
Regulation of Copper Metabolism by Nitrogen Utilization in Saccharomyces cerevisiae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media, and Culture Conditions
2.2. Plasmid Construction
2.3. Nitrogen Starvation
2.4. Northern Blot Assay
2.5. Iron Uptake Assay
2.6. Western Blot Assay
2.7. Statistical Analysis
3. Results
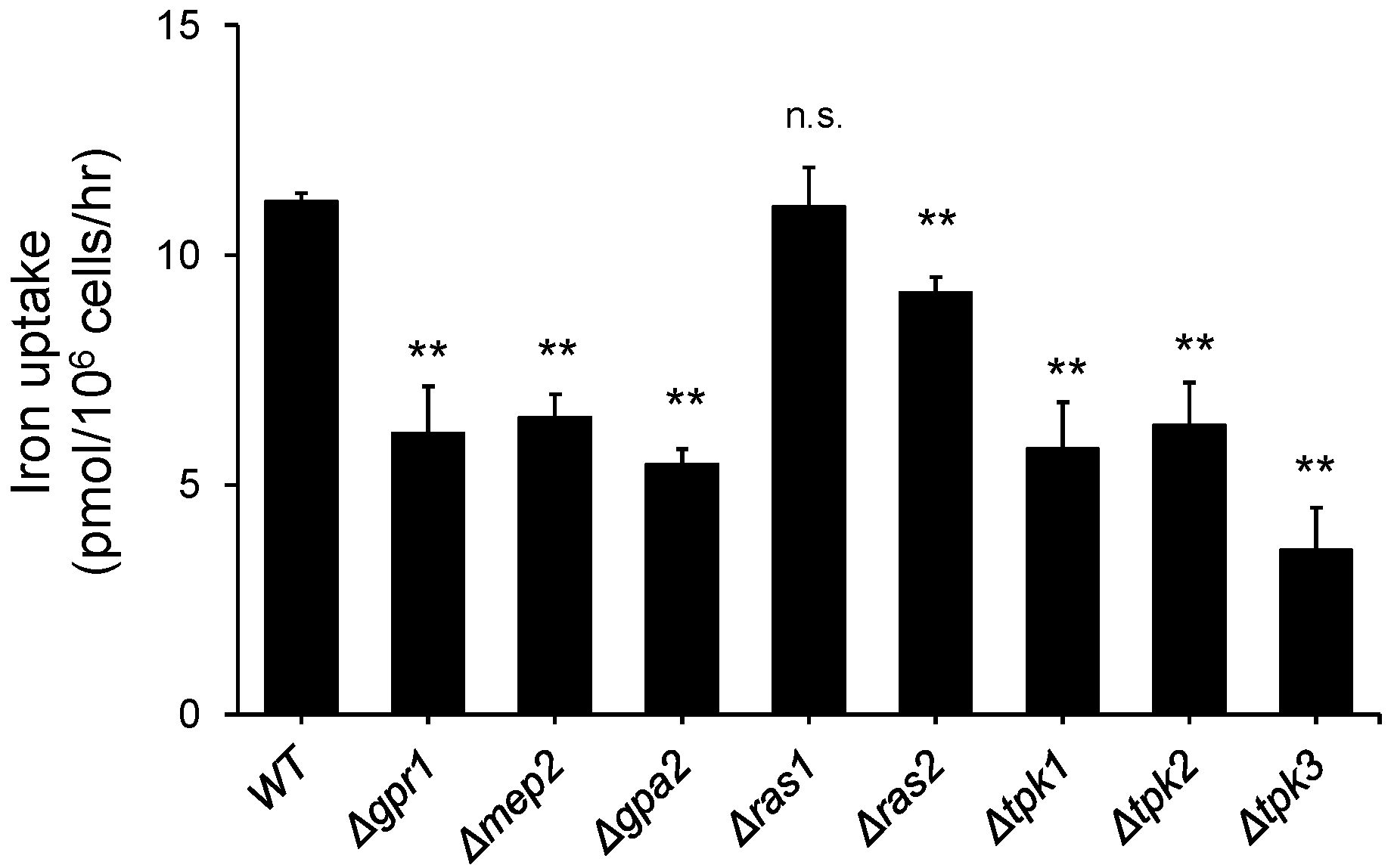
3.1. Deletion of the Genes Involved in Glucose and Nitrogen Metabolism Resulted in a Decrease in Iron Uptake
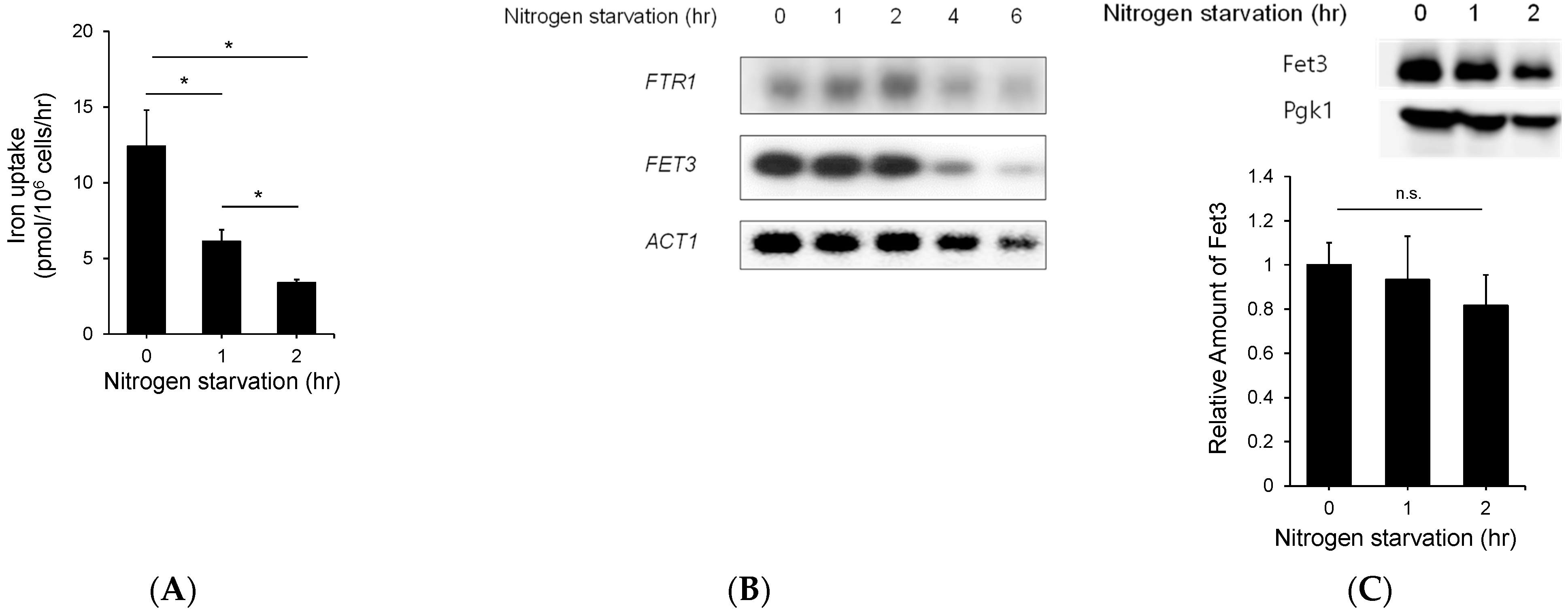
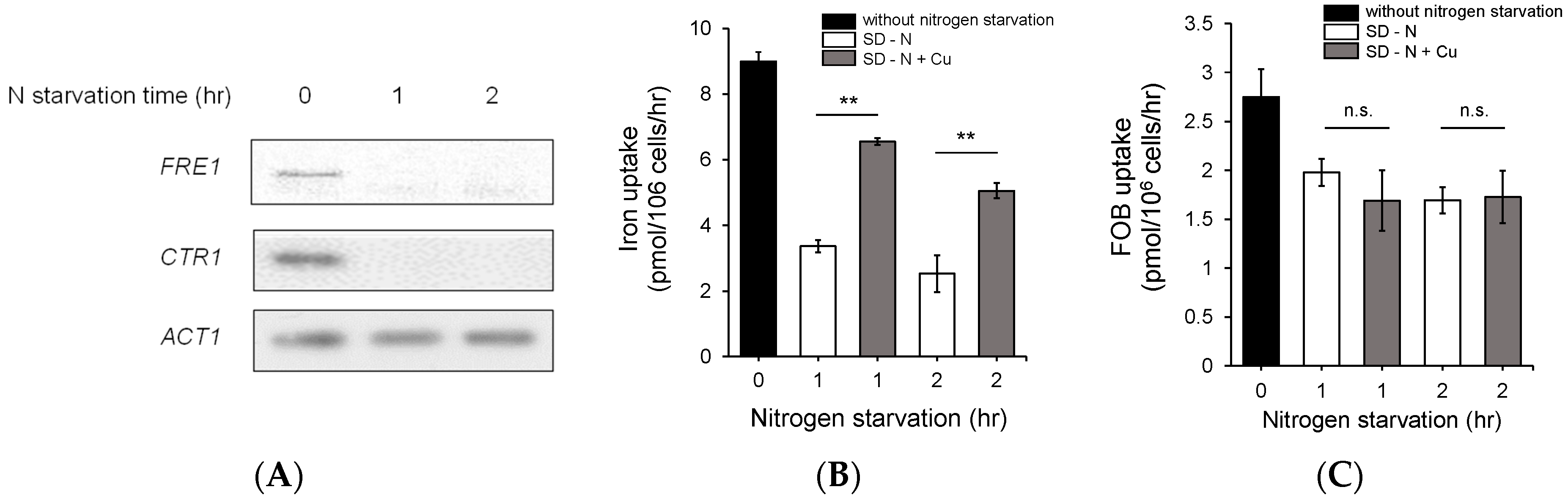
3.2. Nitrogen Starvation Downregulated the Expression of Genes Involved in Copper Metabolism
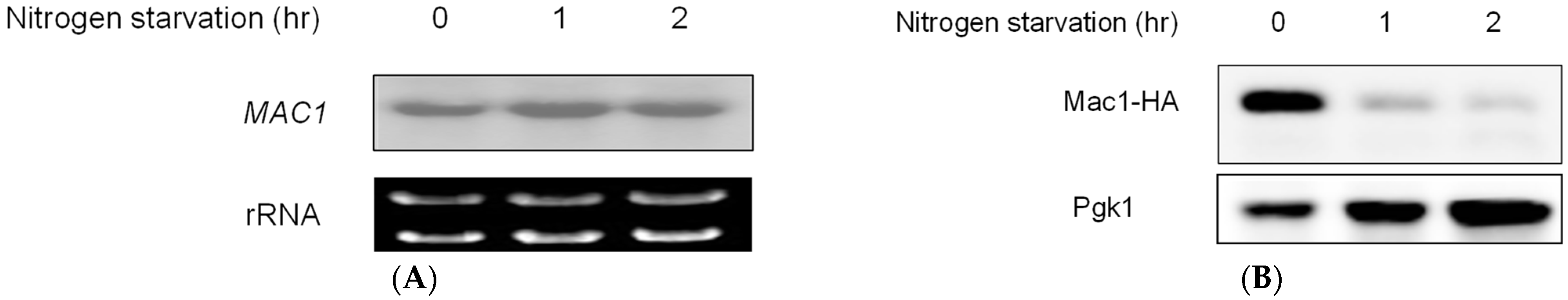
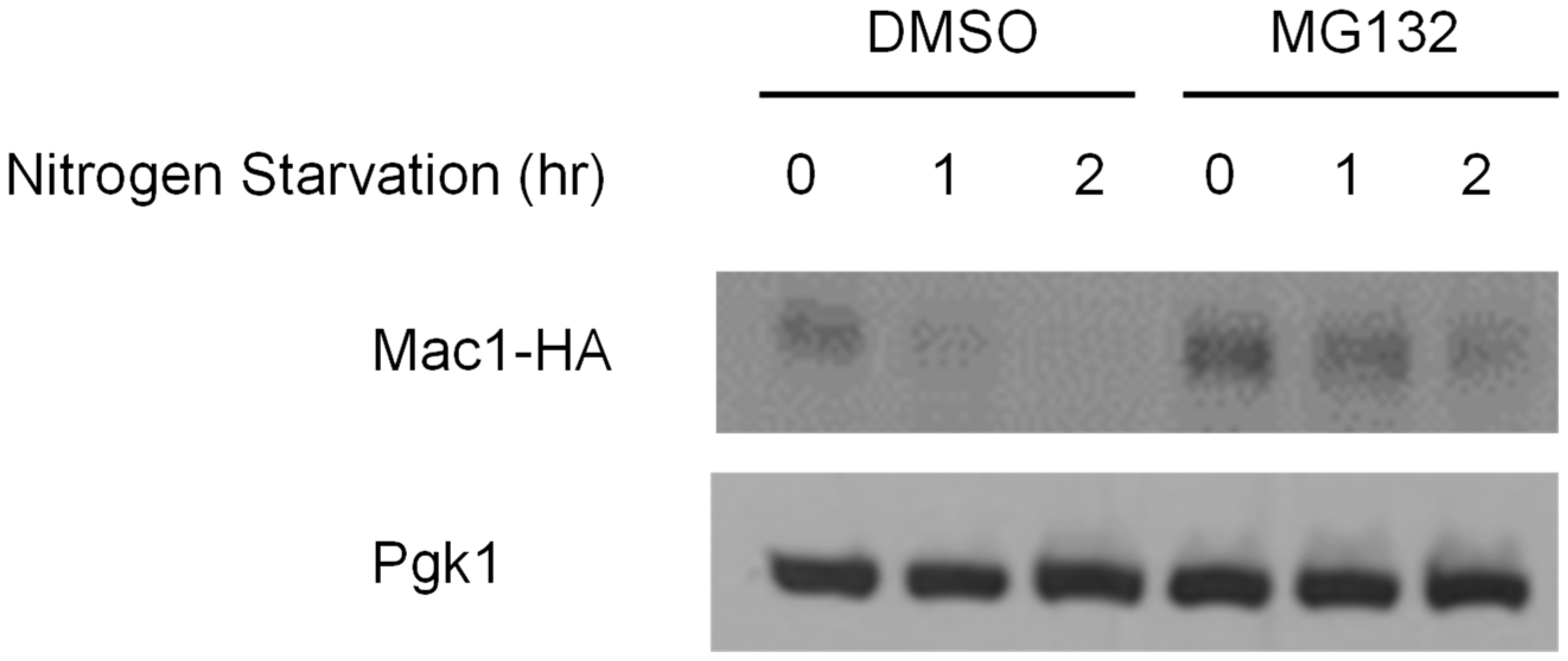
3.3. Nitrogen Starvation Downregulated Mac1 Expression at the Translational Level
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef]
- Bird, A.J. Cellular sensing and transport of metal ions: Implications in micronutrient homeostasis. J. Nutr. Biochem. 2015, 26, 1103–1115. [Google Scholar] [CrossRef]
- Bleackley, M.R.; MacGillivray, R.T.A. Transition metal homeostasis: From yeast to human disease. BioMetals 2011, 24, 785–809. [Google Scholar] [CrossRef]
- Robinson, J.R.; Isikhuemhen, O.S.; Anike, F.N. Fungal-Metal Interactions: A Review of Toxicity and Homeostasis. J. Fungi 2021, 7, 225. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, H.; Kolkowska, P.; Watly, J.; Krzywoszynska, K.; Potocki, S. General aspects of metal toxicity. Curr. Med. Chem. 2014, 21, 3721–3740. [Google Scholar] [CrossRef]
- Philpott, C.C.; Leidgens, S.; Frey, A.G. Metabolic remodeling in iron-deficient fungi. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2012, 1823, 1509–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, A.M.; Ramos-Alonso, L.; Alepuz, P.; Puig, S.; Martínez-Pastor, M.T. Global translational repression induced by iron deficiency in yeast depends on the Gcn2/eIF2α pathway. Sci. Rep. 2020, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Shakoury-Elizeh, M.; Protchenko, O.; Berger, A.; Cox, J.; Gable, K.; Dunn, T.M.; Prinz, W.A.; Bard, M.; Philpott, C.C. Metabolic Response to Iron Deficiency in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 14823–14833. [Google Scholar] [CrossRef] [Green Version]
- Felice, M.R.; De Domenico, I.; Li, L.; Ward, D.M.; Bartok, B.; Musci, G.; Kaplan, J. Post-transcriptional Regulation of the Yeast High Affinity Iron Transport System. J. Biol. Chem. 2005, 280, 22181–22190. [Google Scholar] [CrossRef] [Green Version]
- Dix, D.R.; Bridgham, J.T.; Broderius, M.A.; Byersdorfer, C.A.; Eide, D.J. The FET4 gene encodes the low affinity Fe(II) transport protein of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 26092–26099. [Google Scholar] [CrossRef]
- Askwith, C.; Eide, D.; Van Ho, A.; Bernard, P.S.; Li, L.; Davis-Kaplan, S.; Sipe, D.M.; Kaplan, J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell 1994, 76, 403–410. [Google Scholar] [CrossRef]
- Lin, S.-J.; Pufahl, R.A.; Dancis, A.; O’Halloran, T.V.; Culotta, V.C. A Role for the Saccharomyces cerevisiae ATX1 Gene in Copper Trafficking and Iron Transport. J. Biol. Chem. 1997, 272, 9215–9220. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Jiang, Y.; Yang, Y.; Peng, Y.; Li, C. Copper metabolism in Saccharomyces cerevisiae: An update. BioMetals 2021, 34, 3–14. [Google Scholar] [CrossRef]
- Knight, S.; Labbe, S.; Kwon, L.F.; Kosman, D.J.; Thiele, D.J. A widespread transposable element masks expression of a yeast copper transport gene. Genes Dev. 1996, 10, 1917–1929. [Google Scholar] [CrossRef] [Green Version]
- Hua, H.; Günther, V.; Georgiev, O.; Schaffner, W. Distorted copper homeostasis with decreased sensitivity to cisplatin upon chaperone Atox1 deletion in Drosophila. BioMetals 2011, 24, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Arnesano, F.; Banci, L.; Bertini, I.; Cantini, F.; Ciofi-Baffoni, S.; Huffman, D.L.; O’Halloran, T.V. Characterization of the Binding Interface between the Copper Chaperone Atx1 and the First Cytosolic Domain of Ccc2 ATPase * 210. J. Biol. Chem. 2001, 276, 41365–41376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Labbé, S.; Peña, M.M.O.; Thiele, D.J. Copper Differentially Regulates the Activity and Degradation of Yeast Mac1 Transcription Factor. J. Biol. Chem. 1998, 273, 1277–1280. [Google Scholar] [CrossRef] [Green Version]
- Keller, G.; Gross, C.; Kelleher, M.; Winge, D.R. Functional Independence of the Two Cysteine-rich Activation Domains in the Yeast Mac1 Transcription Factor. J. Biol. Chem. 2000, 275, 29193–29199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Du, W.; Zeng, Q.; Long, N.; Dai, C.; Lu, L. Cu-sensing transcription factor Mac1 coordinates with the Ctr transporter family to regulate Cu acquisition and virulence in Aspergillus fumigatus. Fungal Genet. Biol. 2017, 107, 31–43. [Google Scholar] [CrossRef]
- Marvin, M.E.; Mason, R.P.; Cashmore, A.M. The CaCTR1 gene is required for high-affinity iron uptake and is transcriptionally controlled by a copper-sensing transactivator encoded by CaMAC1. Microbiology 2004, 150, 2197–2208. [Google Scholar] [CrossRef]
- Martins, T.S.; Costa, V.; Pereira, C. Signaling pathways governing iron homeostasis in budding yeast. Mol. Microbiol. 2018, 109, 422–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, F.; De Winde, J.H.; Lemaire, K.; Boles, E.; Thevelein, J.M.; Winderickx, J. Glucose-induced cAMP signalling in yeast requires both a G-protein coupled receptor system for extracellular glucose detection and a separable hexose kinase-dependent sensing process. Mol. Microbiol. 2000, 38, 348–358. [Google Scholar] [CrossRef]
- Robertson, L.S.; Causton, H.C.; Young, R.A.; Fink, G.R. The yeast A kinases differentially regulate iron uptake and respiratory function. Proc. Natl. Acad. Sci. USA 2000, 97, 5984. [Google Scholar] [CrossRef] [Green Version]
- Schothorst, J.; Zeebroeck, G.V.; Thevelein, J.M. Identification of Ftr1 and Zrt1 as iron and zinc micronutrient transceptors for activation of the PKA pathway in Saccharomyces cerevisiae. Microb. Cell 2017, 4, 74–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M.C.; Heitman, J. The MEP2 ammonium permease regulates pseudohyphal differentiation in Saccharomyces cerevisiae. EMBO J. 1998, 17, 1236–1247. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Starke, S.; Li, Y.; Sethupathi, S.; Kung, G.; Dodhiawala, P.; Wang, Y. Nitrogen Starvation-induced Phosphorylation of Ras1 Protein and Its Potential Role in Nutrient Signaling and Stress Response. J. Biol. Chem. 2016, 291, 16231–16239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.W.; Ferea, T.; Rashford, J.; Ardon, O.; Brown, P.O.; Botstein, D.; Kaplan, J.; Philpott, C.C. Desferrioxamine-mediated iron uptake in Saccharomyces cerevisiae-Evidence for two pathways of iron uptake. J. Biol. Chem. 2000, 275, 10709–10715. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.C.; Wu, E.; Sailer, C.; Jando, J.; Styles, E.; Eisenkolb, I.; Kuschel, M.; Bitschar, K.; Wang, X.; Huang, L.; et al. Ubiquitin orchestrates proteasome dynamics between proliferation and quiescence in yeast. Mol. Biol. Cell 2017, 28, 2479–2491. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.; Walcheck, B.; Migaki, G.I.; Jutila, M.A.; Kishimoto, T.K. Calmodulin Regulates L-Selectin Adhesion Molecule Expression and Function through a Protease-Dependent Mechanism. Cell 1998, 92, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Lee Do, H.; Goldberg Alfred, L. Proteasome Inhibitors Cause Induction of Heat Shock Proteins and Trehalose, Which Together Confer Thermotolerance inSaccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 30–38. [Google Scholar] [CrossRef] [Green Version]
- MacGurn, J.A.; Hsu, P.-C.; Smolka, M.B.; Emr, S.D. TORC1 Regulates Endocytosis via Npr1-Mediated Phosphoinhibition of a Ubiquitin Ligase Adaptor. Cell 2011, 147, 1104–1117. [Google Scholar] [CrossRef] [Green Version]
- An, Z.; Tassa, A.; Thomas, C.; Zhong, R.; Xiao, G.; Fotedar, R.; Tu, B.P.; Klionsky, D.J.; Levine, B. Autophagy is required for G₁/G₀ quiescence in response to nitrogen starvation in Saccharomyces cerevisiae. Autophagy 2014, 10, 1702–1711. [Google Scholar] [CrossRef] [Green Version]
- Willis, S.D.; Hanley, S.E.; Beishke, T.; Tati, P.D.; Cooper, K.F. Ubiquitin-proteasome-mediated cyclin C degradation promotes cell survival following nitrogen starvation. Mol. Biol. Cell 2020, 31, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Bae, D.-H.; Jansson, P.J.; Richardson, D.R. The mechanistic role of chemically diverse metal ions in the induction of autophagy. Pharmacol. Res. 2017, 119, 118–127. [Google Scholar] [CrossRef]
- Klausner, R.D.; Dancis, A. A genetic approach to elucidating eukaryotic iron metabolism. FEBS Lett. 1994, 355, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.D.; Logeman, B.L.; Thiele, D.J. Copper Acquisition and Utilization in Fungi. Annu. Rev. Microbiol. 2017, 71, 597–623. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. JBIC J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.; Seo, H.; Lee, M.-G.; Yun, C.-W. Regulation of Copper Metabolism by Nitrogen Utilization in Saccharomyces cerevisiae. J. Fungi 2021, 7, 756. https://doi.org/10.3390/jof7090756
Kang S, Seo H, Lee M-G, Yun C-W. Regulation of Copper Metabolism by Nitrogen Utilization in Saccharomyces cerevisiae. Journal of Fungi. 2021; 7(9):756. https://doi.org/10.3390/jof7090756
Chicago/Turabian StyleKang, Suzie, Hyewon Seo, Min-Gyu Lee, and Cheol-Won Yun. 2021. "Regulation of Copper Metabolism by Nitrogen Utilization in Saccharomyces cerevisiae" Journal of Fungi 7, no. 9: 756. https://doi.org/10.3390/jof7090756
APA StyleKang, S., Seo, H., Lee, M.-G., & Yun, C.-W. (2021). Regulation of Copper Metabolism by Nitrogen Utilization in Saccharomyces cerevisiae. Journal of Fungi, 7(9), 756. https://doi.org/10.3390/jof7090756