Author Contributions
Conceptualization, D.A.S.; Formal analysis, H.N., G.S.; Validation, H.N., G.S.; Writing-review H.N., G.S., P.W., E.D., D.A.S.; Writing editing, P.W., D.A.S.; Investigation, H.N., G.S., D.A.S.; Methodology, H.N., G.S., P.W., D.A.S.; Visualization, H.N., G.S., D.A.S.; Writing original draft, D.A.S., G.S.; Funding acquisition, D.A.S.; Project administration, D.A.S.; Resources, P.W., E.D., D.A.S.; Software, H.N., G.S.; Supervision, D.A.S.; Data curation H.N., G.S., D.A.S. All authors have read and agreed to the published version of the manuscript.
Figure 1.
Molecules in this study. In all, R1 is an H atom. In Pseudomonas quinolone signal (PQS), 2-heptyl-4-hydroxyquinoline (HHQ), 2-heptyl-3-chloro-4(1H)-quinolone (3Cl-PQS) and 2-heptyl-3-amino-4(1H)-quinolone (3NH2-PQS) R2 is an alkyl side chain, C7H15. In C1-PQS, C3-PQS, C5-PQS, and C9-PQS R2 is an alkyl side chain of 1, 3, 5, and 9 carbon molecules, respectively. In PQS and HHQ, R3 is a hydroxyl group or an H atom, respectively. In 3Cl-PQS and 3NH2-PQS, R3 is a chlorine or primary amine group, respectively.
Figure 1.
Molecules in this study. In all, R1 is an H atom. In Pseudomonas quinolone signal (PQS), 2-heptyl-4-hydroxyquinoline (HHQ), 2-heptyl-3-chloro-4(1H)-quinolone (3Cl-PQS) and 2-heptyl-3-amino-4(1H)-quinolone (3NH2-PQS) R2 is an alkyl side chain, C7H15. In C1-PQS, C3-PQS, C5-PQS, and C9-PQS R2 is an alkyl side chain of 1, 3, 5, and 9 carbon molecules, respectively. In PQS and HHQ, R3 is a hydroxyl group or an H atom, respectively. In 3Cl-PQS and 3NH2-PQS, R3 is a chlorine or primary amine group, respectively.
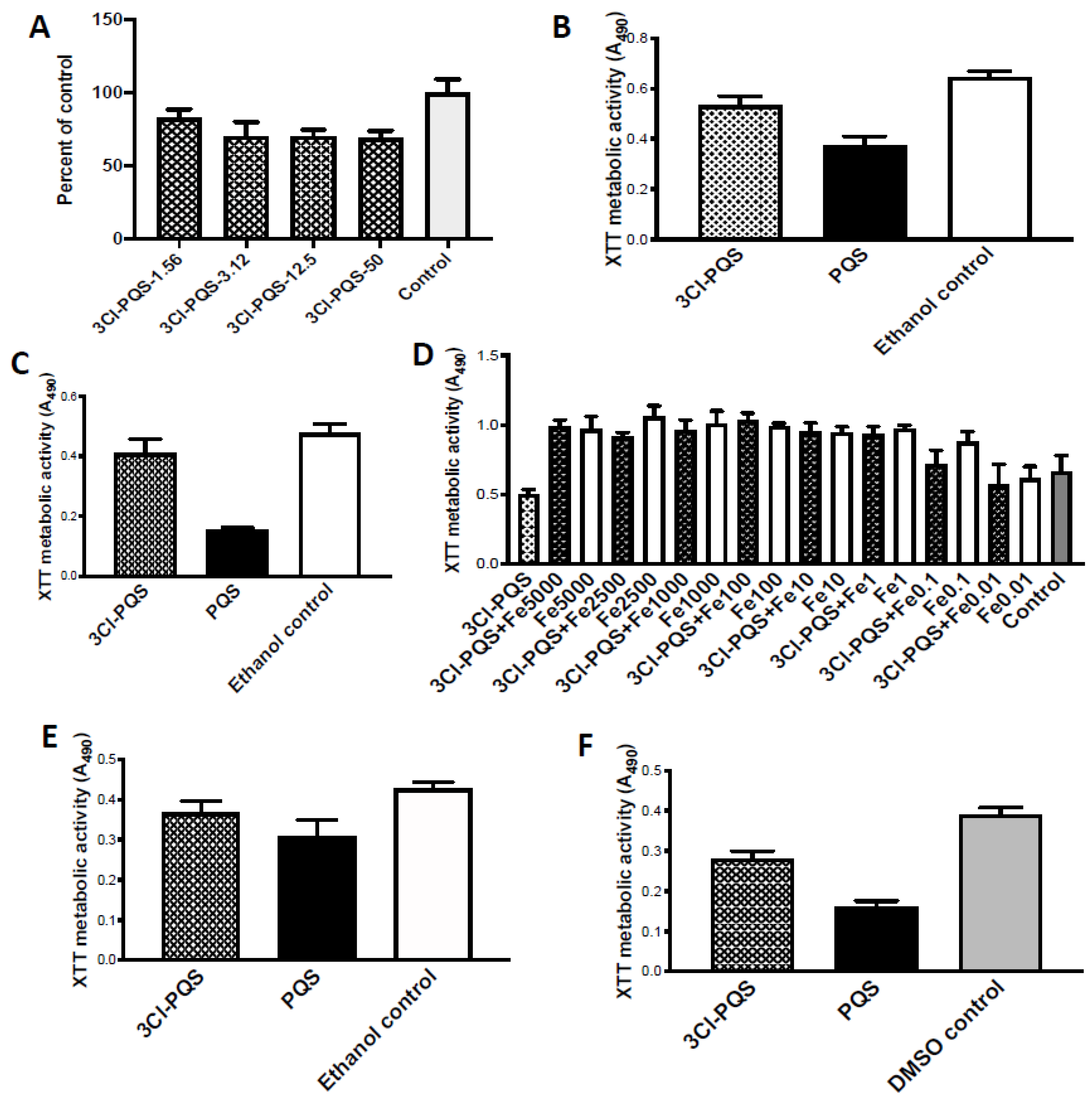
Figure 2.
Studies with 3Cl-PQS on preformed Af biofilm metabolism, assessed by XTT assay. (
A) Dose response study, strain 10AF, control (same amount of DMSO as in test articles). Designations 1.56, etc. are μg/mL. Dose-response is shallow; 1.56 μg/mL,
p < 0.01 compared to control, and other concentrations
p < 0.001 compared to control. (
B,
C), comparison to PQS, all at 1.56 μg/mL, and to ethanol control (shown) identical to DMSO control. 3Cl-PQS is less inhibitory than PQS. In (
B), using 10AF, 3Cl-PQS is
p < 0.01, and PQS
p < 0.001, compared to control; PQS is
p < 0.001 compared to 3Cl-PQS. In (
C), studying strain Af293, 3Cl-PQS is
p > 0.05, and PQS
p < 0.001, compared to control; PQS is
p < 0.001 compared to 3Cl-PQS. (
D) Effect of Fe on 3Cl-PQS (12.5 μg/mL) inhibition of strain 10AF. Control (same amount of DMSO as in test article), numbers 0.01–5000 are Fe μM concentration. 3Cl-PQS inhibits Af metabolism (left bar,
p < 0.01). Fe alone boosts Af metabolism (white bars), starting at Fe 0.1 μM, all
p < 0.01–0.001 compared to control. Fe reverses 3Cl-PQS inhibition in combinations (black bars), beginning at Fe 0.01 μg/mL, where Fe 0.01 μM + 3Cl-PQS is now
p > 0.05 compared to control or to 3Cl-PQS alone, and all other 3Cl-PQS + Fe combinations are greater (
p < 0.001) compared to control or to 3Cl-PQS alone. 3Cl-PQS + Fe combinations do not boost Af metabolism above same concentration of Fe alone, unlike previously reported with PQS [
18] (and at two concentrations, Fe 0.1 or Fe 2500 μM + 3Cl-PQS, are significantly,
p < 0.05, less than corresponding Fe alone). (
E,
F), effects in hypoxic environment. DMSO and ethanol control results shown identical. (
E) is Af isolate 10AF, (
F) is Af isolate Af293, both tested at 1.56 μg/mL of test articles. In (
E), 3Cl-PQS is
p < 0.05 compared to control or to PQS, and PQS
p < 0.001 compared to controls. In (
F), PQS and 3Cl-PQS both
p < 0.001 compared to controls, and PQS and 3Cl-PQS differ by
p < 0.001.
Figure 2.
Studies with 3Cl-PQS on preformed Af biofilm metabolism, assessed by XTT assay. (
A) Dose response study, strain 10AF, control (same amount of DMSO as in test articles). Designations 1.56, etc. are μg/mL. Dose-response is shallow; 1.56 μg/mL,
p < 0.01 compared to control, and other concentrations
p < 0.001 compared to control. (
B,
C), comparison to PQS, all at 1.56 μg/mL, and to ethanol control (shown) identical to DMSO control. 3Cl-PQS is less inhibitory than PQS. In (
B), using 10AF, 3Cl-PQS is
p < 0.01, and PQS
p < 0.001, compared to control; PQS is
p < 0.001 compared to 3Cl-PQS. In (
C), studying strain Af293, 3Cl-PQS is
p > 0.05, and PQS
p < 0.001, compared to control; PQS is
p < 0.001 compared to 3Cl-PQS. (
D) Effect of Fe on 3Cl-PQS (12.5 μg/mL) inhibition of strain 10AF. Control (same amount of DMSO as in test article), numbers 0.01–5000 are Fe μM concentration. 3Cl-PQS inhibits Af metabolism (left bar,
p < 0.01). Fe alone boosts Af metabolism (white bars), starting at Fe 0.1 μM, all
p < 0.01–0.001 compared to control. Fe reverses 3Cl-PQS inhibition in combinations (black bars), beginning at Fe 0.01 μg/mL, where Fe 0.01 μM + 3Cl-PQS is now
p > 0.05 compared to control or to 3Cl-PQS alone, and all other 3Cl-PQS + Fe combinations are greater (
p < 0.001) compared to control or to 3Cl-PQS alone. 3Cl-PQS + Fe combinations do not boost Af metabolism above same concentration of Fe alone, unlike previously reported with PQS [
18] (and at two concentrations, Fe 0.1 or Fe 2500 μM + 3Cl-PQS, are significantly,
p < 0.05, less than corresponding Fe alone). (
E,
F), effects in hypoxic environment. DMSO and ethanol control results shown identical. (
E) is Af isolate 10AF, (
F) is Af isolate Af293, both tested at 1.56 μg/mL of test articles. In (
E), 3Cl-PQS is
p < 0.05 compared to control or to PQS, and PQS
p < 0.001 compared to controls. In (
F), PQS and 3Cl-PQS both
p < 0.001 compared to controls, and PQS and 3Cl-PQS differ by
p < 0.001.
![Jof 07 00343 g002]()
Figure 3.
Studies with 3NH
2-PQS. (
A) Dose-response study with preformed 10AF biofilm. Minimal dose-response. “RPMI control” was biofilm without test article or diluent, “RPMI + Ethanol control” contained same concentration of ethanol diluent as present in test article wells; no difference between these controls. Both controls were also included in sections (
B) through (
D) of
Figure 3, but as the results of these two controls was always the same, for simplification, only one control is shown in sections (
C) through (
F). Twofold dilutions of 3NH
2-PQS were made from 50 to 0.78 μg/mL. Only at the very highest doses (12.5–50 μg/mL, shown) was there a suggestion of dose response, but there was no statistically significant difference among those bars. 3NH
2-PQS at 0.78 μg/mL was not different from controls shown, and bars showing results with 1.56–6.25 μg/mL were not different from that of 12.5 μg/mL, so are not shown in order to simplify the figure. All dilution results from 1.56–50 μg/mL were
p < 0.001 compared to controls. (
B). Dose-response study with 10AF biofilm formation. Same experimental design as in A, but test against Af biofilm formation. Dilutions 0.78–6.25 μg/mL resulted in slight inhibition, but
p > 0.05 compared to controls, and no dose-response. 3NH
2-PQS 12.5 and 25 μg/mL
p < 0.01 compared to controls, and 50 μg/mL
p < 0.001 compared to controls. (
C,
D) Comparison with PQS, preformed biofilm, test reagents at 1.56 μg/mL. C, isolate 10AF. 3NH
2-PQS is less potent inhibitor than PQS (
p < 0.01). Both are
p < 0.01 compared to controls. D, isolate Af293. PQS more inhibitory (
p < 0.001) compared to 3NH2-PQS. Both are
p < 0.001 compared to controls. (
E) 3NH
2-PQS (12.5 μg/mL) (left bar) inhibits preformed 10AF biofilm,
p < 0.001 compared to control (right bar). Fe (0.1–1000 μM; numbers on x axis are these μM concentrations) reverses 3NH
2-PQS inhibition (3NH
2-PQS + Fe concentrations
p < 0.001 compared to 3NH
2-PQS). Fe boosts Af metabolism, but 3NH
2-PQS + Fe does not increase metabolism above Fe alone. (
F) Action of 3NH
2-PQS under hypoxic conditions. 3NH
2-PQS inhibitory to preformed Af293 biofilm (
p < 0.05 compared to controls). PQS more inhibitory (
p < 0.001) than controls, and more inhibitory compared to 3NH
2-PQS (
p < 0.001). Both PQS and 3NH
2-PQS are at 1.56 μg/mL.
Figure 3.
Studies with 3NH
2-PQS. (
A) Dose-response study with preformed 10AF biofilm. Minimal dose-response. “RPMI control” was biofilm without test article or diluent, “RPMI + Ethanol control” contained same concentration of ethanol diluent as present in test article wells; no difference between these controls. Both controls were also included in sections (
B) through (
D) of
Figure 3, but as the results of these two controls was always the same, for simplification, only one control is shown in sections (
C) through (
F). Twofold dilutions of 3NH
2-PQS were made from 50 to 0.78 μg/mL. Only at the very highest doses (12.5–50 μg/mL, shown) was there a suggestion of dose response, but there was no statistically significant difference among those bars. 3NH
2-PQS at 0.78 μg/mL was not different from controls shown, and bars showing results with 1.56–6.25 μg/mL were not different from that of 12.5 μg/mL, so are not shown in order to simplify the figure. All dilution results from 1.56–50 μg/mL were
p < 0.001 compared to controls. (
B). Dose-response study with 10AF biofilm formation. Same experimental design as in A, but test against Af biofilm formation. Dilutions 0.78–6.25 μg/mL resulted in slight inhibition, but
p > 0.05 compared to controls, and no dose-response. 3NH
2-PQS 12.5 and 25 μg/mL
p < 0.01 compared to controls, and 50 μg/mL
p < 0.001 compared to controls. (
C,
D) Comparison with PQS, preformed biofilm, test reagents at 1.56 μg/mL. C, isolate 10AF. 3NH
2-PQS is less potent inhibitor than PQS (
p < 0.01). Both are
p < 0.01 compared to controls. D, isolate Af293. PQS more inhibitory (
p < 0.001) compared to 3NH2-PQS. Both are
p < 0.001 compared to controls. (
E) 3NH
2-PQS (12.5 μg/mL) (left bar) inhibits preformed 10AF biofilm,
p < 0.001 compared to control (right bar). Fe (0.1–1000 μM; numbers on x axis are these μM concentrations) reverses 3NH
2-PQS inhibition (3NH
2-PQS + Fe concentrations
p < 0.001 compared to 3NH
2-PQS). Fe boosts Af metabolism, but 3NH
2-PQS + Fe does not increase metabolism above Fe alone. (
F) Action of 3NH
2-PQS under hypoxic conditions. 3NH
2-PQS inhibitory to preformed Af293 biofilm (
p < 0.05 compared to controls). PQS more inhibitory (
p < 0.001) than controls, and more inhibitory compared to 3NH
2-PQS (
p < 0.001). Both PQS and 3NH
2-PQS are at 1.56 μg/mL.
![Jof 07 00343 g003]()
Figure 4.
Studies of alkyl side chains on PQS. Dose-response assays. (A–D) are C1-PQS, C3-PQS, C5-PQS, C9-PQS respectively, tested against preformed 10AF biofilm, DMSO control shown. (A) Twofold dilutions C1-PQS 50 to 0.2 μg/mL were made. Dilutions 12.5 to 0.2 μg/mL produced slight and insignificant (p > 0.05) inhibition compared to control, and not different from each other (not shown). Only at 25 and 50 μg/mL was inhibition significant, and only at p < 0.05. (B) Same plan as in A, C3-PQS; 0.2 μg/mL produced slight and insignificant (p > 0.05) inhibition, and 0.39 to 50 μg/mL were all inhibitory compared to control (p < 0.01) and not significantly different from each other in effect. (C) Same plan as in A, C5-PQS. All dilutions were inhibitory (p < 0.05) compared to control, but were not different from each other. (D) Same plan as in A, C9-PQS. Slight dose-response, as shown. All dilutions were inhibitory compared to controls, but were not significantly different from each other; 0.2 to 0.78 μg/mL were p < 0.05 compared to controls, and 1.56 to 50 μg/mL were p < 0.001 compared to controls.
Figure 4.
Studies of alkyl side chains on PQS. Dose-response assays. (A–D) are C1-PQS, C3-PQS, C5-PQS, C9-PQS respectively, tested against preformed 10AF biofilm, DMSO control shown. (A) Twofold dilutions C1-PQS 50 to 0.2 μg/mL were made. Dilutions 12.5 to 0.2 μg/mL produced slight and insignificant (p > 0.05) inhibition compared to control, and not different from each other (not shown). Only at 25 and 50 μg/mL was inhibition significant, and only at p < 0.05. (B) Same plan as in A, C3-PQS; 0.2 μg/mL produced slight and insignificant (p > 0.05) inhibition, and 0.39 to 50 μg/mL were all inhibitory compared to control (p < 0.01) and not significantly different from each other in effect. (C) Same plan as in A, C5-PQS. All dilutions were inhibitory (p < 0.05) compared to control, but were not different from each other. (D) Same plan as in A, C9-PQS. Slight dose-response, as shown. All dilutions were inhibitory compared to controls, but were not significantly different from each other; 0.2 to 0.78 μg/mL were p < 0.05 compared to controls, and 1.56 to 50 μg/mL were p < 0.001 compared to controls.
![Jof 07 00343 g004]()
Figure 5.
Studies of alkyl side chains on PQS. Comparisons of congeners with PQS. (A) Target 10AF, all molecules studied at 12.5 μg/mL. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05, p < 0.01, p < 0.05, p < 0.05, p < 0.001 compared to control. DMSO control results same as ethanol control shown. C1-PQS, C3-PQS, C5-PQS, C9-PQS are all p < 0.001 compared to PQS. (B) Target 10AF, all molecules studied at 1.56 μg/mL; separate plates, so data is referent to controls on each plate. DMSO controls shown, were same as ethanol and RPMI controls. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05 (actual boost above control), p < 0.01, p < 0.05, p < 0.05, p < 0.001 compared to control. C1-PQS, C3-PQS, C5-PQS, C9-PQS are p < 0.001, p < 0.001, p < 0.01, p < 0.01, respectively, compared to inhibition by PQS. (C) Target Af293, all molecules studied at 1.56 μg/mL. Shown are DMSO control same as ethanol control. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05 (actual boost above control), p > 0.05, p < 0.05, p < 0.001, p < 0.001 respectively, compared to control. C1-PQS, C3-PQS, C5-PQS, C9-PQS are all less inhibitory than PQS (p < 0.001).
Figure 5.
Studies of alkyl side chains on PQS. Comparisons of congeners with PQS. (A) Target 10AF, all molecules studied at 12.5 μg/mL. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05, p < 0.01, p < 0.05, p < 0.05, p < 0.001 compared to control. DMSO control results same as ethanol control shown. C1-PQS, C3-PQS, C5-PQS, C9-PQS are all p < 0.001 compared to PQS. (B) Target 10AF, all molecules studied at 1.56 μg/mL; separate plates, so data is referent to controls on each plate. DMSO controls shown, were same as ethanol and RPMI controls. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05 (actual boost above control), p < 0.01, p < 0.05, p < 0.05, p < 0.001 compared to control. C1-PQS, C3-PQS, C5-PQS, C9-PQS are p < 0.001, p < 0.001, p < 0.01, p < 0.01, respectively, compared to inhibition by PQS. (C) Target Af293, all molecules studied at 1.56 μg/mL. Shown are DMSO control same as ethanol control. C1-PQS, C3-PQS, C5-PQS, C9-PQS, PQS respectively, are p > 0.05 (actual boost above control), p > 0.05, p < 0.05, p < 0.001, p < 0.001 respectively, compared to control. C1-PQS, C3-PQS, C5-PQS, C9-PQS are all less inhibitory than PQS (p < 0.001).
![Jof 07 00343 g005]()
Figure 6.
Studies of alkyl side chains on PQS. Fe reverses inhibition by side chain congeners, but combination of congeners with Fe does not boost metabolism above Fe alone. Target 10AF, all congeners studied at 12.5 μg/mL, DMSO controls are shown. Dilutions of Fe were made from 0.01 to 1000 μM final concentrations (these are the numbers on the x axis). In some experiments there were visible solubility problems with Fe 100–1000 μM, and the study values were the same as the next lower concentration; those bars are omitted. (A) C3-PQS (left bar) inhibits 10AF (p < 0.01) compared to control (right bar). Fe begins to reverse C3-PQS effect at 0.01 μM Fe, and with concentrations of Fe 100 μM with C3-PQS, are all at p < 0.001 compared to C3-PQS alone. No significant boosting across these concentrations above Fe alone, for C3-PQS + Fe. (B) C5-PQS (left bar) inhibits 10AF (p < 0.001) compared to controls (right bar). Fe evidently begins to reverse C5-PQS effect at 0.1 μM Fe, and at concentrations of Fe 1 to 1000 μM with C5-PQS, loss of inhibition is significant (p < 0.001) compared to C5-PQS alone. No significant boosting across these concentrations above Fe alone, for C5-PQS + Fe. (C) C9-PQS (left bar) inhibits 10AF (p < 0.01) compared to controls (right bar). Fe reverses C9-PQS effect at 0.1 to 10 μM Fe, loss of inhibition is significant (p < 0.001) compared to C9-PQS alone. No significant boosting across these concentrations above Fe alone, for C9-PQS + Fe.
Figure 6.
Studies of alkyl side chains on PQS. Fe reverses inhibition by side chain congeners, but combination of congeners with Fe does not boost metabolism above Fe alone. Target 10AF, all congeners studied at 12.5 μg/mL, DMSO controls are shown. Dilutions of Fe were made from 0.01 to 1000 μM final concentrations (these are the numbers on the x axis). In some experiments there were visible solubility problems with Fe 100–1000 μM, and the study values were the same as the next lower concentration; those bars are omitted. (A) C3-PQS (left bar) inhibits 10AF (p < 0.01) compared to control (right bar). Fe begins to reverse C3-PQS effect at 0.01 μM Fe, and with concentrations of Fe 100 μM with C3-PQS, are all at p < 0.001 compared to C3-PQS alone. No significant boosting across these concentrations above Fe alone, for C3-PQS + Fe. (B) C5-PQS (left bar) inhibits 10AF (p < 0.001) compared to controls (right bar). Fe evidently begins to reverse C5-PQS effect at 0.1 μM Fe, and at concentrations of Fe 1 to 1000 μM with C5-PQS, loss of inhibition is significant (p < 0.001) compared to C5-PQS alone. No significant boosting across these concentrations above Fe alone, for C5-PQS + Fe. (C) C9-PQS (left bar) inhibits 10AF (p < 0.01) compared to controls (right bar). Fe reverses C9-PQS effect at 0.1 to 10 μM Fe, loss of inhibition is significant (p < 0.001) compared to C9-PQS alone. No significant boosting across these concentrations above Fe alone, for C9-PQS + Fe.
![Jof 07 00343 g006]()
Figure 7.
Studies of alkyl side chains on PQS. Effects in hypoxic environment. All molecules tested at 1.56 μg/mL. DMSO control results same as ethanol control. (A) Target 10AF. C1-PQS appears to boost slightly, compared to controls. C9-PQS and PQS inhibit significantly (both p < 0.001) compared to controls. PQS more inhibitory compared to C1-PQS, C3-PQS, C5-PQS, C9-PQS, respectively p < 0.001, p < 0.001, p< 0.05, p > 0.05. (B) Target Af293. C3-PQS, C5-PQS, C9-PQS, PQS all inhibit (p < 0.001) compared to controls, C1-PQS is p > 0.05 compared to controls. PQS more inhibitory compared to C1-PQS, C3-PQS, C5-PQS (p < 0.001 for all), and C9-PQS (p < 0.01).
Figure 7.
Studies of alkyl side chains on PQS. Effects in hypoxic environment. All molecules tested at 1.56 μg/mL. DMSO control results same as ethanol control. (A) Target 10AF. C1-PQS appears to boost slightly, compared to controls. C9-PQS and PQS inhibit significantly (both p < 0.001) compared to controls. PQS more inhibitory compared to C1-PQS, C3-PQS, C5-PQS, C9-PQS, respectively p < 0.001, p < 0.001, p< 0.05, p > 0.05. (B) Target Af293. C3-PQS, C5-PQS, C9-PQS, PQS all inhibit (p < 0.001) compared to controls, C1-PQS is p > 0.05 compared to controls. PQS more inhibitory compared to C1-PQS, C3-PQS, C5-PQS (p < 0.001 for all), and C9-PQS (p < 0.01).
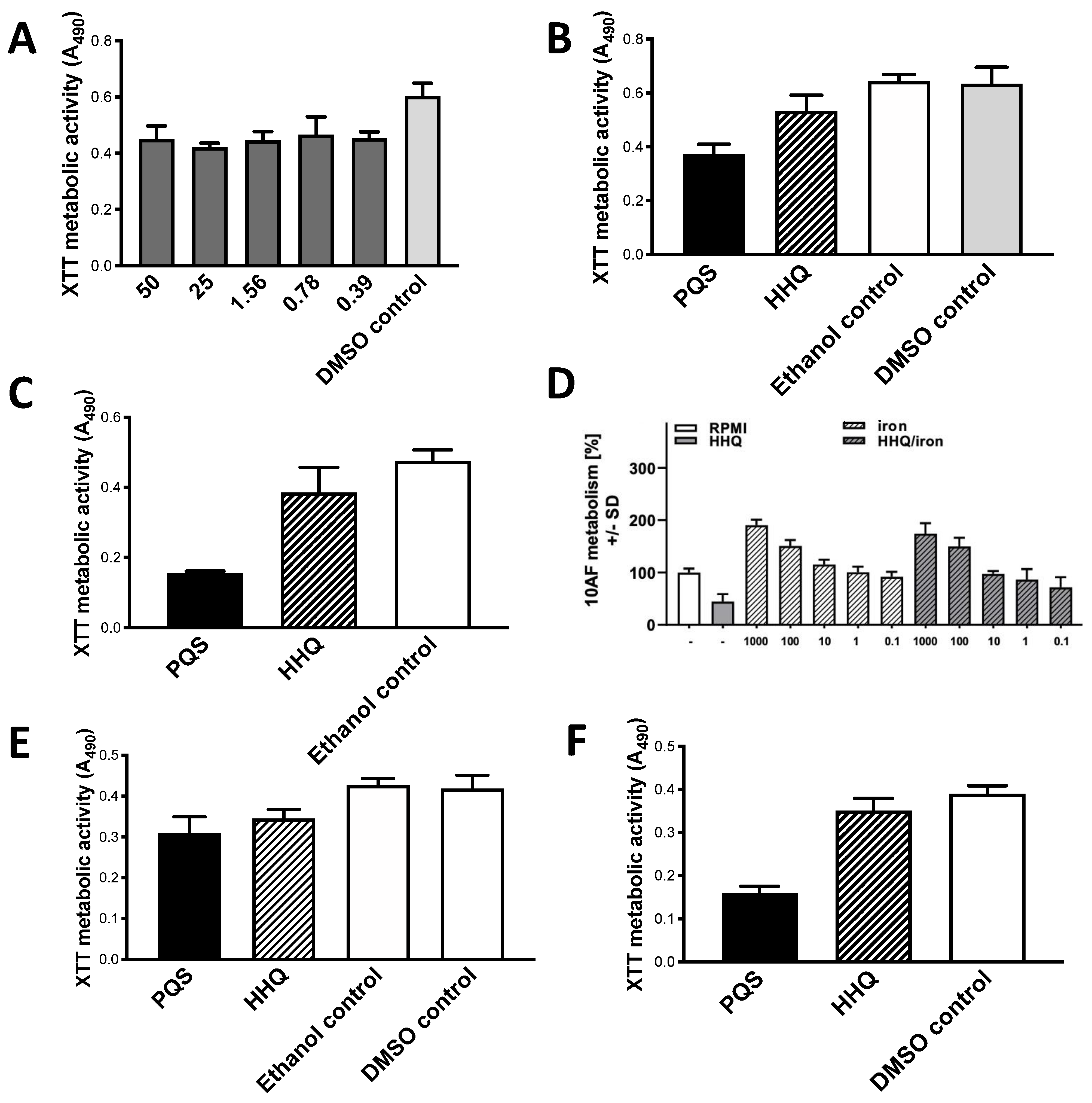
Figure 8.
HHQ inhibits Af preformed biofilm; comparison to PQS. (
A) Dose-response study with preformed 10AF biofilm. Flat dose-response shown. “DMSO control” is same concentration of DMSO diluent as present in test article wells. Twofold dilutions of HHQ were made from 50 to 0.39 μg/mL. Bars showing results with 3.12–12.5 μg/mL were not different from the 1.56 or 25 μg/mL bars, so are not shown in order to simplify the figure. All concentrations shown were
p < 0.001 compared to control except 0.78 μg/mL, which was
p < 0.01. (
B) through (
F): molecules tested at 1.56 μg/mL. DMSO control results same as ethanol control or RPMI alone control, in every panel. (
B,
C), comparison with PQS. (
B) HHQ and PQS inhibit strain 10AF compared to respective controls,
p < 0.01 and
p < 0.001, respectively. PQS more potent than HHQ,
p < 0.001. (
C) HHQ inhibits strain Af293 by 15%, but not significantly. PQS different from HHQ and control,
p < 0.001 both comparisons. (
D) Reversal of inhibition by Fe, no boosting above Fe alone at high Fe concentrations, Af strain 10AF. Fe was diluted twofold from 1000 to 0.1 μM final concentrations (these are the numbers on the x axis), in presence or absence of HHQ. Data presented as % of controls. HHQ alone was inhibitory compared to control,
p < 0.001 (left two bars). At Fe 0.1 μM + HHQ (right bar), inhibition by HHQ is partially reversed; the combination still shows inhibition compared to control (
p < 0.05), but less inhibition (
p < 0.05) compared to HHQ alone. Fe 1–10 μM completely reverses HHQ inhibition in combination (2nd and 3rd bars from right), these are not significantly different from control, and Fe 10 μM + HHQ is
p < 0.05 compared to HHQ alone. Fe alone at 10 μM (5th bar from left) begins to significantly boost Af metabolism (
p < 0.05 compared to control) and higher concentrations are
p < 0.001 compared to control. HHQ inhibition in combinations blunts the Fe boosting about 10-fold, since it is not until the combination of HHQ with Fe 100 μM (4th bar from the right) that there is significant boosting above control (
p < 0.01; Fe 1000 μM is
p < 0.001 compared to control). No combination of HHQ with high concentrations of Fe is boosted above the same concentration of Fe alone. Studies in hypoxic conditions,
Figure 8E,
F. In 8
E, strain 10AF, HHQ and PQS inhibit to 82% and 73% of respective controls, both
p < 0.001; not significantly different. In 8
F, strain Af293, HHQ and PQS inhibit to 87% and 40% of respective controls,
p < 0.05 and
p < 0.001, respectively. HHQ and PQS effect differ by
p < 0.001.
Figure 8.
HHQ inhibits Af preformed biofilm; comparison to PQS. (
A) Dose-response study with preformed 10AF biofilm. Flat dose-response shown. “DMSO control” is same concentration of DMSO diluent as present in test article wells. Twofold dilutions of HHQ were made from 50 to 0.39 μg/mL. Bars showing results with 3.12–12.5 μg/mL were not different from the 1.56 or 25 μg/mL bars, so are not shown in order to simplify the figure. All concentrations shown were
p < 0.001 compared to control except 0.78 μg/mL, which was
p < 0.01. (
B) through (
F): molecules tested at 1.56 μg/mL. DMSO control results same as ethanol control or RPMI alone control, in every panel. (
B,
C), comparison with PQS. (
B) HHQ and PQS inhibit strain 10AF compared to respective controls,
p < 0.01 and
p < 0.001, respectively. PQS more potent than HHQ,
p < 0.001. (
C) HHQ inhibits strain Af293 by 15%, but not significantly. PQS different from HHQ and control,
p < 0.001 both comparisons. (
D) Reversal of inhibition by Fe, no boosting above Fe alone at high Fe concentrations, Af strain 10AF. Fe was diluted twofold from 1000 to 0.1 μM final concentrations (these are the numbers on the x axis), in presence or absence of HHQ. Data presented as % of controls. HHQ alone was inhibitory compared to control,
p < 0.001 (left two bars). At Fe 0.1 μM + HHQ (right bar), inhibition by HHQ is partially reversed; the combination still shows inhibition compared to control (
p < 0.05), but less inhibition (
p < 0.05) compared to HHQ alone. Fe 1–10 μM completely reverses HHQ inhibition in combination (2nd and 3rd bars from right), these are not significantly different from control, and Fe 10 μM + HHQ is
p < 0.05 compared to HHQ alone. Fe alone at 10 μM (5th bar from left) begins to significantly boost Af metabolism (
p < 0.05 compared to control) and higher concentrations are
p < 0.001 compared to control. HHQ inhibition in combinations blunts the Fe boosting about 10-fold, since it is not until the combination of HHQ with Fe 100 μM (4th bar from the right) that there is significant boosting above control (
p < 0.01; Fe 1000 μM is
p < 0.001 compared to control). No combination of HHQ with high concentrations of Fe is boosted above the same concentration of Fe alone. Studies in hypoxic conditions,
Figure 8E,
F. In 8
E, strain 10AF, HHQ and PQS inhibit to 82% and 73% of respective controls, both
p < 0.001; not significantly different. In 8
F, strain Af293, HHQ and PQS inhibit to 87% and 40% of respective controls,
p < 0.05 and
p < 0.001, respectively. HHQ and PQS effect differ by
p < 0.001.
![Jof 07 00343 g008]()
Figure 9.
PQS and C9-PQS inhibit 10AF growth on agar. Effects of PQS, 3Cl-PQS (3Cl), NH2-PQS (NH2), HHQ, and C1-, C3-, C5-, C9-PQS were tested against 10AF growth on RPMI agar. All molecules were tested at 12.5 μg/mL. Only C9-PQS and PQS inhibited significantly (both p < 0.001), compared to control. PQS was more inhibitory compared to C1-PQS, C3-PQS, C5-PQS (all p < 0.001), and to NH2-PQS, 3Cl-PQS, HHQ (all p < 0.05). PQS and C9-PQS inhibited growth equally.
Figure 9.
PQS and C9-PQS inhibit 10AF growth on agar. Effects of PQS, 3Cl-PQS (3Cl), NH2-PQS (NH2), HHQ, and C1-, C3-, C5-, C9-PQS were tested against 10AF growth on RPMI agar. All molecules were tested at 12.5 μg/mL. Only C9-PQS and PQS inhibited significantly (both p < 0.001), compared to control. PQS was more inhibitory compared to C1-PQS, C3-PQS, C5-PQS (all p < 0.001), and to NH2-PQS, 3Cl-PQS, HHQ (all p < 0.05). PQS and C9-PQS inhibited growth equally.
Table 1.
Alkyl quinolones.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







