
A Medium-Term Field Experiment to Study the Effect of Managing Soil Chemical Properties on Fusarium Wilt in Banana (Musa AAA)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Setup
2.3. Plant Inoculation
2.4. Soil Management
2.5. Plant Monitoring
2.6. Statistical Analysis
3. Results
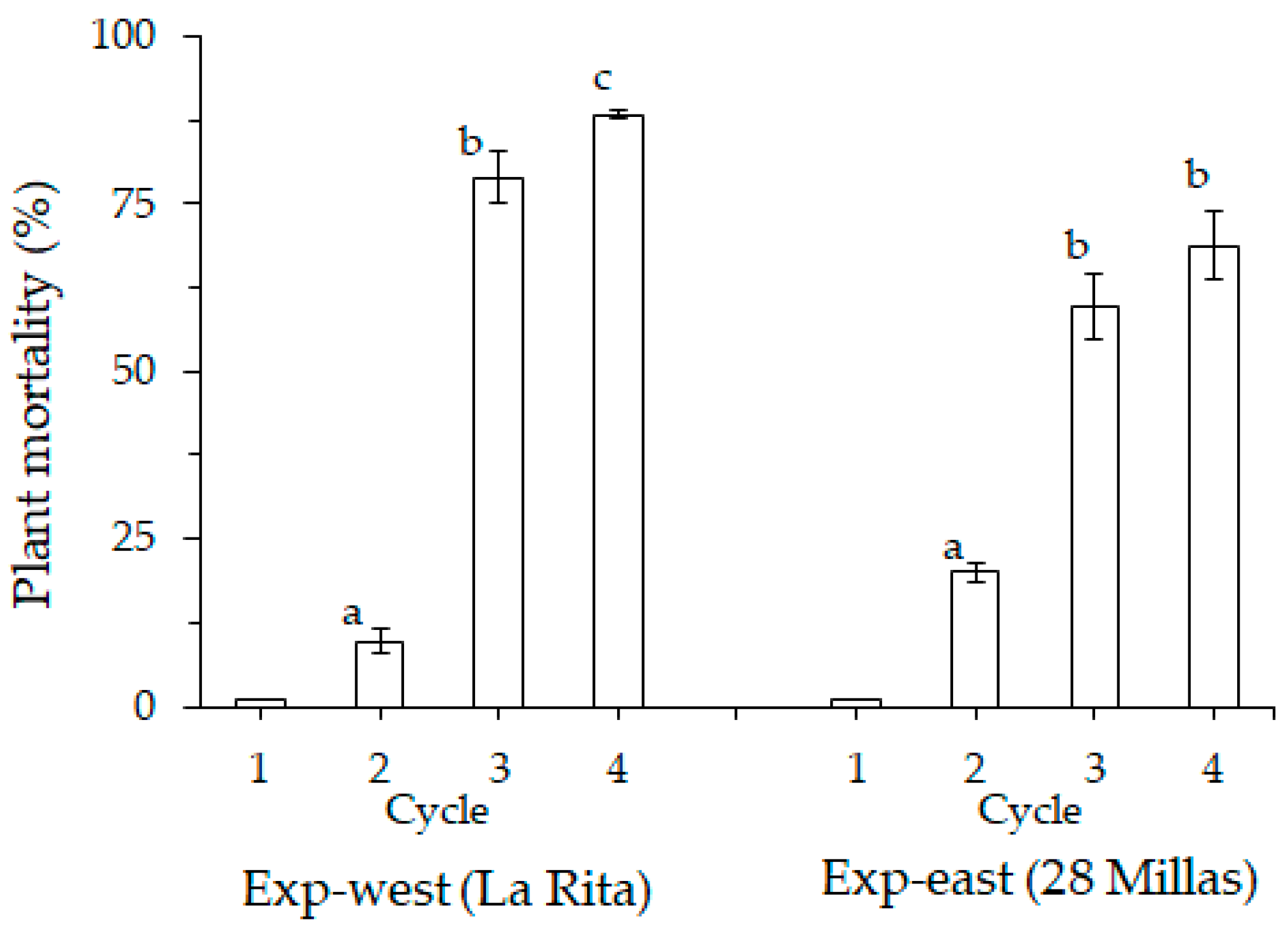
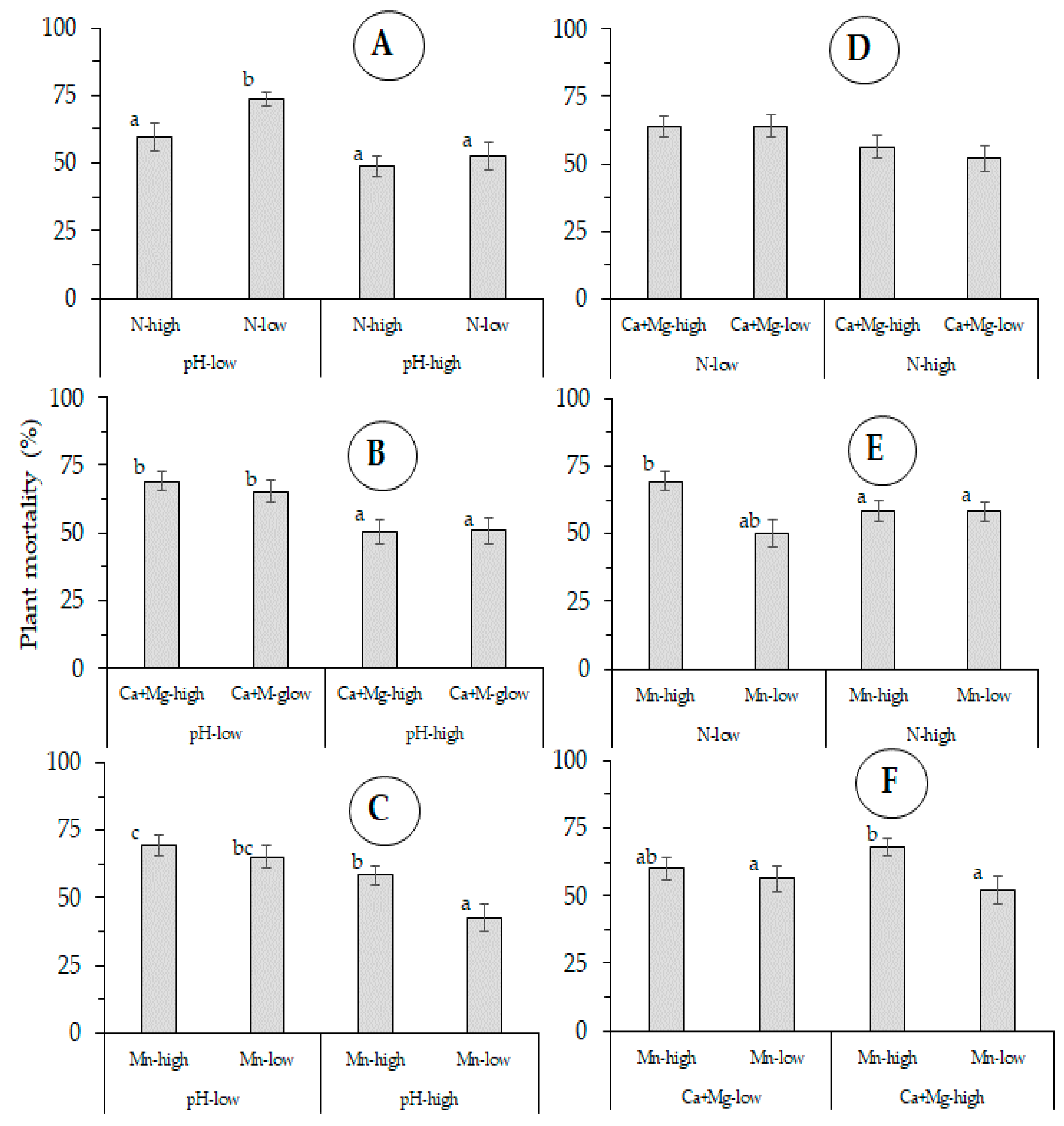
3.1. Plant Mortality
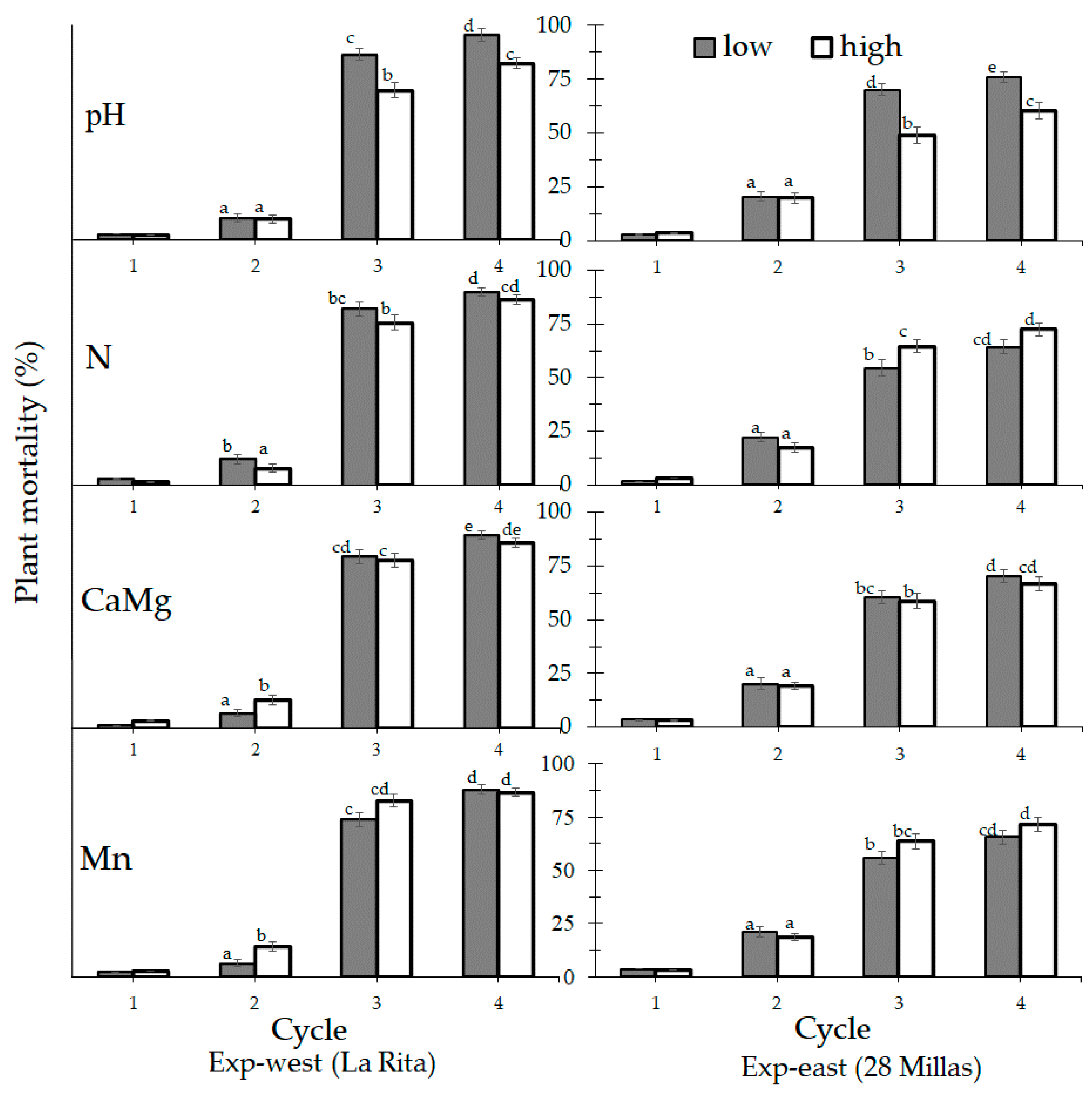
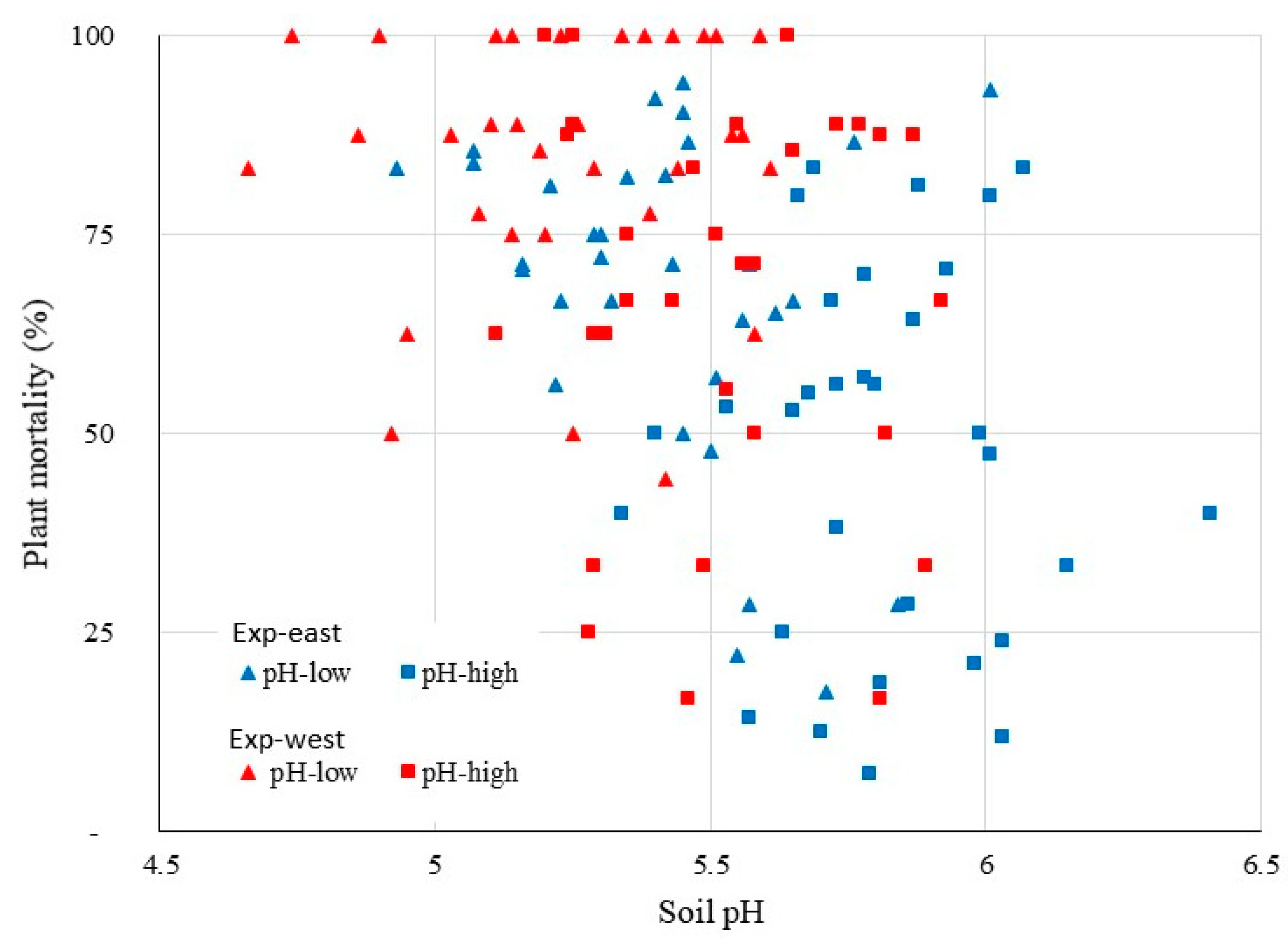
3.2. Analysis for Managed Soil Chemical Properties
3.3. Leaf Nutrient Concentration
3.4. Agronomic Performance
4. Discussion
4.1. Plant Mortality
4.2. Soil Analysis for Tested Soil Properties
4.3. Leaf Nutrient Concentration According to Soil Management
4.4. Agronomic Performance According to Soil Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.-C.; Dehne, H.-W. Safeguarding production—Losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2005, 144, 31. [Google Scholar] [CrossRef]
- Savary, S.; Ficke, A.; Aubertot, J.-N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Stover, R.H. Fusarial Wilt (Panama Disease) of Bananas and Other Musa Species; The Common wealth Mycological Institute: Kew Surrey, UK, 1962. [Google Scholar]
- Ploetz, R.C. Fusarium wilt of banana. Phytopathology 2015, 105, 1512–1521. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. Cubense. Phytopathology 2006, 96, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.F.; Morrison, P.C. Evolution of the banana industry of Costa Rica. Econ. Geogr. 1952, 28, 1–19. [Google Scholar] [CrossRef]
- Harper, J.L. Studies in the resistance of certain varieties of banana to panama disease. Plant Soil 1950, 2, 383–394. [Google Scholar] [CrossRef]
- Ploetz, R.C. Vegetative compatibility in Fusarium Oxysporum f.sp.cubense: Classifying previosly noncharacterized strains. Acta Hortic. 1990, 699–706. [Google Scholar] [CrossRef]
- Perez-Vicente, L. Fusarium wilt (Panama disease) of bananas: An updating review of the current knowledge on the disease and its causal agent. Fitosanidad 2004, 8, 27–38. [Google Scholar]
- Escobedo, A. Cadena Productiva de Banano Criollo (‘Gros Michel’) de Costa Rica; CATIE: Turrialba, Costa Rica, 2010. [Google Scholar]
- Soto, M. Situación y avances tecnologicos en la producción bananera mundial. Rev. Bras. Frutic. 2011, 33, 13–28. [Google Scholar] [CrossRef]
- Aurore, G.; Parfait, B.; Fahrasmane, L. Bananas, raw materials for making processed food products. Trends Food Sci. Technol. 2009, 20, 78–91. [Google Scholar] [CrossRef]
- Buddenhagen, I. Understanding strain diversity in Fusarium oxysporum f. Sp. Cubense and history of introduction of “Tropical race 4” to better manage banana production. Acta Hortic. 2009, 828, 193–204. [Google Scholar] [CrossRef]
- Stover, R.H.; Thornton, N.C.; Dunlap, V.C. Flood-fallowing for eradications of Fusarium Oxysporum F. Cubense: I. Effect of flooding on fungus flora of clay loam soils in Ulua Valley, Honduras. Soil Sci. 1953, 76, 225–238. [Google Scholar] [CrossRef]
- Stover, R.H. Studies on Fusarium wilt of bananas. I. field control. Can. J. Bot. 1961, 39, 197–206. [Google Scholar] [CrossRef]
- Rishbeth, J. Fusarium wilt of bananas in Jamaica. II. Some aspects of host-parasite relationships. Ann. Bot. NS 1955, XXI, 215–245. [Google Scholar]
- Borges, A.; Trujillo JDC, I.; Gutierrez, F.; Angulo, D. Estudio sonre el Mal de Panamá en las Islas Canarias. II.—Influencia de los mecanismos de resistencia de la plantera (Cavendish enana) al Mal de Panamá. Fruits 1983, 38, 755–758. [Google Scholar]
- Domínguez, J.; Negrín, M.A.; Rodríguez, C.M. Soil potassium indices and clay-sized particles affecting banana-wilt expression caused by soil fungus in banana plantation development on transported volcanic soils. Commun. Soil Sci. Plant Anal. 2008, 39, 397–412. [Google Scholar] [CrossRef]
- Orr, R.; Nelson, P.N. Impacts of soil abiotic attributes on Fusarium wilt, focusing on bananas. Appl. Soil Ecol. 2018, 132, 20–33. [Google Scholar] [CrossRef]
- Segura, R.A.; Stoorvogel, J.J.; Bosman, M.; Sandoval, J.A. Fusarium wilt (Foc race 1) in relation to soil properties in smallholder´s farms with ‘Gros Michel’ banana (Musa AAA) in Costa Rica. CORBANA 2019, 45, 67–82. [Google Scholar]
- Segura, R.A.; Stoorvogel, J.J.; Samuels, J.Z.; Sandoval, J.A. Managing the interactions between soil abiotic factors to alleviate the effect of Fusarium wilt in bananas. Acta Hortic. 2018, 163–168. [Google Scholar] [CrossRef]
- Doran, J.W.; Sarrantonio, M.; Liebig, M.A. Soil health and sustainability. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: San Diego, CA, USA, 1996; Volume 56, pp. 1–54. ISBN 978-0-12-000756-1. [Google Scholar]
- Ghorbani, R.; Wilcockson, S.; Koocheki, A.; Leifert, C. Soil management for sustainable crop disease control: A review. Environ. Chem. Lett. 2008, 6, 149–162. [Google Scholar] [CrossRef]
- Huber, D.; Römheld, V.; Weinmann, M. Relationship between nutrition, plant diseases and pests. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 283–298. ISBN 978-0-12-384905-2. [Google Scholar]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Amtmann, A.; Troufflard, S.; Armengaud, P. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant. 2008, 133, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Oritsejafor, J.J. Influence of moisture and pH on growth and survival of Fusarium oxysporum f.sp. elaeidis in soil. Trans. Br. Mycol. Soc. 1986, 87, 511–517. [Google Scholar] [CrossRef]
- Höper, H.; Alabouvette, C. Importance of Physical and chemical properties in the suppressiveness of soils to plant diseases. Eur. J. Soil Biol. 1996, 32, 41–58. [Google Scholar]
- Harrison, U.J.; Shew, H.D. Effects of soil pH and nitrogen fertility on the population dynamics of Thielaviopsis basicola. Plant Soil 2001, 228, 147–155. [Google Scholar] [CrossRef]
- Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Mateille, T.; Steinberg, C. Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biol. Biochem. 2007, 39, 1–23. [Google Scholar] [CrossRef]
- Sugimoto, T.; Watanabe, K.; Yoshida, S.; Aino, M.; Matsuyama, M.; Maekawa, K.; Irie, K. The Effects of inorganic elements on the reduction of phytophthora stem rot disease of soybean, the growth rate and zoospore release of phytophthora sojae. J. Phytopathol. 2007, 155, 97–107. [Google Scholar] [CrossRef]
- Serrano, M.S.; Fernández-Rebollo, P.; Vita, P.; Sánchez, M.E. Calcium mineral nutrition increases the tolerance of quercus ilex to phytophthora root disease affecting oak rangeland ecosystems in Spain. Agrofor. Syst. 2012, 87, 173–179. [Google Scholar] [CrossRef]
- Huber, D.M.; Jones, J.B. The role of magnesium in plant disease. Plant Soil 2012, 368, 73–85. [Google Scholar] [CrossRef]
- Brennan, R.F. The role of manganese and nitrogen nutrition in the susceptibility of wheat plants to take-all in Western Australia. Fertil. Res. 1992, 31, 35–41. [Google Scholar] [CrossRef]
- Stoorvogel, J.J.; Segura, R.A. Nutrition and soil management in banana cultivation. In Achieving Sustainable Cuntivations of Banana, Volume 1: Cultivation Techniques; Kema, G.H.J., Drenth, A., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 223–234. ISBN 978-1-78676-156-9. [Google Scholar]
- Gibson, D.J.; Connolly, J.; Hartnett, D.C.; Weidenhamer, J.D. Designs for greenhouse studies of interactions between plants. J. Ecol. 1999, 87, 1–16. [Google Scholar] [CrossRef]
- Tripp, R.; Woolley, J. The Planning Stage of on-Farm Research: Identifying Factors for Experimentation; CIMMYT: Mexico City, Mexico; CIAT: Cali, Colombia, 1989; ISBN 978-968-6127-02-7. [Google Scholar]
- Dale, J.; Paul, J.-Y.; Dugdale, B.; Harding, R. Modifying bananas: From transgenics to organics? Sustainability 2017, 9, 333. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.-C.; Ko, W.-H. Cavendish Banana cultivars resistant to fusarium wilt acquired through somaclonal variation in Taiwan. Plant Dis. 2004, 88, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Segura-Mena, R.A.; Stoorvogel, J.J.; García-Bastidas, F.; Salacinas-Niez, M.; Kema, G.H.J.; Sandoval, J.A. Evaluating the potential of soil management to reduce the effect of Fusarium oxysporum f. sp. cubense in banana (Musa AAA). Eur. J. Plant Pathol. 2021. [Google Scholar] [CrossRef]
- García-Bastidas, F.A.; Quintero-Vargas, J.C.; Ayala-Vasquez, M.; Schermer, T.; Seidl, M.F.; Santos-Paiva, M.; Noguera, A.M.; Aguilera-Galvez, C.; Wittenberg, A.; Hofstede, R.; et al. First report of fusarium wilt tropical race 4 in cavendish bananas caused by Fusarium odoratissimum in Colombia. Plant Dis. 2020, 104, 994. [Google Scholar] [CrossRef]
- Lambert, J. Alarm as devastating banana fungus reaches the Americas. Nature 2019. [Google Scholar] [CrossRef] [PubMed]
- Segura, R.A.; Serrano, E.; Pocasangre, L.; Acuña, O.; Bertsch, F.; Stoorvogel, J.J.; Sandoval, J.A. Chemical and microbiological interactions between soils and roots in commercial banana plantations (Musa AAA, cv. Cavendish). Sci. Hortic. 2015, 197, 66–71. [Google Scholar] [CrossRef]
- Lopez, A.; Solís, P. Contenidos e interacciones de los nutrimentos en tres zonas bananeras de Costa Rica. CORBANA 1991, 15, 15–25. [Google Scholar]
- Arias, F.; Mata, R.; Alvarado, A.; Serrano, E.; Laguna, J. Chemical properties and classifica-tion of soils cultivated with banana in the Caribbean lowlands of Costa Rica. Agron. Costarric. 2010, 32, 177–195. [Google Scholar]
- Arias, F.; Alvarado, A.; Mata, R.; Serrano, E.; Laguna, J. Clay mineralogy and soil fertility of banana plantation alluvial soils at Costa Rica’s Caribbean lowlands. Agron. Costarric. 2010, 32, 223–236. [Google Scholar]
- Klinkert, S. A Mechanistic Approach to Soil Variability at Different Scale Levels: A Case Study for the Atlantic Zone of Costa Rica; Wageningen UR: Wageningen, The Netherlands, 2014. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing: Reference Index; R Foundation for Statistical Computing: Vienna, Austria, 2010; ISBN 978-3-900051-07-5. [Google Scholar]
- Højsgaard, S.; Yan, J. The R Package geepack for Generalized Estimating Equations. J. Stat. Softw. 2006, 15. [Google Scholar] [CrossRef] [Green Version]
- Yan, J. Geepack: Yet another package for generalized estimating equations. R-News 2002, 2/3, 12–14. [Google Scholar]
- Yan, J.; Fine, J. Estimating equations for association structures. Stat. Med. 2004, 23, 859–874. [Google Scholar] [CrossRef]
- Russell, V.; Buerkner, P.; Herve, M.; Love, J.; Rielbl, H.; Singmann, H. emmeans: Estimated marginal means, aka least-squares means. 2021, 34, pp. 216–221. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 15 February 2021).
- Pattison, A.B.; Moody, P.W.; Badcock, K.A.; Smith, L.J.; Armour, J.A.; Rasiah, V.; Cobon, J.A.; Gulino, L.-M.; Mayer, R. Development of key soil health indicators for the Australian banana industry. Appl. Soil Ecol. 2008, 40, 155–164. [Google Scholar] [CrossRef]
- Chaperon, S.; Sauvé, S. Toxicity Interaction of Metals (Ag, Cu, Hg, Zn) to Urease and Dehydrogenase Activities in Soils. Soil Biol. Biochem. 2007, 39, 2329–2338. [Google Scholar] [CrossRef]
- Almeida, N.O.; Teixeira, R.A.; Carneiro, F.A.; de Oliveira, C.M.; Ribeiro, V.A.; Lobo Júnior, M.; Rocha, M.R. da Occurrence and Correlations of Nematodes, Fusarium Oxysporum and Edaphic Factors on Banana Plantations. J. Phytopathol. 2018, 166, 265–272. [Google Scholar] [CrossRef]
- Huber, D.M.; Watson, R. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Geense, P.; Pattison, A.B.; Kukulies, T.L.; Scholberg, J.M.S.; Molina, A.B. Can changes in soil properties in organic banana production suppress Fusarium wilt? Nat. Resour. 2015, 06, 181–195. [Google Scholar] [CrossRef] [Green Version]
- De Rocha, A.J.; dos Ferreira, M.S.; de Rocha, L.S.; Oliveira, S.A.S.; Amorim, E.P.; Mizubuti, E.S.G.; Haddad, F. Interaction between Fusarium oxysporum f. sp. cubense and Radopholus similis can lead to changes in the resistance of banana cultivars to Fusarium wilt. Eur. J. Plant Pathol. 2020, 158, 403–417. [Google Scholar] [CrossRef]
- Guillén, C.; Tixier, P.; Tapia Fernández, A.; Conejo Barboza, A.M.; Sandoval Fernández, J.A.; Lapeyre de Bellaire, L. Can the banana weevil Cosmopolites sordidus be a vector of Fusarium oxysporum f.sp. cubense race 1? Unravelling the internal and external acquisition of effective inoculum. Pest Manag. Sci. 2021, ps.6339. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.A.; Guzmán, J.A.; Gonzálex, M.; Segura, R.A. Impacto de Eventos Climáticos Extremos Sobre la Producción de Banano en Costa Rica. Santa Catarina, Brasil. 2015. Available online: http://banana-networks.org/musalac/files/2015/11/Jorge-Sandoval-Rafael-Segura.pdf (accessed on 15 February 2021).
- Batista, E.; de Brito, E.; Henrique, C.; Arlington, R.; Cavalcanti, I. Influência de Fatores Químicos Do Solo Sobre a Incidência Do Mal-Do-Panamá Na Bananeira Cv. Pacovan Na Paraíba. Rev. Biol. E Ciênc. Terra 2008, 8, 100–109. [Google Scholar]
- Deltour, P.; França, S.C.; Liparini Pereira, O.; Cardoso, I.; De Neve, S.; Debode, J.; Höfte, M. Disease Suppressiveness to Fusarium Wilt of Banana in an Agroforestry System: Influence of Soil Characteristics and Plant Community. Agric. Ecosyst. Environ. 2017, 239, 173–181. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Management | Application | Timing | |
|---|---|---|---|
| General | |||
| Compost | 1 kg per plant in each planting hole | At planting | |
| Startup fertilization | 100 N, 18 K2O, 27 MgO, 53 CaO, 20 S, 5.8 Zn and 0.8 B (in kg ha−1 year−1) | During the first 7 weeks, divided over weekly applications. | |
| Basic nutritional package [36] | 105 P2O5, 610 K2O, 4.0 CaO, 60 S, 9.2 Zn and 3.2 B (in kg ha−1 year−1) | After week 8, divided over 17 3-weekly applications per year | |
| Treatments | |||
| pH | Low: 300 mL of 5% HCl solution and 303 g of CaO 1 per plant per application High: 400 g of Ca(OH)2 per plant per application | 1 week before planting; 8 weeks after planting; between cycle 2 and 3 | |
| N | Low: 232 kg N ha−1 year−1 with NH4NO3 and CaNO3 High: 605 kg ha−1 year−1 with NH4NO3 and CaNO3 | Starting in week 8. Divided over 17 3-weekly applications per year | |
| CaMg 2 | Low: - High: 315 kg CaO ha−1 year−1 with Ca3(BO3)2 and CaNO3. 150 kg MgO ha−1 year−1 with MgSO4 | Starting in week 8. Divided over 17 3-weekly applications per year | |
| Mn | Low: - High: 32 kg Mn ha−1 year−1 with MnSO4 | Starting in week 8. Divided over 17 3-weekly applications per year | |
| Treatment | Units | Soil Analysis | ||
|---|---|---|---|---|
| Start | Cycle 1–2 | Cycle 3–4 | ||
| Exp-west (La Rita) | ||||
| N-low | % | n.d | 0.30 | 0.30 |
| N-high | % | n.d | 0.31 | 0.30 |
| pH-low | - | 5.5 | 5.4 *** | 5.1 *** |
| pH-high | - | 5.5 | 5.7 | 5.6 |
| Ca-low | cmol (+) kg−1 | 4.60 | 4.18 | 4.06 |
| Ca-high | cmol (+) kg−1 | 4.60 | 4.26 | 4.26 |
| Mg-low | cmol (+) kg−1 | 2.90 | 1.41 ** | 1.84 *** |
| Mg-high | cmol (+) kg−1 | 2.90 | 1.67 | 2.35 |
| Mn-low | mg kg−1 | 11 | 15 *** | 16 *** |
| Mn-high | mg kg−1 | 11 | 18 | 23 |
| Exp-east (28 Millas) | ||||
| N-low | % | n.d | 0.16 | 0.15 |
| N-high | % | n.d | 0.17 | 0.14 |
| pH-low | - | 6.3 | 5.4 ** | 5.4 *** |
| pH-high | - | 6.3 | 5.8 | 5.9 |
| Ca-low | cmol (+) kg−1 | 22.2 | 21.9 | 22.6 |
| Ca-high | cmol (+) kg−1 | 22.2 | 21.8 | 22.8 |
| Mg-low | cmol (+) kg−1 | 10.9 | 9.65 | 9.83 |
| Mg-high | cmol (+) kg−1 | 10.9 | 9.54 | 10.2 |
| Mn-low | mg kg−1 | 88 | 90 *** | 88 |
| Mn-high | mg kg−1 | 88 | 133 | 87 |
| Treatment | N | P | K | Ca | Mg | S | Fe | Cu | Zn | Mn | B |
|---|---|---|---|---|---|---|---|---|---|---|---|
| % dry matter–1 | mg Kg–1 | ||||||||||
| Exp-west (La Rita), 1st cycle | |||||||||||
| pH-low | 2.93 | 0.21 | 4.32 | 0.60 | 0.28 | 0.19 | 67 | 9 | 17 | 172 * | 8 * |
| pH-high | 2.90 | 0.21 | 4.39 | 0.57 | 0.27 | 0.19 | 64 | 9 | 17 | 147 | 9 |
| N-low | 2.86 | 0.21 | 4.43 | 0.56 * | 0.27 | 0.19 * | 67 | 9 | 17 | 149 * | 9 |
| N-high | 2.96 | 0.21 | 4.28 | 0.61 | 0.29 | 0.20 | 64 | 9 | 16 | 169 | 8 |
| CaMg-low | 2.92 | 0.21 | 4.34 | 0.58 | 0.27 * | 0.19 | 67 | 9 | 17 | 165 | 8 |
| CaMg-high | 2.91 | 0.21 | 4.36 | 0.59 | 0.29 | 0.19 | 64 | 9 | 17 | 154 | 9 |
| Mn-low | 2.89 | 0.21 | 4.40 | 0.57 | 0.28 | 0.19 | 65 | 9 | 17 | 149 * | 9 |
| Mn-high | 2.94 | 0.21 | 4.30 | 0.60 | 0.28 | 0.20 | 66 | 9 | 17 | 170 | 8 |
| Exp-west (La Rita), 2nd cycle | |||||||||||
| pH-low | 2.55 | 0.19 | 3.77 | 0.67 | 0.28 | 0.18 | 58 | 7 | 17 | 183 * | 9 * |
| pH-high | 2.54 | 0.19 | 3.82 | 0.75 | 0.26 | 0.18 | 57 | 7 | 17 | 163 | 11 |
| N-low | 2.50 * | 0.20 * | 3.86 * | 0.66 * | 0.27 | 0.19 * | 59 * | 7 | 18 * | 155 * | 10 * |
| N-high | 2.59 | 0.18 | 3.73 | 0.76 | 0.27 | 0.18 | 56 | 7 | 17 | 190 | 9 |
| CaMg-low | 2.53 | 0.19 | 3.80 | 0.71 | 0.26 * | 0.18 | 58 | 7 | 17 | 176 | 10 |
| CaMg-high | 2.56 | 0.19 | 3.79 | 0.71 | 0.28 | 0.18 | 57 | 7 | 17 | 170 | 10 |
| Mn-low | 2.53 | 0.19 | 3.83 | 0.71 | 0.27 | 0.18 | 58 | 7 | 17 | 157 * | 10 |
| Mn-high | 2.56 | 0.19 | 3.76 | 0.72 | 0.27 | 0.18 | 58 | 7 | 17 | 189 | 10 |
| Exp-east (28 Millas), 1st cycle | |||||||||||
| pH-low | 2.42 | 0.19 | 3.70 * | 0.66 | 0.28 | 0.17 | 58 | 7 | 17 * | 408 | 12 |
| pH-high | 2.44 | 0.19 | 3.58 | 0.63 | 0.27 | 0.17 | 56 | 7 | 16 | 351 | 12 |
| N-low | 2.34 * | 0.20 * | 3.73 * | 0.60 * | 0.26 * | 0.17 | 57 | 7 | 17 | 355 | 12 |
| N-high | 2.52 | 0.19 | 3.55 | 0.69 | 0.28 | 0.17 | 58 | 7 | 17 | 403 | 11 |
| CaMg-low | 2.44 | 0.19 | 3.74 * | 0.65 | 0.27 | 0.17 | 58 | 7 | 17 | 378 | 12 |
| CaMg-high | 2.43 | 0.19 | 3.54 | 0.64 | 0.28 | 0.17 | 57 | 7 | 17 | 382 | 12 |
| Mn-low | 2.42 | 0.19 | 3.71 * | 0.64 | 0.27 | 0.17 | 57 | 7 | 17 | 366 * | 11 |
| Mn-high | 2.45 | 0.19 | 3.57 | 0.65 | 0.28 | 0.17 | 58 | 7 | 17 | 394 | 12 |
| Exp-east (28 Millas), 2nd cycle | |||||||||||
| pH-low | 2.50 | 0.19 | 3.27 | 0.81 * | 0.36 | 0.18 | 66 | 8 | 17 | 298 | 10 |
| pH-high | 2.49 | 0.19 | 3.27 | 0.89 | 0.36 | 0.18 | 67 | 7 | 17 | 272 | 11 |
| N-low | 2.43 * | 0.20 * | 3.39 * | 0.78 * | 0.34 * | 0.18 | 67 | 7 | 17 | 289 | 11 |
| N-high | 2.55 | 0.18 | 3.16 | 0.91 | 0.38 | 0.18 | 67 | 8 | 17 | 282 | 10 |
| CaMg-low | 2.50 | 0.19 | 3.34 | 0.85 | 0.36 | 0.18 | 67 | 8 | 17 | 294 | 11 |
| CaMg-high | 2.49 | 0.19 | 3.21 | 0.85 | 0.37 | 0.18 | 67 | 8 | 17 | 276 | 10 |
| Mn-low | 2.51 | 0.20 * | 3.29 | 0.83 * | 0.36 | 0.18 | 67 | 8 | 17 | 287 | 11 |
| Mn-high | 2.48 | 0.18 | 3.26 | 0.87 | 0.36 | 0.18 | 66 | 7 | 17 | 283 | 10 |
| Variable | Exp-West La Rita | Exp-East 28 Millas | ||
|---|---|---|---|---|
| 1st Cycle | 2nd Cycle | 1st Cycle | 2nd Cycle | |
| Plant height (cm) | 337 | 369 | 300 | 355 |
| Pseudostem circumference (cm) | 75 | 83 | 64 | 79 |
| Bunch weight (kg) | 25.2 | 29 | 19.3 | 24.8 |
| Clusters bunch–1 | 9.2 | 10.3 | 8.5 | 8.9 |
| Harvested fruit § (ton ha–1) | 39.9 | 41.3 | 30.6 | 31.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segura M., R.A.; Stoorvogel, J.J.; Blanco R., F.A.; Sandoval F., J.A. A Medium-Term Field Experiment to Study the Effect of Managing Soil Chemical Properties on Fusarium Wilt in Banana (Musa AAA). J. Fungi 2021, 7, 261. https://doi.org/10.3390/jof7040261
Segura M. RA, Stoorvogel JJ, Blanco R. FA, Sandoval F. JA. A Medium-Term Field Experiment to Study the Effect of Managing Soil Chemical Properties on Fusarium Wilt in Banana (Musa AAA). Journal of Fungi. 2021; 7(4):261. https://doi.org/10.3390/jof7040261
Chicago/Turabian StyleSegura M., Rafael A., Jetse J. Stoorvogel, Fabio A. Blanco R., and Jorge A. Sandoval F. 2021. "A Medium-Term Field Experiment to Study the Effect of Managing Soil Chemical Properties on Fusarium Wilt in Banana (Musa AAA)" Journal of Fungi 7, no. 4: 261. https://doi.org/10.3390/jof7040261
APA StyleSegura M., R. A., Stoorvogel, J. J., Blanco R., F. A., & Sandoval F., J. A. (2021). A Medium-Term Field Experiment to Study the Effect of Managing Soil Chemical Properties on Fusarium Wilt in Banana (Musa AAA). Journal of Fungi, 7(4), 261. https://doi.org/10.3390/jof7040261