; Travassos, L.R. The use of cell-free antigens of Paracoccidioides brasiliensis in serological tests. J. Med. Vet. Mycol. Bimon. Publ. Int. Soc. Hum. Anim. Mycol. 1991, 29, 31–38. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Romani, N.; Gruner, S.; Brang, D.; Kämpgen, E.; Lenz, A.; Trockenbacher, B.; Konwalinka, G.; Fritsch, P.O.; Steinman, R.M.; Schuler, G. Proliferating dendritic cell progenitors in human blood. J. Exp. Med. 1994, 180, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, J.O.; Ampel, N.M.; Lake, D.F. Reversal of Coccidioidal Anergy In Vitro by Dendritic Cells from Patients with Disseminated Coccidioidomycosis. J. Immunol. 2002, 169, 2020–2025. [Google Scholar] [CrossRef] [PubMed]
- Sadahiro, A.; Diogo, C.L.; Oshiro, T.M.; Shikanai-Yasuda, M.A. Kinetics of IFN-gamma, TNF-alpha, IL-10 and IL-4 production by mononuclear cells stimulated with gp43 peptides, in patients cured of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2007, 40, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, P.K. Monocyte-Derived Dendritic Cells in Human Paracoccidioidomycosis. Unpublished work. 2021. [Google Scholar]
- Amorim, B.C.; Pereira-Latini, A.C.; Golim, M.D.A.; Júnior, R.L.R.; Yoo, H.H.B.; De Arruda, M.S.P.; Tavares, A.H.; Cavalcante, R.D.S.; Mendes, R.P.; Pontillo, A.; et al. Enhanced expression of NLRP3 inflammasome components by monocytes of patients with pulmonary paracoccidioidomycosis is associated with smoking and intracellular hypoxemia. Microbes Infect. 2020, 22, 137–143. [Google Scholar] [CrossRef]
- De Araújo, E.F.; Preite, N.W.; Veldhoen, M.; Loures, F.V.; Calich, V.L.G. Pulmonary paracoccidioidomycosis in AhR deficient hosts is severe and associated with defective Treg and Th22 responses. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Czakai, K.; Dittrich, M.; Kaltdorf, M.; Müller, T.; Krappmann, S.; Schedler, A.; Bonin, M.; Edühring, S.; Schuster, S.; Speth, C.; et al. Influence of Platelet-rich Plasma on the immune response of human monocyte-derived dendritic cells and macrophages stimulated with Aspergillus fumigatus. Int. J. Med. Microbiol. 2017, 307, 95–107. [Google Scholar] [CrossRef]
- Coady, A.; Sil, A. MyD88-Dependent Signaling Drives Host Survival and Early Cytokine Production during Histoplasma capsulatum Infection. Infect. Immun. 2015, 83, 1265–1275. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.P.; Heiss, C.; Black, I.; Azadi, P.; Gerlach, J.Q.; Travassos, L.R.; Joshi, L.; Kilcoyne, M.; Puccia, R. Extracellular vesicles from Paracoccidioides pathogenic species transport polysaccharide and expose ligands for DC-SIGN receptors. Sci. Rep. 2015, 5, srep14213. [Google Scholar] [CrossRef]
- Romano, C.C.; Mendesgiannini, M.J.S.; Duarte, A.J.; Benard, G. The role of interleukin-10 in the differential expression of interleukin-12p70 and its β2 receptor on patients with active or treated paracoccidioidomycosis and healthy infected subjects. Clin. Immunol. 2005, 114, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Arruda, C.; Franco, M.F.; Kashino, S.S.; Nascimento, F.R.F.D.; Fazioli, R.D.A.; Vaz, C.A.; Russo, M.; Calich, V.L.G. Interleukin-12 Protects Mice against Disseminated Infection Caused by Paracoccidioides brasiliensis but Enhances Pulmonary Inflammation. Clin. Immunol. 2002, 103, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Livonesi, M.C.; Souto, J.T.; Campanelli, A.P.; Maffei, C.M.L.; Martinez, R.; Rossi, M.A.; Da Silva, J.S. Deficiency of IL-12p40 subunit determines severe paracoccidioidomycosis in mice. Med. Mycol. 2008, 46, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopera, D.; Naranjo, T.W.; Cruz, O.G.; Restrepo, Á.; Cano, L.E.; Lenzi, H.L. Structural and Topographic Dynamics of Pulmonary Histopathology and Local Cytokine Profiles in Paracoccidioides brasiliensis Conidia-Infected Mice. PLoS Neglected Trop. Dis. 2011, 5, e1232. [Google Scholar] [CrossRef]
- Tristão, F.S.M.; Rocha, F.A.; Carlos, D.; Ketelut-Carneiro, N.; Souza, C.O.S.; Milanezi, C.M.; Silva, J.S. Th17-Inducing Cytokines IL-6 and IL-23 Are Crucial for Granuloma Formation during Experimental Paracoccidioidomycosis. Front. Immunol. 2017, 8, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahara, T.; Urabe, K.; Fukagawa, S.; Uchi, H.; Inaba, K.; Furue, M.; Moroi, Y. Engagement of human monocyte-derived dendritic cells into interleukin (IL)-12 producers by IL-1b + interferon (IFN)-g. Clin. Exp. Immunol. 2005, 139, 476–482. [Google Scholar] [CrossRef]
- Vulcano, M.; Struyf, S.; Scapini, P.; Cassatella, M.; Bernasconi, S.; Bonecchi, R.; Calleri, A.; Penna, G.; Adorini, L.; Luini, W.; et al. Unique Regulation of CCL18 Production by Maturing Dendritic Cells. J. Immunol. 2003, 170, 3843–3849. [Google Scholar] [CrossRef]
- Xander, P.; Vigna, A.F.; Feitosa, L.D.S.; Pugliese, L.; Bailão, A.M.; Soares, C.M.D.A.; Mortara, R.A.; Mariano, M.; Lopes, J.D. A surface 75-kDa protein with acid phosphatase activity recognized by monoclonal antibodies that inhibit Paracoccidioides brasiliensis growth. Microbes Infect. 2007, 9, 1484–1492. [Google Scholar] [CrossRef]
- Schutyser, E.; Richmond, A.; Van Damme, J. Involvement of CC chemokine ligand 18 (CCL18) in normal and pathological processes. J. Leukoc. Biol. 2005, 78, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Rademacher, J.; Kill, A.; Mattat, K.; Dragun, D.; Siegert, E.; Günther, J.; Riemekasten, G. Monocytic Angiotensin and Endothelin Receptor Imbalance Modulate Secretion of the Profibrotic Chemokine Ligand 18. J. Rheumatol. 2016, 43, 587–591. [Google Scholar] [CrossRef]
- Bruna-Romero, O.; Schmieg, J.; Del Val, M.; Buschle, M.; Tsuji, M. The Dendritic Cell-Specific Chemokine, Dendritic Cell-Derived CC Chemokine 1, Enhances Protective Cell-Mediated Immunity to Murine Malaria. J. Immunol. 2003, 170, 3195–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez, A.; Hernández-Pando, R.; Contreras, S.; Aguilar, D.; Rook, G. CCL2, CCL18 and sIL-4R in renal, meningeal and pulmonary TB; a 2 year study of patients and contacts. Tuberculosis 2011, 91, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Kusano, F.; Tanaka, Y.; Marumo, F.; Sato, C. Expression of C-C chemokines is associated with portal and periportal inflammation in the liver of patients with chronic hepatitis C. Lab. Investig. 2000, 80, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, C.O.; Fliesser, M.; Dittrich, M.; Mueller, T.; Bauer, R.; Kneitz, S.; Hope, W.; Rogers, T.R.; Einsele, H.; Loeffler, J. Gene Expression Profiles of Human Dendritic Cells Interacting with Aspergillus fumigatus in a Bilayer Model of the Alveolar Epithelium/Endothelium Interface. PLoS ONE 2014, 9, e98279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hector, A.; Kröner, C.; Carevic, M.; Bakele, M.; Rieber, N.; Riethmüller, J.; Griese, M.; Zissel, G.; Hartl, D. The chemokine CCL18 characterises Pseudomonas infections in cystic fibrosis lung disease. Eur. Respir. J. 2014, 44, 1608–1615. [Google Scholar] [CrossRef]
- Rong, B.; Fu, T.; Rong, C.; Liu, W.; Li, K.; Liu, H. Association between serum CCL-18 and IL-23 concentrations and disease progression of chronic obstructive pulmonary disease. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Korbecki, J.; Olbromski, M.; Dziegiel, P. CCL18 in the Progression of Cancer. Int. J. Mol. Sci. 2020, 21, 7955. [Google Scholar] [CrossRef]
- Bosco, M.C.; Puppo, M.; Santangelo, C.; Anfosso, L.; Pfeffer, U.; Fardin, P.; Battaglia, F.; Varesio, L. Hypoxia Modifies the Transcriptome of Primary Human Monocytes: Modulation of Novel Immune-Related Genes and Identification Of CC-Chemokine Ligand 20 as a New Hypoxia-Inducible Gene. J. Immunol. 2006, 177, 1941–1955. [Google Scholar] [CrossRef]
- Benard, G.; Romano, C.C.; Cacere, C.R.; Juvenale, M.; Mendes-Giannini, M.J.S.; Duarte, A.J. Imbalance of IL-2, IFN-γ and IL-10 Secretion in the Immunosuppression Associated with Human Paracoccidioidomycosis. Cytokine 2001, 13, 248–252. [Google Scholar] [CrossRef]
- Panunto-Castelo, A.; Freitas-Da-Silva, G.; Bragheto, I.C.; Martinez, R.; Roque-Barreira, M.C. Paracoccidioides brasiliensis exoantigens: Recognition by IgG from patients with different clinical forms of paracoccidioidomycosis. Microbes Infect. 2003, 5, 1205–1211. [Google Scholar] [CrossRef]
- Cavassani, K.A.; Tristao, F.S.M.; De Oliveira, L.L.; Rocha, F.A.; Vancim, J.O.; Moreira, A.P.; Campanelli, A.P.; Panagio, L.A.; Milanezi, C.M.; Martinez, R.; et al. Cell-Free Antigens from Paracoccidioides brasiliensis Drive IL-4 Production and Increase the Severity of Paracoccidioidomycosis. PLoS ONE 2011, 6, e21423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mello, L.M.; Silva-Vergara, M.L.; Junior, V.R.; Junior, V.R. Patients with active infection with Paracoccidioides brasiliensis present a Th2 immune response characterized by high Interleukin-4 and Interleukin-5 production. Hum. Immunol. 2002, 63, 149–154. [Google Scholar] [CrossRef]
- Marchant, A.; Amedei, A.; Azzurri, A.; Vekemans, J.; Benagiano, M.; Tamburini, C.; Lienhardt, C.; Corrah, T.; McAdam, K.P.W.J.; Romagnani, S.; et al. Polarization of PPD-Specific T-Cell Response of Patients with Tuberculosis from Th0 to Th1 Profile after Successful Antimycobacterial Therapy or In Vitro Conditioning with Interferon- α or Interleukin-12. Am. J. Respir. Cell Mol. Biol. 2001, 24, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Arruda, C.; Valente, R.; Pina, A.; Kashino, S.S.; Fazioli, R.A.; Vaz, C.A.C.; Franco, M.F.; Keller, A.C.; Calich, V.L.G. Dual Role of Interleukin-4 (IL-4) in Pulmonary Paracoccidioidomycosis: Endogenous IL-4 Can Induce Protection or Exacerbation of Disease Depending on the Host Genetic Pattern. Infect. Immun. 2004, 72, 3932–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Costa, T.A.; Bazan, S.B.; Feriotti, C.; De Araujo, E.F.; Bassi, Ê.J.; Loures, F.V.; Calich, V.L.G. In Pulmonary Paracoccidioidomycosis IL-10 Deficiency Leads to Increased Immunity and Regressive Infection without Enhancing Tissue Pathology. PLoS Neglected Trop. Dis. 2013, 7, e2512. [Google Scholar] [CrossRef] [PubMed]
- Jannuzzi, G.P.; Tavares, A.H.F.P.; Kaihami, G.H.; De Almeida, J.R.F.; De Almeida, S.R.; Ferreira, K.S. scFv from Antibody That Mimics gp43 Modulates the Cellular and Humoral Immune Responses during Experimental Paracoccidioidomycosis. PLoS ONE 2015, 10, e0129401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.B.R.; Taira, C.L.; Dias, L.S.; Souza, A.C.O.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Experimental Therapy of Paracoccidioidomycosis Using P10-Primed Monocyte-Derived Dendritic Cells Isolated from Infected Mice. Front. Microbiol. 2019, 10, 1727. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, P.K.; Oshiro, T.M.; Passos, É.C.; Miranda, T.G.R.; Diogo, C.L.; Fonseca, C.d.A.; Sadahiro, A.; de Almeida, S.R.; Shikanai-Yasuda, M.A. Monocyte-Derived Dendritic Cells Can Revert In Vitro Antigen-Specific Cellular Anergy in Active Human Paracoccidioidomycosis. J. Fungi 2021, 7, 201. https://doi.org/10.3390/jof7030201
Sato PK, Oshiro TM, Passos ÉC, Miranda TGR, Diogo CL, Fonseca CdA, Sadahiro A, de Almeida SR, Shikanai-Yasuda MA. Monocyte-Derived Dendritic Cells Can Revert In Vitro Antigen-Specific Cellular Anergy in Active Human Paracoccidioidomycosis. Journal of Fungi. 2021; 7(3):201. https://doi.org/10.3390/jof7030201
Chicago/Turabian StyleSato, Paula Keiko, Telma Miyuki Oshiro, Érika Cano Passos, Tatiana Giselle Rodrigues Miranda, Constância Lima Diogo, Claudia de Abreu Fonseca, Aya Sadahiro, Sandro Rogério de Almeida, and Maria Aparecida Shikanai-Yasuda. 2021. "Monocyte-Derived Dendritic Cells Can Revert In Vitro Antigen-Specific Cellular Anergy in Active Human Paracoccidioidomycosis" Journal of Fungi 7, no. 3: 201. https://doi.org/10.3390/jof7030201
APA StyleSato, P. K., Oshiro, T. M., Passos, É. C., Miranda, T. G. R., Diogo, C. L., Fonseca, C. d. A., Sadahiro, A., de Almeida, S. R., & Shikanai-Yasuda, M. A. (2021). Monocyte-Derived Dendritic Cells Can Revert In Vitro Antigen-Specific Cellular Anergy in Active Human Paracoccidioidomycosis. Journal of Fungi, 7(3), 201. https://doi.org/10.3390/jof7030201