
Epidemiological Studies of Brown Rot in Spanish Cherry Orchards in the Jerte Valley




Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchard and Experimental Design
2.2. Determination of Pathogen Primary Inoculum Sources
2.3. Brown-Rot Evaluation
2.4. Monitoring of Environmental Variables
2.5. Data Analysis
3. Results
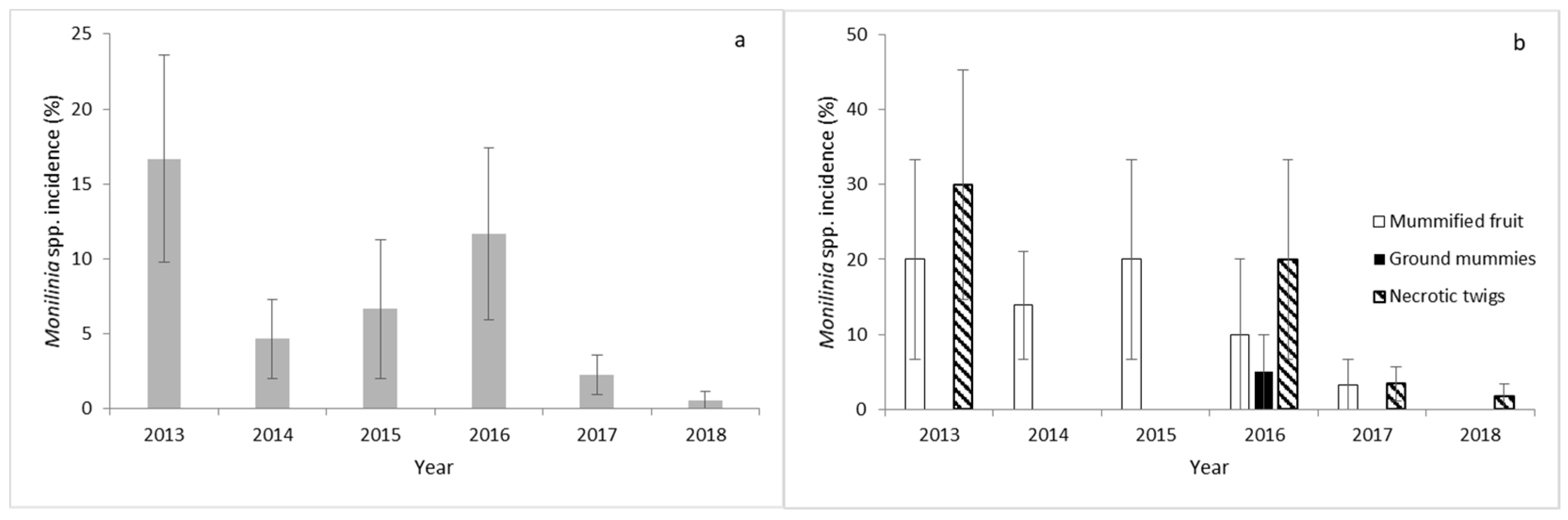
3.1. Pathogen Primary Inoculum Sources
3.2. Climatic Condition Effects on Primary Inoculum
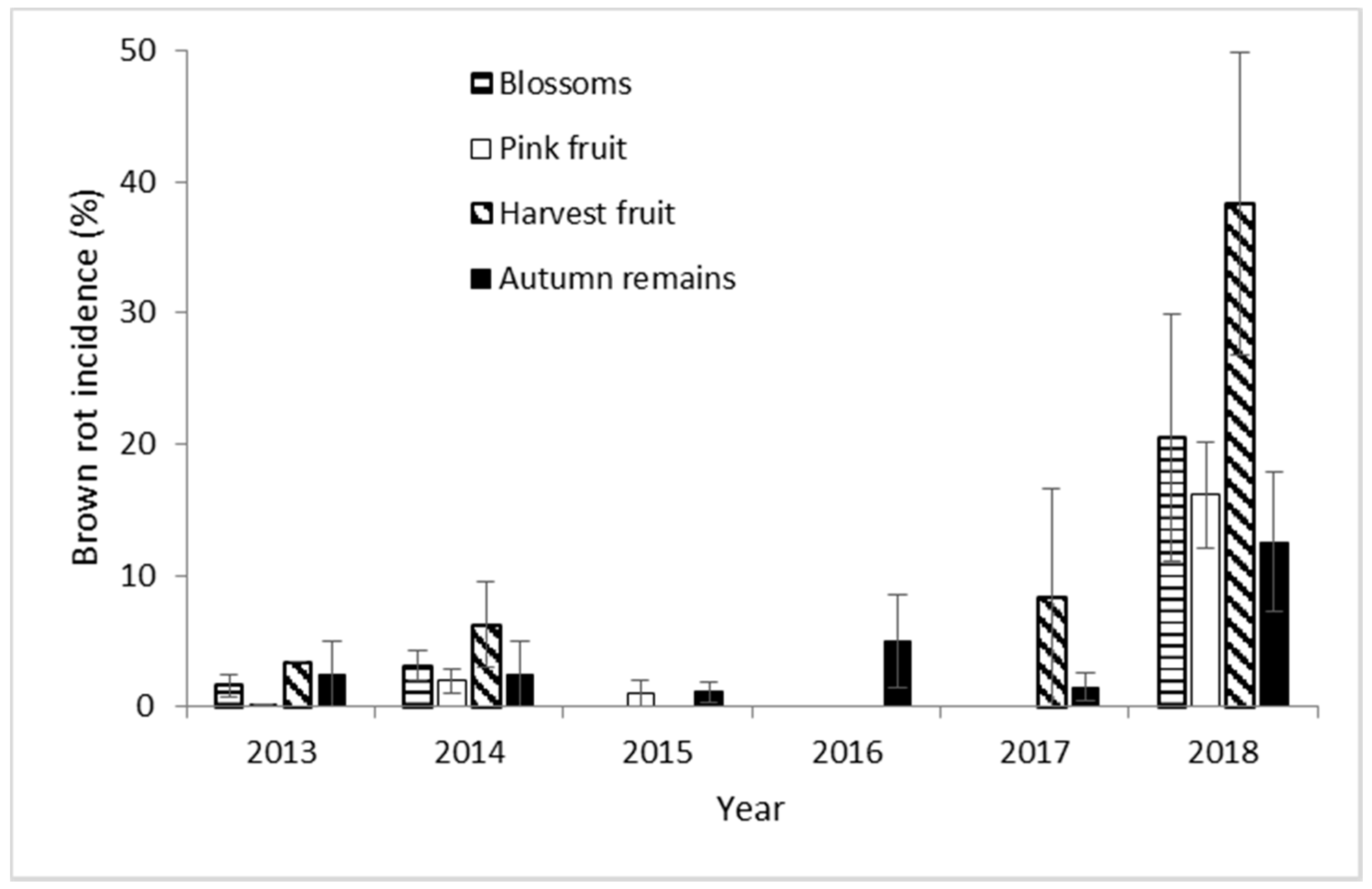
3.3. Cherry Brown Rot Development
3.4. Effects of Climatic Conditions on Cherry Brown Rot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Becedas, M.T. Enfermedades más frecuentes del cerezo en Extremadura. Vida Rural 2011, 338, 51–55. [Google Scholar]
- Byrde, R.J.W.; Willetts, H.J. The Brown Rot Fungi of Fruit: Their Biology and Control; Pergamon Press: Oxford, UK, 1977. [Google Scholar]
- Xu, X.; Bertone, C.; Berrie, A. Effects of wounding, fruit age and wetness duration on the development of cherry brown rot in the UK. Plant Pathol. 2007, 56, 114–119. [Google Scholar] [CrossRef]
- Anuario Estadistica Agraria MAPA. Available online: http://www.mapa.gob.es/es/ (accessed on 20 September 2019).
- Holb, I.J.; Szőke, S.; Abonyi, F. Temporal development and relationship amongst brown rot blossom blight, fruit blight and fruit rot in integrated and organic sour cherry orchards. Plant Pathol. 2013, 62, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Corbin, J.B.; Ogawa, J.M.; Schultz, H.B. Fluctuations in numbers of Monilinia laxa conidia in an apricot orchard during the 1966 season. Phytopathology 1968, 58, 1387–1394. [Google Scholar]
- Holb, I.J. Monitoring conidial density of Monilinia fructigena in the air in relation to brown rot development in integrated and organic apple orchards. Eur. J. Plant Pathol. 2008, 120, 397–408. [Google Scholar] [CrossRef]
- Biggs, A.R.; Northover, J. Early and late-season susceptibility of peach fruits to Monilinia fructicola. Plant Dis. 1988, 72, 1070–1074. [Google Scholar] [CrossRef]
- Phillips, D.J. Effect of temperature on Monilinia fructicola spores produced on fresh stone fruits. Plant Dis. 1984, 68, 610–612. [Google Scholar] [CrossRef]
- Bannon, F.; Gort, G.; Van Leeuwen, G.; Holb, I.; Jeger, M. Diurnal patterns in dispersal of Monilinia fructigena conidia in an apple orchard in relation to weather factors. Agric. For. Meteorol. 2009, 149, 518–525. [Google Scholar] [CrossRef]
- Gell, I.; De Cal, A.; Torres, R.; Usall, J.; Melgarejo, P. Conidial density of Monilinia spp. on peach fruit surfaces in relation to the incidences of latent infections and brown rot. Eur. J. Plant Pathol. 2009, 123, 415–424. [Google Scholar] [CrossRef]
- Landgraf, F.A.; Zehr, E.I. Inoculum sources for Monilinia fructicola in South Carolina peach orchards. Phytopathology 1982, 72, 185–190. [Google Scholar] [CrossRef]
- Watson, W.A.; Zehr, E.I.; Grimes, L.W. Influence of temperature and wetting period on inoculum production by Monilinia fructicola in peach twig cankers. Plant Dis. 2002, 86, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.P.; Bertolini, P. Effect of temperature during conidial formation of Monilinia laxa on conidial size, germination and infection of stored nectarines. J. Phytopathol. Phytopathol. 1999, 147, 635–641. [Google Scholar] [CrossRef]
- Wilcox, W.F. Influence of environment and inoculum density on the incidence of brown rot blossom blight of sour cherry. Phytopathology 1989, 79, 530–534. [Google Scholar] [CrossRef]
- Tamm, L.; Minder, C.E.; Fluckiger, W. Phenological Analysis of brown-rot blossom blight of sweet cherry caused by Monilinia laxa. Phytopathology 1995, 85, 401–408. [Google Scholar] [CrossRef]
- Xu, X.M.; Guerin, L.; Robinson, J.D. Effects of temperature and relative humidity on conidial germination and viability, colonization and sporulation of Monilinia fructigena. Plant Pathol. 2001, 50, 561–568. [Google Scholar] [CrossRef]
- Holb, I.J. Harvest and postharvest brown rot of fruit in relation to early latent infection caused by Monilinia spp. in Hungary. Int. J. Hortic. Sci. 2015, 21, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Gell, I.; De Cal, A.; Torres, R.; Usall, J.; Melgarejo, P. Relationship between the incidence of latent infections caused by Monilinia spp. and the incidence of brown rot of peach fruit: Factors affecting latent infection. Eur. J. Plant Pathol. 2008, 121, 487–498. [Google Scholar] [CrossRef]
- Luo, Y.; Michailides, T.J. Factors affecting latent infection of prune fruit by Monilinia fructicola. Phytopathology 2001, 91, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Boletin Fitosanitario de Avisos e Informaciones Junta de Extremadura Servicio de Sanidad Vegetal. Dirección Programas P. Integrada, Diagnósticos y Avisos Agrícolas. Estación de Avisos Agrícolas. 2017. Año: XLV Boletín No: 8. Available online: http://www.juntaex.es/filescms/con03/uploaded_files/SectoresTematicos/Agricultura/SanidadVegetal/Boletinfitosanitario/2017/Boletin_n8_2017.pdf (accessed on 11 April 2017).
- De Cal, A.; Melgarejo, P. Effects of long-wave UV light on Monilinia growth and identification of species. Plant Dis. 1999, 83, 62–65. [Google Scholar] [CrossRef] [Green Version]
- Gell, I.; Cubero, J.; Melgarejo, P. Two different PCR approaches for universal diagnosis of brown rot and identification of Monilinia spp. in stone fruit trees. J. Appl. Microbiol. 2007, 103, 2629–2637. [Google Scholar] [CrossRef]
- Garcia-Benitez, C.; Casals, C.; Usall, J.; Sánchez-Ramos, I.; Melgarejo, P.; De Cal, A. Impact of Postharvest Handling on Preharvest Latent Infections Caused by Monilinia spp. in Nectarines. J. Fungi 2020, 6, 266. [Google Scholar] [CrossRef] [PubMed]
- Snedecor, G.; Cochram, W.G. Statistical Methods; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Van Leeuwen, G.C.M.; Stein, A.; Holb, I.; Jeger, M.J. Yield loss in apple caused by Monilinia fructigena (Aderh. & Ruhl.) Honey, and spatio-temporal dynamics of disease development. Eur. J. Plant Pathol. 2000, 106, 519–528. [Google Scholar]
- Xu, X.M.; Robinson, J.D. Epidemiology of brown rot (Monilinia fructigena) on apple: Infection of fruits by conidia. Plant Pathol. 2000, 49, 201–206. [Google Scholar] [CrossRef]
- Ogawa, J.M.; Zehr, E.I.; Biggs, A.R. Brown rot. In Compendium of Stone Fruit Disease; Ogawa, J.M., Zehr, E.I., Bird, G.W., Ritchie, D.F., Uriu, K., Uyemoto, J.K., Eds.; American Phytopathological Society: St. Paul, MN, USA, 1995. [Google Scholar]
- Stensvand, A.; Talgø, V.; Børve, J. Seasonal production of conidia of Monilinia laxa from mummified fruits, blighted spurs, and flowers of sweet cherry. Gartenbauwissenschaft 2001, 66, 273–281. [Google Scholar]
- Tamm, L. Epidemiological Aspects of Sweet Cherry Blossom Blight Caused by Monilinia laxa. Ph.D. Thesis, University of Basel, Schonenbuch, Switzerland, 1994. [Google Scholar]
- Thomidis, T. Influence of relative virulence and latent infections on the development of Monilinia to Greek peach orchards. Crop Prot. 2017, 94, 159–165. [Google Scholar] [CrossRef]
- Everhart, S.E.; Askew, A.; Seymour, L.; Holb, I.J.; Scherm, H. Characterization of three-dimensional spatial aggregation and association patterns of brown rot symptoms within intensively mapped sour cherry trees. Ann. Bot. 2011, 108, 1195–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauvert, P.; Fournet, J.; Rapilly, F. Etudes sur la dispersion d’un inoculum par des gouttes d’eau en fonction du conceptacle sporife’re. Ann. Phytopathol. 1969, 1, 491–493. [Google Scholar]
- Garcia-Benitez, C.; Melgarejo, P.; De Cal, A. Fruit maturity and post-harvest environmental conditions influence the pre-penetration stages of Monilinia infections in peaches. Int. J. Food Microbiol. 2016, 10, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casals, C.; Vinas, I.; Torres, R.; Griera, C.; Usall, J. Effect of temperature and water activity on in vitro germination of Monilinia spp. J. Appl. Microbiol. 2010, 108, 47–54. [Google Scholar] [CrossRef]
- Koball, D.C.; Wilcox, W.F.; Seem, R.C. Influence of incubation—Period humidity on the development of brown rot blossom blight of sour cherry. Phytopathology 1997, 87, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Tamm, L.; Fluckiger, W. Influence of temperature and moisture on growth, spore production, and conidial germination of Monilinia laxa. Phytopathology 1993, 83, 1321–1326. [Google Scholar] [CrossRef]
- Simeone, A.M.; Corazza, L. Susceptibility of some stone fruit cultivars to brown rot. Ann. Ist. Sper. Fruttic. 1986, 27, 55–60. [Google Scholar]
- Villarino, M.; Melgarejo, P.; Usall, J.; Segarra, J.; Lamarca, N.; De Cal, A. Secondary inoculum dynamics of Monilinia spp. and relationship to the incidence of postharvest brown rot in peaches and the weather conditions during the growing season. Eur. J. Plant Pathol. 2012, 133, 585–598. [Google Scholar] [CrossRef]
- Villarino, M.; Sandin-España, P.; Melgarejo, P.; De Cal, A. High chlorogenic and neochlorogenic acid levels in immature peaches reduce Monilinia laxa infection by Interfering with Fungal Melanin Biosynthesis. J. Agric. Food Chem. 2011, 59, 3205–3213. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ma, Z.; Michailides, T.J. Analysis of factors affecting latent infection and sporulation of Monilinia fructicola on prune fruit. Plant Dis. 2001, 85, 999–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Period | Mean T (°C) | Mean RH (%) | Dew Point (°C) | Primary Inoculum Observed (%) | Primary Inoculum (%) Estimated by Model (1) |
|---|---|---|---|---|---|
| 2013–14 | 7.52 | 76.02 | 2.93 | 4.65 | 4.32 |
| 2014–15 | 8.60 | 59.35 | 0.12 | 6.67 | 7.95 |
| 2015–16 | 9.44 | 74.81 | 4.88 | 11.6 | 10.79 |
| 2016–17 | 6.91 | 78.65 | 3.22 | 2.25 | 2.26 |
| 2017–18 | 6.36 | 74.84 | 2.02 | 0.56 | 0.41 |
| r 1 | 0.98 | −0.27 | 0.39 | ||
| p-value 1 | 0.003 | 0.660 | 0.519 |
| Period | Mean T (°C) | Primary Inoculum (%) Estimated by Model (1) |
|---|---|---|
| 2012–2013 | 7.75 | 5.09 |
| 2013–2014 | 8.00 | 5.95 |
| 2014–2015 | 7.83 | 5.37 |
| 2015–2016 | 9.28 | 10.24 |
| 2016–2017 | 8.89 | 8.94 |
| 2017–2018 | 7.80 | 5.26 |
| 2018–2019 | 8.58 | 7.88 |
| 2019–2020 | 9.34 | 10.47 |
| Year | Mean T (°C) | Mean RH (%) | MCD-RH ≥ 80 | Annual BR Observed (%) | BR Observed at Harvest (%) | BR Estimated at Harvest (%) by Model (2) |
|---|---|---|---|---|---|---|
| 2013 | 11.58 | 70.96 | 5.00 | 1.70 | 3.39 | 16.93 |
| 2014 | 13.36 | 63.44 | 3.75 | 3.79 | 6.25 | 8.17 |
| 2015 | 13.05 | 57.60 | 1.50 | 0.50 | 0.00 | 0.00 |
| 2016 | 10.12 | 65.09 | 1.50 | 0.00 | 0.00 | 0.00 |
| 2017 | 12.86 | 58.59 | 2.75 | 2.77 | 8.30 | 2.97 |
| 2018 | 9.47 | 73.89 | 6.50 | 24.97 | 38.30 | 30.74 |
| r 2 | −0.57 | 0.63 | 0.86 | 0.99 | 0.87 | |
| p-value 2 | 0.23 | 0.17 | 0.02 | 0.0001 | 0.02 |
| MCD-RH > 80% in Each Fortnight | BR (%) Estimated by Model (2) |
|---|---|
| 0 | 0.0 |
| 1 | 0.0 |
| 2 | 0.12 |
| 3 | 4.12 |
| 4 | 9.72 |
| 5 | 16.93 |
| 6 | 25.74 |
| 7 | 36.14 |
| 8 | 48.15 |
| 9 | 61.76 |
| 10 | 76.98 |
| 11 | 93.79 |
| 12 | 100.0 |
| 13 | 100.0 |
| 14 | 100.0 |
| 15 | 100.0 |
| Year | Mean T (°C) | Mean RH (%) | MCD-RH ≥ 80 | R (mm) | MD-R ≥10 | Wind Speed (m/s) | BR (%) Estimated by Model (2) |
|---|---|---|---|---|---|---|---|
| 2013 | 11.25 | 67.07 | 5 | 8.00 | 3.75 | 1.74 | 16.91 |
| 2014 | 13.71 | 59.59 | 2.5 | 3.22 | 1.75 | 1.62 | 1.91 |
| 2016 | 10.81 | 63.61 | 3.5 | 4.40 | 2.25 | 1.74 | 6.71 |
| 2017 | 14.36 | 50.47 | 0.75 | 2.49 | 1 | 1.69 | 0.00 |
| 2018 | 10.83 | 69.02 | 4.75 | 8.79 | 4.25 | 2.08 | 14.96 |
| 2020 | 12.47 | 70.77 | 5.5 | 4.34 | 3 | 1.44 | 21.11 |
| r 2 | −0.60 | 0.91 | 0.95 | 0.66 | 0.86 | −0.015 | |
| p-value 2 | 0.20 | 0.01 | 0.003 | 0.15 | 0.037 | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larena, I.; Villarino, M.; Melgarejo, P.; Cal, A.D. Epidemiological Studies of Brown Rot in Spanish Cherry Orchards in the Jerte Valley. J. Fungi 2021, 7, 203. https://doi.org/10.3390/jof7030203
Larena I, Villarino M, Melgarejo P, Cal AD. Epidemiological Studies of Brown Rot in Spanish Cherry Orchards in the Jerte Valley. Journal of Fungi. 2021; 7(3):203. https://doi.org/10.3390/jof7030203
Chicago/Turabian StyleLarena, Inmaculada, Maria Villarino, Paloma Melgarejo, and Antonieta De Cal. 2021. "Epidemiological Studies of Brown Rot in Spanish Cherry Orchards in the Jerte Valley" Journal of Fungi 7, no. 3: 203. https://doi.org/10.3390/jof7030203
APA StyleLarena, I., Villarino, M., Melgarejo, P., & Cal, A. D. (2021). Epidemiological Studies of Brown Rot in Spanish Cherry Orchards in the Jerte Valley. Journal of Fungi, 7(3), 203. https://doi.org/10.3390/jof7030203