
The Metarhizium anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori



Abstract
:1. Introduction
2. Materials and Methods
2.1. Destruxin A (DA)
2.2. Bio-Layer Interferometry (BLI)
2.3. Insect Two-Hybrid (I2H)
2.4. RNAi Bioassay
3. Results
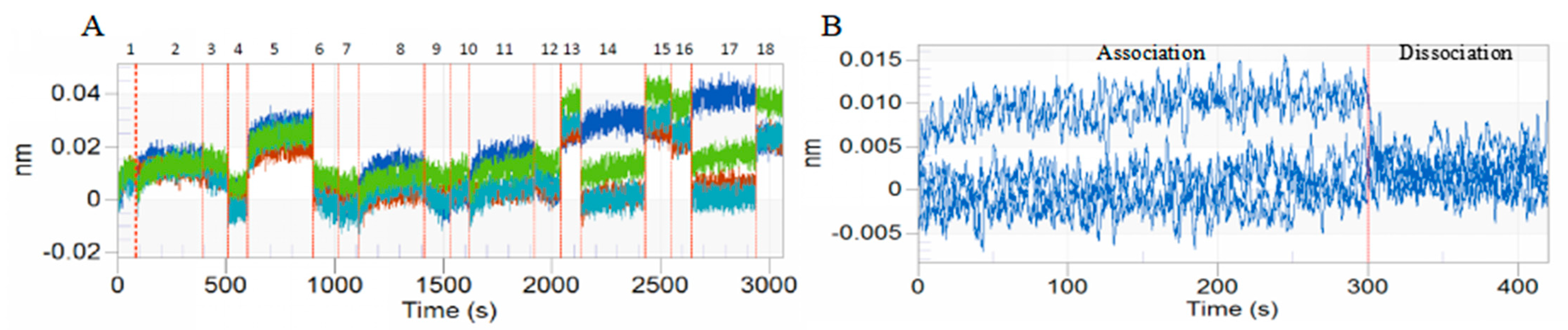
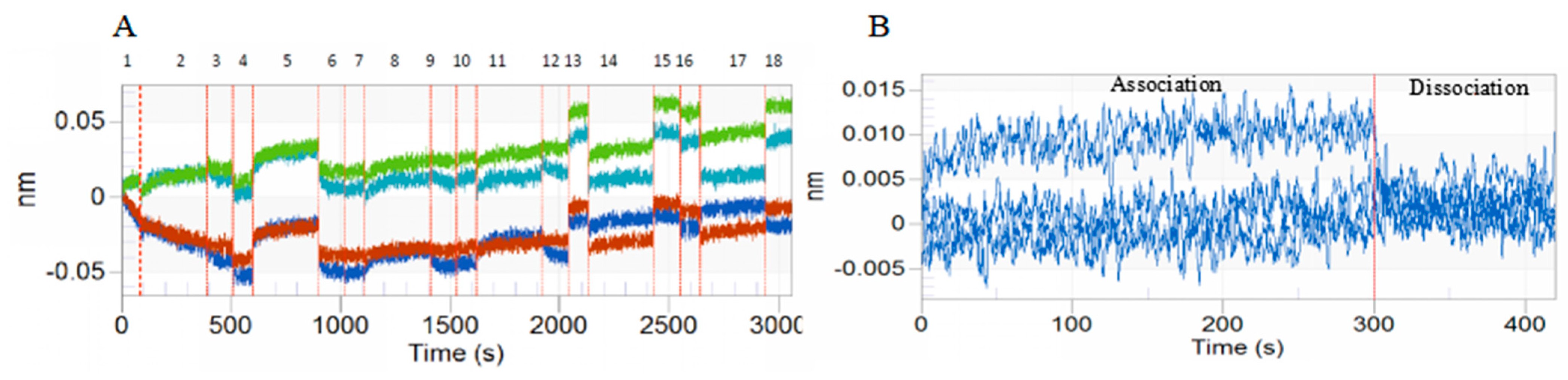
3.1. BLI of DA with BmTEME214 and BmSEC23
3.2. I2H of DA with BmTMEM214 and BmSEC23
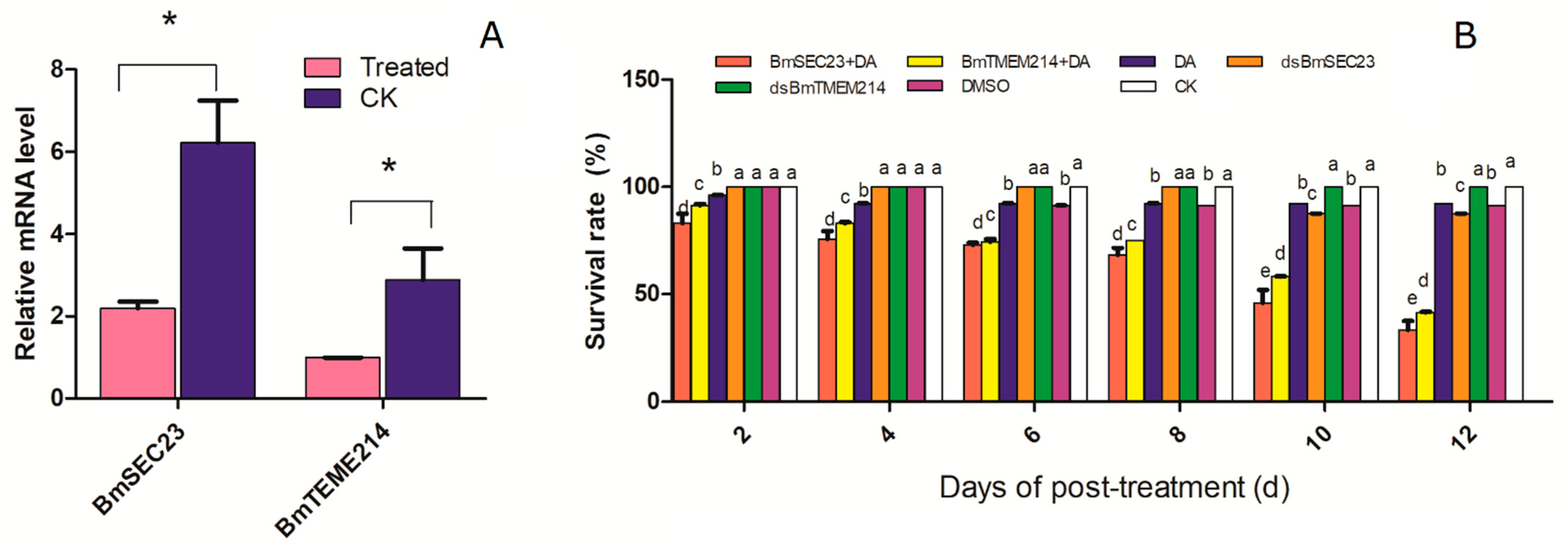
3.3. Toxicity of DA against Silkworm under RNAi
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arthurs, S.; Dara, S.K. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, I.; Fernandes, E.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Jaronski, S.T. Opportunities for microbial control of pulse crop pests. Ann. Entomol. Soc. Am. 2018, 111, 228–237. [Google Scholar] [CrossRef]
- Liu, B.L.; Tzeng, Y.M. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Zaharia, L.I.; Ward, D.E. The destruxins: Synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 2002, 59, 579–596. [Google Scholar] [CrossRef]
- Fan, J.Q.; Chen, X.R.; Hu, Q.B. Effects of destruxin A on hemocytes morphology of Bombyx mori. J. Integr. Agric. 2013, 12, 1042–1048. [Google Scholar] [CrossRef]
- Pal, S.; Leger, R.J.S.; Wu, L.P. Fungal peptide destruxin a plays a specific role in suppressing the innate immune response in Drosophila melanogaster. J. Biol. Chem. 2007, 282, 8969–8977. [Google Scholar] [CrossRef] [Green Version]
- Cavelier, F.; Verducci, J.; Andre, F.; Haraux, F.; Sigalat, C.; Traris, M.; Vey, A. natural cyclopeptides as leads for novel pesticides: tentoxin and destruxin. Pestic. Sci. 1998, 52, 81–89. [Google Scholar] [CrossRef]
- Lomenick, B.; Hao, R.; Jonai, N.; Chin, R.M.; Aghajan, M.; Warburton, S.; Wang, J.; Wu, R.P.; Gomez, F.; Loo, J.A.; et al. Target identification using drug affinity responsive target stability (DARTS). Proc. Natl. Acad. Sci. USA 2009, 106, 21984–21989. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wei, J.; Li, Y.; He, X.; Zhou, Q.; Yan, J.; Zhang, J.; Liu, Y.; Liu, Y.; Shu, H.-B. Transmembrane protein 214 (TMEM214) mediates endoplasmic reticulum stress-induced caspase 4 enzyme activation and apoptosis. J. Biol. Chem. 2013, 288, 17908–17917. [Google Scholar] [CrossRef] [Green Version]
- Paccaud, J.P.; Reith, W.; Carpentier, J.L.; Ravazzola, M.; Amherdt, M.; Schekman, R.; Orci, L. Cloning and functional characterization of mammalian homologues of the COPII component Sec23. Mol. Biol. Cell 1996, 7, 1535–1546. [Google Scholar] [CrossRef] [Green Version]
- Mancias, J.D.; Goldberg, J. Structural basis of cargo membrane protein discrimination by the human COPII coat machinery. EMBO J. 2008, 27, 2918–2928. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xu, X.; Hu, J.; Jin, F.; Hu, Q.; Sun, Q.; Yu, X.; Ren, S. Toxicity and differential protein analysis following destruxin A treatment of Spodoptera litura (Lepidoptera: Noctuidae) SL-1 cells. Toxicon 2011, 58, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Lee, B.; Lee, A.S. Endoplasmic reticulum stress-induced apoptosis—Multiple pathways and activation of p53-up-regulated modulator of apoptosis (PUMA) and NOXA by p53. J. Biol. Chem. 2006, 281, 7260–7270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.B.; Ren, S.-X.; Wu, J.-H.; Chang, J.-M.; Musa, P.D. Investigation of destruxin A and B from 80 Metarhizium strains in China, and the optimization of cultural conditions for the strain MaQ10. Toxicon 2006, 48, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Hu, W.N.; Xiao, M.M.; Ou, S.Y.; Hu, Q.B. Destruxin A induces and binds HSPs in Bombyx mori Bm12 cells. J. Agric. Food Chem. 2017, 65, 9850–9854. [Google Scholar] [CrossRef]
- Wang, J.; Hu, W.; Hu, Q. BmTudor-sn is a binding protein of destruxin A in silkworm Bm12 cells. Toxins 2019, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, J.; Weng, Q.; Hu, Q. Effects of destruxin A on silkworm’s immunophilins. Toxins 2019, 11, 349. [Google Scholar] [CrossRef] [Green Version]
- Parmigiani, A.; Nourbakhsh, A.; Ding, B.; Wang, W.; Kim, Y.C.; Akopiants, K.; Guan, K.L.; Karin, M.; Budanov, A.V. Sestrins inhibit mTORC1 kinase activation through the GATOR complex. Cell Rep. 2014, 9, 1281–1291. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Chantranupong, L.; Knockenhauer, K.E.; Schwartz, T.U.; Sabatini, D.M. Mechanism of arginine sensing by CASTOR1 upstream of mTORC1. Nat. Rev. Drug Discov. 2016, 536, 229–233. [Google Scholar]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Stagg, S.M.; Gurkan, C.; Fowler, D.M.; LaPointe, P.; Foss, T.R.; Potter, C.S.; Carragher, B.; Balch, W.E. Structure of the Sec13/31 COPII coat cage. Nature 2006, 439, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Stagg, S.M.; LaPointe, P.; Razvi, A.; Gurkan, C.; Potter, C.S.; Carragher, B.; Balch, W.E. Structural basis for cargo regulation of COPII coat assembly. Cell 2008, 134, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, N.; O’Donnell, J.; Stagg, S.M. The Structure of the Sec13/31 COPII Cage Bound to Sec23. J. Mol. Biol. 2012, 420, 324–334. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Protein | Interacting Protein | Interaction Score (String) | ||||
|---|---|---|---|---|---|---|
| Name | ID (NCBI) | Description | Name | ID (NCBI) | Description | |
| BmTMEM214 | XP_004933467.1 | transmembrane protein 214 | BmSEC13L | NP_001040420.1 | SEC13-like protein | 0.578 |
| BmSEC23 | XP_012553105.1 | protein transport protein SEC23A isoform X2 | BmSEC13 | XP_004923349.1 | Protein SEC13 homolog | 0.995 |
| Protein | KD (µM) a | KON (1/µMs) b | KOFF (1/s) c | Full R2 |
|---|---|---|---|---|
| BmTMEM214 | 0.286 | 0.184 | 0.0528 | 0.754 |
| BmSEC23 | 0.291 | 1.50 | 0.436 | 0.935 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, F.; Xiao, M.; Berestetskiy, A.; Hu, Q. The Metarhizium anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori. J. Fungi 2021, 7, 460. https://doi.org/10.3390/jof7060460
Yin F, Xiao M, Berestetskiy A, Hu Q. The Metarhizium anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori. Journal of Fungi. 2021; 7(6):460. https://doi.org/10.3390/jof7060460
Chicago/Turabian StyleYin, Fei, Miaomiao Xiao, Alexander Berestetskiy, and Qiongbo Hu. 2021. "The Metarhizium anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori" Journal of Fungi 7, no. 6: 460. https://doi.org/10.3390/jof7060460
APA StyleYin, F., Xiao, M., Berestetskiy, A., & Hu, Q. (2021). The Metarhizium anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori. Journal of Fungi, 7(6), 460. https://doi.org/10.3390/jof7060460