
Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review




Abstract
:1. Introduction
2. Worldwide Association of F. verticillioides
3. Management of F. verticillioides
4. Physical Methods for Management of F. verticillioides
5. Chemical Methods for Management of F. verticillioides
6. Limitations of Physical and Chemical Methods
7. Management of F. verticillioides by Natural Clay
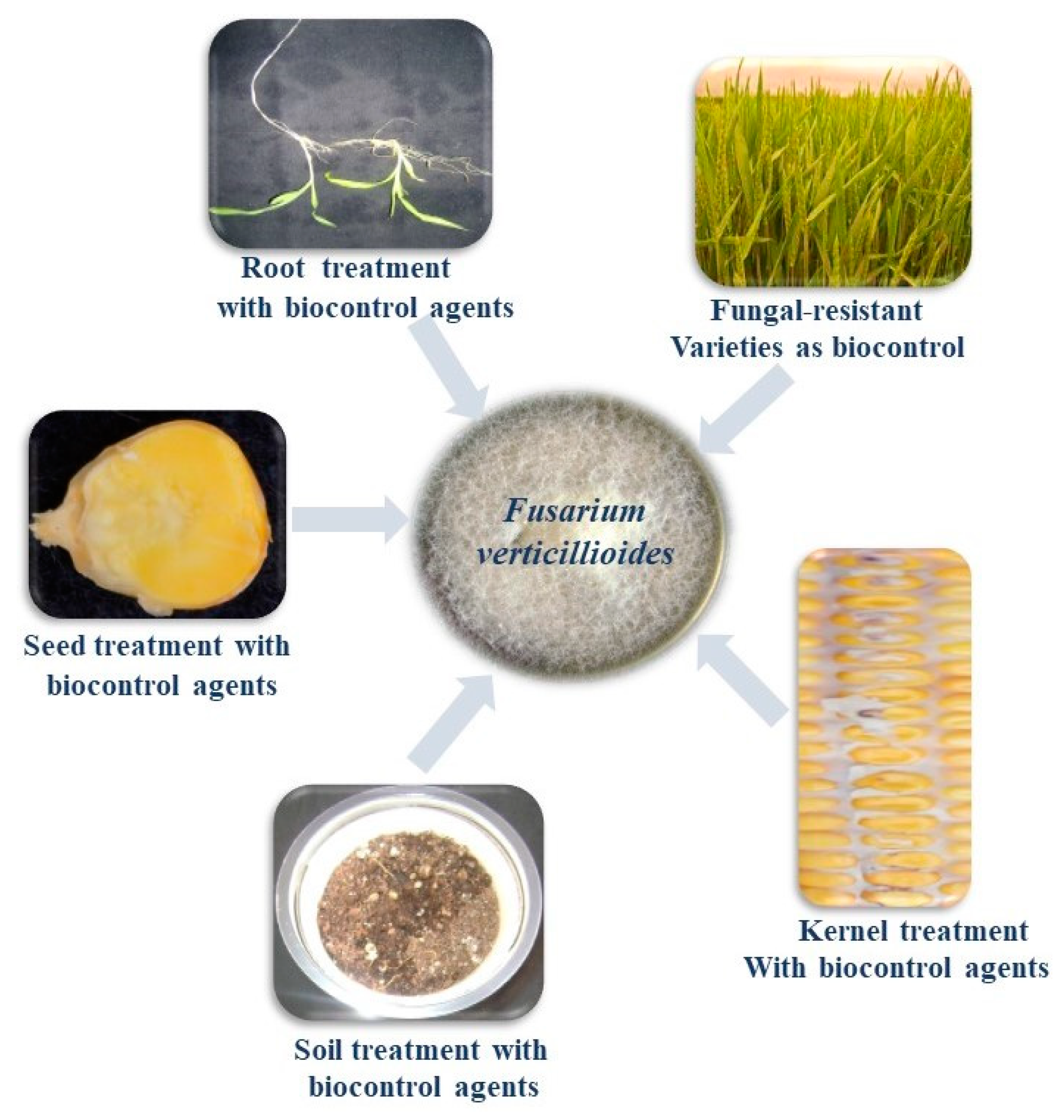
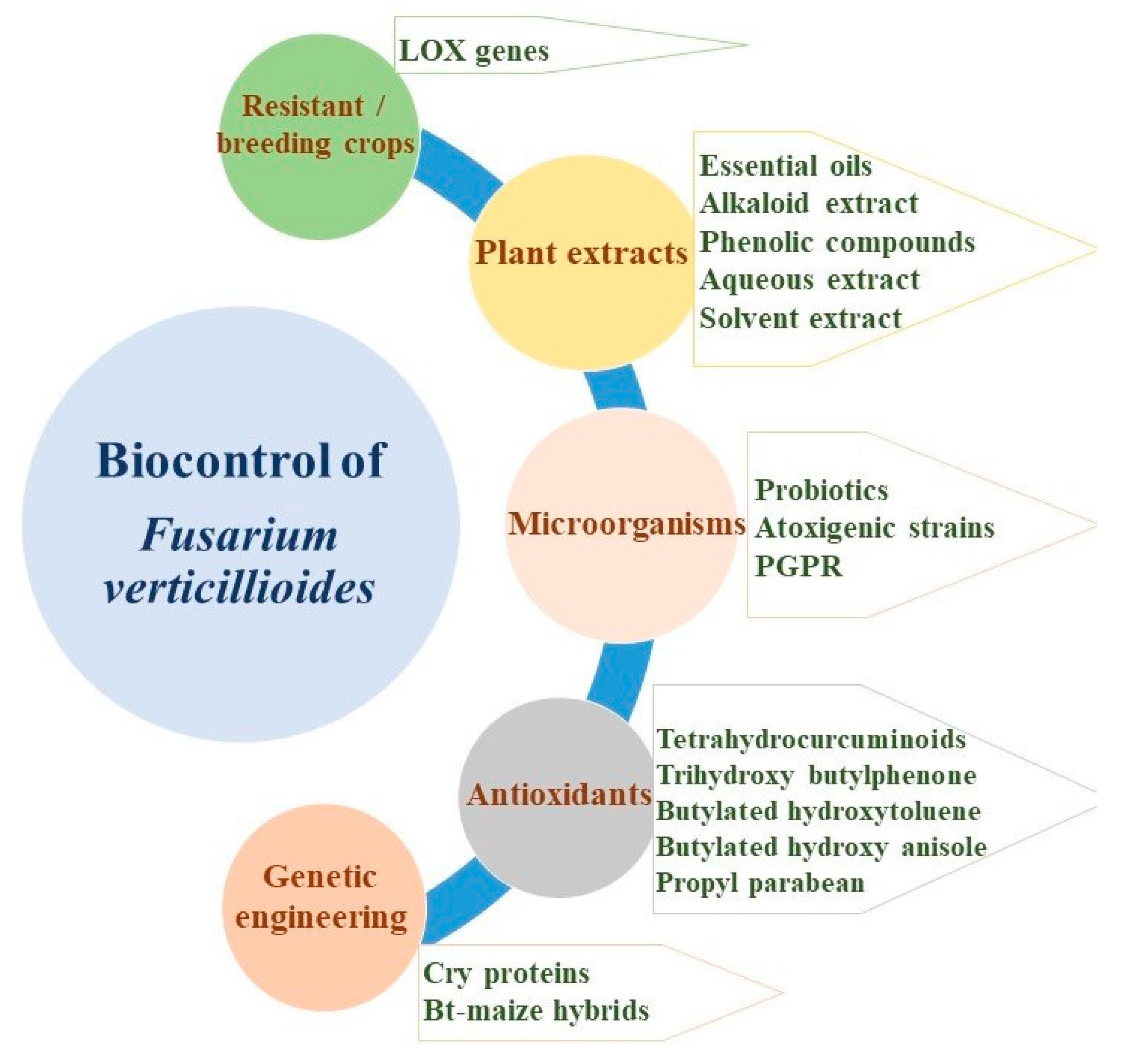
8. Biological Control of F. verticillioides
8.1. Microorganisms as Biocontrol Agents
8.1.1. Bacteria as Biocontrol Agents

{kind=link}
{kind=link}
| Serial Number | Test Organism | Methods Used for Screening | Targeted Feature of F. verticillioides | Percent of Reduction | Reference |
|---|---|---|---|---|---|
| Microorganisms | |||||
| 1. | Bacillus subtilis | Antifungal activity | Fungal growth | 50% | [91] |
| 2. | Lactobacillus rhamnococcus | Antifungal activity | Mold growth | 78–92% | [90,92] |
| 3. | Saccharomyces cerevisiae | Antifungal activity | Mold growth and FB1 | 77–89% | |
| 4. | Pediococcus pentosaceus | Antifungal activity and spectrum | Zone of inhibition | 89% | [93] |
| 5. | Enterococcus casseliflavus M4A | Antifungal activity | Fungal growth | 100% | [94] |
| 6. | Trichoderma viridae | Antifungal activity | Zone of inhibition | 90% | [70,95,96] |
| 7. | Trichoderma harzianum | Antifungal activity | Zone of inhibition | < 70% | |
| 8. | Pseudomonas solanaceacum | Antifungal activity | Zone of inhibition | 70% | [81] |
| 9. | Mixture of E. cloacae and M. oleovorans; mixture of P. solanaceacum and B. subtilis | Antibiosis | Root colonization | 50% | [87] |
| Antioxidants | |||||
| 10. | Butylated hydroxyanisole (BHA) | Water activity | Mycelial growth | 94–98% | [68] |
| 11. | Propylparabean (PP) | 100% | |||
| 12. | Trihydroxybutyrophenone (THBP) | >85% | [97] | ||
| 13. | Butylated hydroxytoluene (BHT) | 95% | |||
| 14. | Tetrahydrocurcuminoids (THC) | Antifungal activity | Fungal conidial growth | 100% | [98] |
| Phenolic Compounds and Plant Extracts | |||||
| 15. | Geranial, eugenol, and singerone from Zingiber officinale | Poisoned food technique | mycelial growth | 100% | [99] |
| 16. | Carvacrol, eugenol, 2-hexanal from oregano, thyme, cinnamon, clove, fruits, and vegetables | Antifungal activity | Conidial germination and mycelial growth | 37–97% | [100] |
| 17. | Chitin-binding protein from Ginko biloba, | Antifungal activity | Mycelial growth | >50% | [101] |
| 18. | Vanillic acid and caffeic acid | Antifungal activity | Fungal growth | 80% 100% | [102,103] |
| 19. | Chlorophorin, iroko, maakianin, and ferulic acid | Agar-well diffusion and HPLC | Fungal growth and FB1 | 88–94% | [103] |
| 20. | Flavonoids, phenolic acid, and terpine-rich ethanol extracts from Equisetum arvense and Stevia rebaudiana | Antifungal activity | Fungal growth | 79% | [104] |
| 21. | Aqueous extract, methanol extract, and alkaloid extract from Prosopis juliflora | Poisoned food technique | Mycelial growth | 100% | [105] |
| 22. | 1,8-Cineole from Rosnainum officinalis | Antifungal activity | Conidial production | 53.48% | [106] |
| 23. | Eugenol, methyl eugenol, and tumerone from Syzygium aromaticum, Pimenta dioica, and C. longa | Fungal growth | 40–80% | ||
8.1.2. Probiotics as Biocontrol Agent
8.1.3. Fungi as Biocontrol Agents
8.1.4. PGPR as Biocontrol Agents
8.1.5. Mycoviruses as Biocontrol Agents
8.1.6. Non-Toxigenic Strains as Biocontrol Agents
8.2. Antioxidants as Biocontrol Agents
| Serial Number | Biocontrol Agents | Detection Method | Percent of FB1 Reduction | Level of Study | Reference |
|---|---|---|---|---|---|
| Microorganisms | |||||
| 1. | Bacillus subtilis | HPLC | 50% | In vitro | [87] |
| 2. | Bacillus amyloliquifaciens | HPLC | >70% | Field study | [82,89] |
| 3. | Microbacterium oleovarans | HPLC | Field study | ||
| 4. | Enterobacter hormacchei | HPLC | Field study | ||
| 5. | Lactobacillus rhamnococcus | HPLC | 78–92% | In vitro | [90,92] |
| 6. | Saccharomyces cerevisiae | HPLC | 77–89% | In vitro | |
| 7. | Pseudomonas solanaceacum | HPLC | 70–100% | In vitro | [68,81,87] |
| 8. | Mixture of E. cloacae/M. oleovorans; mixture of P. solanaceacum/B. subtilis | HPLC | 100% | Field study | [91] |
| Antioxidants | |||||
| 9. | Butylated hydroxyanisole (BHA) | HPLC | 100% | In vitro | [68,117] |
| 10. | Propylparabean (PP) | 94–98% | |||
| 11. | Trihydroxybutyrophenone (THBP) | 94–98% | [97] | ||
| 12. | Butylated hydroxytoluene (BHT) | ||||
| 13. | 3,6,7-trihydroxy-α-tetralone (TT) | HPLC | >95% | In vitro | [118] |
| Phenolic Compounds and Plant Extracts | |||||
| 14. | Ferulic acid | HPLC | 98–100% | In vitro | [103] |
| 15. | Vanillic acid, and caffeic acid | HPLC | 98–100% | In vitro | [102] |
| 16. | Acetonin | HPLC | 88.75% | In vitro | [94] |
| 17. | Acetone extract | HPLC | 96% | In vitro | [119] |
8.3. Plant Extracts as Biocontrol Agents
8.4. Plant-Based Essential Oils as Biocontrol Agents
8.5. Resistant Crops via Breeding as Biocontrol Methods
8.6. Genetic Engineering as Tools for Biocontrol
8.7. Commercially Available Products as Biocontrol Agents
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberts, J.F.; Van Zyl, W.H.; Gelderblom, W.C.A. Biologically Based Methods for Control of Fumonisin-Producing Fusarium Species and Reduction of the Fumonisins. Front. Microbiol. 2016, 7, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, J.I.; Wild, C.P.; Baan, R.; Gelderblom, W.C.A.; Miller, J.D.; Riley, R. Improving Public Health through Mycotoxin Control; International Agency for Res on Cancer (IARC) Scientific Publication No.; IARC Press: Lyon, France, 2012; 158p. [Google Scholar]
- Deepa, N.; Sreenivasa, M.Y. Fumonisin: A review on its Global Occurrence, Epidemiology, Toxicity and Detection. J. Vet. Med. Res. 2017, 4, 1093. [Google Scholar]
- Sreenivasa, M.; Diwakar, B.; Raj, A.P.C.; Dass, R.S.; Naidu, K.; Janardhana, G. Determination of toxigenic potential of Fusarium species occurring on sorghum and maize grains produced in Karnataka, India by using Thin Layer Chromatography. Int. J. Life Sci. 2012, 6, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.F.; Ledet, A.E.; Owens, D.L.; Rice, L.G.; Nelson, H.A.; Osweiler, G.D.; Wilson, T.M. Experimental Equine Leukoencephalomalacia, Toxic Hepatosis, and Encephalopathy Caused by Corn Naturally Contaminated with Fumonisins. J. Veter-Diagn. Investig. 1993, 5, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, B.M.; Harrison, L.R. Fumonisin-induced pulmonary edema and hydrothorax in swine. Mycopathologia 1992, 117, 79–82. [Google Scholar] [CrossRef]
- Voss, K.A.; Riley, R.T.; Snook, M.E.; Waes, J.G. Reproductive and sphingolipid metabolic effects of fumonisin B1 and its alka-line hydrolysis product in LM/Bc mice: Hydrolyzed fumonisin B1 did not cause neural tube defects. Toxicol. Sci. 2009, 112, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacic, S.; Pepeljnjak, S.; Petrinec, Z.; Klarić, M. Fumonisin B1 Neurotoxicity in Young Carp (Cyprinus Carpio L.). Arch. Ind. Hyg. Toxicol. 2009, 60, 419–426. [Google Scholar] [CrossRef]
- Thiel, P.G.; Marasas, W.F.O.; Sydenham, E.W.; Shephard, G.S.; Gelderblom, W.C.A. The implications of naturally occurring levels of fumonisins in corn for human and animal health. Mycopathologia 1992, 117, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, S.; Bidoli, E.; Baron, A.E.; La Vecchia, C. Maize and risk of cancers of the oral cavity, pharynx, and oesophagus in Northeastern Italy. J. Nat. Can. Inst. 1990, 82, 1407–1411. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.S.; Li, G.V. Simultaneous occurrence of fumonisin B1 and other mycotoxins in moldy corn collected from People’s Re-public of China in regions with high incidence of esophageal cancer. Appl. Environ. Microbiol. 1994, 60, 847–852. [Google Scholar]
- Alizadeh, A.M. Fumonisin B1 contamination of cereals and risk of esophageal cancer in a high-risk area in north eastern Iran. Asian Pac. J. Can. Prev. 2012, 13, 2625–2628. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer (IARC); World Health Organisation (WHO). Fumonisin B1. In IARC Mono-Graphs on the Evaluation of Carcinogenic Risks to Humans, Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; IARC Press: Lyon, France, 2002; Volume 82, pp. 301–366. [Google Scholar]
- Oren, L.; Ezrati, S.; Cohen, D.; Sharon, A. Early Events in the Fusarium verticillioides-Maize Interaction Characterized by Using a Green Fluorescent Protein-Expressing Transgenic Isolate. Appl. Environ. Microbiol. 2003, 69, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E.; Manandhar, G.; Plattner, R.D.; Maragos, C.M.; Shrestha, K.; McCormick, S. Occurrence of Fusarium Species and Mycotoxins in Nepalese Maize and Wheat and the Effect of Traditional Processing Methods on Mycotoxin Levels. J. Agric. Food Chem. 2000, 48, 1377–1383. [Google Scholar] [CrossRef]
- Ghiasian, S.A.; Rezayat, S.M.; Kord-Bacheh, P.; Maghsood, A.H.; Yazdanpanah, H.; Shephard, G.S.; van der Westhuizen, L.; Vismer, H.F.; Marasas, W.F. Fumonisin production by Fusarium species isolated from freshly harvested corn in Iran. Mycopathologia 2005, 159, 31–40. [Google Scholar] [CrossRef]
- Munkvold, G.; Stahr, H.M.; Logrieco, A.; Moretti, A.; Ritieni, A. Occurrence of fusaproliferin and beauvericin in Fusarium-contaminated livestock feed in Iowa. Appl. Environ. Microbiol. 1998, 64, 3923–3926. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, A.; Nejad, R.F.; Mofrad, N.N. F. verticillioides from sugarcan, vegetative compatability groups and pathogene-city. Plant Prot. Sci. 2012, 48, 80–84. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Busman, M.; Proctor, R.H.; Stessman, R. Wheat kernel black point and fumonisin contamination by Fusarium proliferatum. Food Addit. Cont. 2007, 24, 1131–1137. [Google Scholar] [CrossRef]
- Anthony, S.; Abeywickrama, K.; Dayananda, R.; Wijeratnam, S.W.; Arambewela, L. Fungal pathogens associated with banana fruit in Sri Lanka, and their treatment with essential oils. Mycopathologia 2004, 157, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Stephens, C.T.; de Vries, R.M.; Sink, K.C. Evaluation of Asparagus species for resistance to Fusarium oxysporoum and Fusarium verticillioides. Horticul. Sci. 1989, 24, 365–368. [Google Scholar]
- Corpas-Hervias, C.; Melero-Vara, J.M.; Molinero-Ruiz, L.; Zurera-Muñoz, C.; Basallote-Ureba, M.J. Characterization of Isolates of Fusarium spp. Obtained from Asparagus in Spain. Plant Dis. 2006, 90, 1441–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesso, T.; Claflin, L.E.; Tuinstra, M.R. Estimation of Combining Ability for Resistance to Fusarium Stalk Rot in Grain Sorghum. Crop Sci. 2004, 44, 1195–1199. [Google Scholar] [CrossRef]
- Rocha, L.D.O.; Reis, G.M.; Da Silva, V.N.; Braghini, R.; Teixeira, M.M.G.; Corrêa, B. Molecular characterization and fumonisin production by Fusarium verticillioides isolated from corn grains of different geographic origins in Brazil. Int. J. Food Microbiol. 2011, 145, 9–21. [Google Scholar] [CrossRef]
- Dass, R.S.; Sreenivasa, M.Y.; Charith Raj, A.P.; Janardhana, G.R. PCR-based assay for the rapid detection of Fumonisin-producing Fusarium species in Maize-based animal and poultry feeds in Karnataka, India. Arch. Phytopathol. Plant Prot. 2009, 42, 796–804. [Google Scholar] [CrossRef]
- Deepa, N.; Nagaraja, H.; Sreenivasa, M. Prevalence of fumonisin producing Fusarium verticillioides associated with cereals grown in Karnataka (India). Food Sci. Hum. Wellness 2016, 5, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.F.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. Eur. J. Plant. Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Covarelli, L.; Stifano, S.; Beccari, G.; Raggi, L.; Lattanzio, V.M.T.; Albertini, E. Characterization of Fusarium verticillioides strains isolated from maize in Italy: Fumonisin production, pathogenicity and genetic variability. Food Microbiol. 2012, 31, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Rahjoo, V.; Zad, J.; Javan-Nikkhah, M.; Gohari, A.M. Morphological and molecular identification of Fusarium isolated from maize ears in Iran. J. Plant Pathol. 2008, 90, 463–468. [Google Scholar]
- Sreenivasa, M.Y.; Gonzalez Jaen, M.T.; Dass, R.S.; Charithraj, A.P.; Janrdhana, G.R. A PCR based assay for the detetction and differentiation of potential fumonisin producing Fusarium verticillioides isolated from maize kernels. Food Biotechnol. 2008, 22, 160–170. [Google Scholar] [CrossRef]
- Qian, G.; Tang, L.; Lin, S.; Xue, K.S.; Mitchell, N.J.; Su, J.; Gelderblom, W.C.; Riley, R.T.; Phillips, T.D.; Wang, J.S. Sequential dietary exposure to aflatoxin B1 and fumonisin B1 in F3 44 rats increases liver preneoplastic changes indicative of a synergistic interaction. Food Chem. Toxicol. 2016, 95, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraja, H.; Chennappa, G.; Rao, K.P.C.; Prasad, G.M.; Sreenivasa, M.Y. Diversity of toxic and phytopathogenic Fusarium species occurring on cereals grown in Karnataka state, India. 3 Biotech 2016, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Xue, K.S.; Qian, G.; Lin, S.; Su, J.; Tang, L.; Gelderblom, W.C.; Riley, R.T.; Phillips, T.D.; Wang, J.S. Modulation of pre-neoplastic biomarkers induced by sequential aflatoxin B1 and fumnisin B1 exposure in F344 rat treated with UPSN clay. Food Chem. Toxicol. 2018, 114, 316–324. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on Agriculture, Food and Human health and their management strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasa, M.Y.; Dass, R.S.; Charithraj, A.P.; Janardhana, G.R. Molecular Detection of Fumonisin Producing Fusarium Species of freshly harvested maize kernels using Polymerase Chain Reaction (PCR). Taiwania-Taipei 2006, 51, 251–257. [Google Scholar]
- Deepa, N.; Sreenivasa, M. Biocontrol Strategies for Effective Management of Phytopathogenic Fungi Associated with Cereals. In New and Future Developments in Microbial Biotechnology and Bioengineering-Microbial Biotechnology in Agro-Sustainability; Elsevier: Amsterdam, The Netherlands, 2019; Chapter 13; pp. 177–189. [Google Scholar] [CrossRef]
- Deepa, N.; Charith Raj, A.P.; Sreenivasa, M.Y. Multiplex PCR method for the early detection of fumonisin producing Fusarium verticillioides. Food Biosci. 2016, 13, 84–88. [Google Scholar]
- Deepa, N.; Adkar-Purushothama, C.R.; Sreenivasa, M.Y. Nested PCR method for the early detection of fumonisin producing Fusarium verticillioides in pure cultures, cereal samples and plant parts. Food Biotechnol. 2016, 30, 18–29. [Google Scholar]
- Sreenivasa, M.Y. Fumonisin–A potential carcinogen is of global concern. Res. J. Biotechnol. 2012, 7, 1–2. [Google Scholar]
- Deepa, N.; Rakesh, S.; Sreenivasa, M.Y. Morphological, pathological and mycotoxicological variations among Fusarium verticillioides isolated from cereals. 3 Biotech 2018, 8, 105. [Google Scholar] [CrossRef]
- Joint FAO WHO Expert Committee on Food Additives, World Health Organization. Safety evaluation of certain food additives and contaminants. In Proceedings of the Seventy Fourth Meetings of the Joint FAO/WHO Expert Committee on Food Additives (JECFA), Rome, Italy, 14–23 June 2011. [Google Scholar]
- Miller, J.D. Factors That Affect the Occurrence of Fumonisin. Environ. Health Perspect. 2001, 109, 321. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasa, M.Y.; Dass, R.S.; Janardhana, G.R. Survey of post-harvest fungi associated with Sorghum grain produced in Karnatka (India). J. Plant Protect. Res. 2010, 50, 335–339. [Google Scholar] [CrossRef]
- Bacon, C.W.; Yates, I.E.; Hinton, D.M.; Meredith, F. Biological control of Fusarium moniliforme in maize. Environ. Health Persp. 2001, 109, 325–332. [Google Scholar]
- Bullerman, L.B.; Bianchini, A. Stability of mycotoxins during food processing. Int. J. Food Microbiol. 2007, 119, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, T.B.; Dickens, W.; Giesbrecht, F.G. Testing animal feedstuffs for mycotoxins: Sampling, subsampling, and analysis. In Mycotoxins and Animal Food; Smith, J.E., Henderson, R.S., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 153–164. [Google Scholar]
- Bankole, S.A.; Adebanjo, A. Mycotoxins in food in West Africa: Current situation and possibilities of controlling it. Afr. J. Biotechnol. 2003, 2, 254–263. [Google Scholar]
- Bennett, G.A.; Richard, J.L. Influence of processing on Fusarium mycotoxins in contaminated grains. Food Technol. 1996, 50, 235–238. [Google Scholar]
- Jackson Di, L.S.; DeVries, J.W.; Bullerman, L.B. Fumonisin in food. In Advances in Experimental Medicine and Biology; Plenum Press: New York, NY, USA, 1996; 392p. [Google Scholar]
- Misra, N.N.; Yadav, B.; Roopesh, M.S. Cold plasma for effective fungal and Mycotoxin control in foods: Mechanisms, inac-tivation effects, and applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, P.; Brunda, D.K.; Moses, J.; Anandharamakrishnan, C. Photolytic and photocatalytic detoxification of mycotoxins in foods. Food Control 2020, 123, 107748. [Google Scholar] [CrossRef]
- Lopez-Garcia, R.; Park, D.L. Effectiveness of post-harvest procedures in management of mycotoxin hazards. In Mycotoxins in Agriculture and Food Safety; Bhatnagar, D., Sinha, S., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1998; Volume 511, pp. 407–433. [Google Scholar]
- Sreenivasa, M.Y.; Maheshwar, P.K.; Sanjay, S.R.; Diwakar, B.T.; Naidu, K.A.; Janardhana, G.R. Effect of gamma irradiation on the incidence and fumonisin production by Fusarium species occurring on maize and sorghum grains. J. Agric. Technol. 2009, 5, 325–335. [Google Scholar]
- Deepthi, B.V.; Gnanaprakash, A.P.; Sreenivasa, M.Y. Effect of γ-irradiation on fumoniisn producing Fusarium associated with animal and poultry feed mixtures. 3 Biotech 2017, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in food and feed. Adv. Food Nutr. Res. 2019, 89, 297–345. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P.; Desjardins, A.E. Fumonisin in maize, can we reduce their occurrence? Plant Dis. 1997, 81, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Dantzer, W.R.; Hopmans, E.C.; Prisk, V.; Cunnick, J.E.; Murphy, P.A.; Hendrich, S. Reaction with Fructose Detoxifies Fumonisin B1while Stimulating Liver-Associated Natural Killer Cell Activity in Rats. J. Agric. Food Chem. 1997, 45, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Park, D.L.; Rua, S.M.; Mirocha, C.J.; Abd-Alla, E.S.A.M.; Weng, C.Y. Mutagenic potentials of fumonisin contaminated corn following ammonia decontamination procedure. Mycopathologia 1992, 117, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Sydenham, E.W.; Stockenstrom, S.; Thiel, P.G.; Shephard, G.S.; Koch, K.R.; Marasas, W.F.O. Potential of Alkaline Hydrolysis for the Removal of Fumonisins from Contaminated Corn. J. Agric. Food Chem. 1995, 43, 1198–1201. [Google Scholar] [CrossRef]
- Scott, P.M.; Lawrence, G.A. Analysis of Beer for Fumonisins. J. Food Prot. 1995, 58, 1379–1382. [Google Scholar] [CrossRef]
- Murphy, P.A.; Hendrich, S.; Hopmans, E.C.; Hauck, C.C.; Lu, Z.; Buseman, G.; Munkvold, G. Effect of Processing on Fumonisin Content of Corn. In Fumonisin in Food; Jackson, L.S., Devries, J.W., Bullerman, L.B., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 323–334. [Google Scholar]
- Hendrich, S.; Miller, K.A.; Wilson, T.M.; Murphy, P.A. Toxicity of Fusarium proliferatum-fermented nixtamalized corn-based diets fed to rats: Effect of nutritional status. J. Agric. Food Chem. 1993, 41, 1649–1654. [Google Scholar] [CrossRef] [Green Version]
- Dombrink-Kurtzman, M.A.; Javed, T.; Bennett, G.A.; Richard, J.L.; Cote, L.M.; Buck, W.B. Lymphocyte cytotoxicity and erythrocytic abnormalities induced in broiler chicks by fumonisins B1 and B2 and moniliformin from Fusarium proliferatum. Mycopathologia 1993, 124, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Park, D.L.; López-García, R.; Trujillo-Preciado, S.; Price, R.L. Reduction of Risks Associated with Fumonisin Contamination in Corn. In Fumonisin in Food; Jackson, L.S., Devries, J.W., Bullerman, L.B., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 335–344. [Google Scholar]
- Voss, K.A.; Bacon, C.W.; Meredith, F.I.; Norred, W.P. Comparative subchronic toxicity studies of nixtamalized and water-extracted Fusarium moniliforme culture material. Food Chem. Toxicol. 1996, 34, 623–632. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nut. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef]
- Visconti, A.; Boenke, A.; Solfrizzo, M.; Pascale, M.; Doko, M.B. European inter comparison study for the determination of the fumonisin content in two maize materials. Food Addit. Cont. 1996, 13, 909–927. [Google Scholar] [CrossRef]
- Cavaglieri, L.R.; Andres, L.; Ibanez, M.; Etcheverry, M.G. Rhizobacteria and their potential to control Fusarium verticillioides: Effect of maize bacterization and inoculum density. Antonie van Leeuwenhoek. 2005, 87, 179–187. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Migheli, Q.; Steinberg, C. Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt inducing Fusarium oxysporum. New Phytol. 2009, 184, 529–544. [Google Scholar] [CrossRef]
- Larkin, R.P.; Fravel, R. Efficacy of various fungal and bacterial biocontrol organisms for control of Fusarium wilt of tomato. Plant Dis. 1998, 82, 1022–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Cruz Cabral, L.; Pinto, V.F.; Patriarca, A. Application of plant derived compounds to control fungal spoilage and myco-toxin production in food. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal dis-eases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- United States Food and Drug Administration (US FDA). RAS Substances Evaluated by Select Committee on GRAS Substances (SCOGS): CFSAN/Office of Food Additive Safety. 2015. Available online: https://www.fda.gov/food/generally-recognized-safe-gras/gras-substances-scogs-database (accessed on 17 September 2021).
- Aly, S.E.; Abdel-Galil, M.M.; Abdel-Wahhab, M.A.; Abdel-Wahhab, M.A. Application of adsorbent agents technology in the removal of aflatoxin B1 and fumonisin B1 from malt extract. Food Chem. Toxicol. 2004, 42, 1825–1831. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.; Johnson, N.M.; Strey, A.; Taylor, J.F.; Marroquin-Cardona, A.; Mitchell, N.J. Calcium montmorillonite clay re-duces urinary biomarkers of fumonisin B1 exposure in rats and humans. Food Add. Cont. 2012, 29, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Howes, A.D.; Newman, K.E. Compositions and Methods for Removal of Mycotoxins from Animal Feed. U.S. Patent No. 6045834, 4 April 2000. [Google Scholar]
- He, J.; Zhou, T. Patented techniques for detoxification of mycotoxins in feeds and food matrices. Recent Pat. Food Nutr. Agric. 2010, 2, 96–104. [Google Scholar] [CrossRef]
- Deepa, N.; Sreenivasa, M.Y. Fusarium verticillioides, a globally important pathogen of Agriculture and Livestock: A review. J. Vet. Med. Res. 2017, 4, 1084. [Google Scholar]
- Huwig, A.K.; Freimund, S.; Kappeli, O. Mycotoxin detoxification of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Bacon, C.W.; Hinton, D.M. In planta reduction of maize seedling stalk lesions by the bacterial endophyte Bacillus mojavensis. Can. J. Microbiol. 2011, 57, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Cavaglieri, L.; Orlando, J.; Rodríguez, M.I.; Chulze, S.; Etcheverry, M. Biocontrol of Bacillus subtilis against Fusarium verticil-lioides in vitro and at the maize root level. Res. Microbiol. 2005, 156, 748–754. [Google Scholar] [CrossRef]
- Pereira, P.; Sabrina, G.I.; Agostini, E.; Etcheverry, M. Effects of maize inoculation with Fusarium verticillioides and with 2 bac-terial biocontrol agents on seedlings growth and antioxidative enzymatic activities. Appl. Soil Ecol. 2011, 51, 52–59. [Google Scholar] [CrossRef]
- Gelderblom, W.; Cawood, M.; Snyman, S.; Vleggaar, R.; Marasas, W. Structure-activity relationships of fumonisins in short-term carcinogenesis and cytotoxicity assays. Food Chem. Toxicol. 1993, 31, 407–414. [Google Scholar] [CrossRef]
- Duvick, J.; Rood, T.; Maddox, J.; Gilliam, J. Detoxification of Mycotoxins in Planta as a Strategy for Improving Grain Quality and Disease Resistance: Identification of Fumonisin-Degrading Microbes from Maize. Mol. Gen. Host.-Spec. Toxins Plant Dis. 1998, 13, 369–381. [Google Scholar] [CrossRef]
- Duvick, J.; Rood, T.; Wang, X. Fumonisin Detoxification Enzymes. U.S. Patent No. 5716820, 10 February 1998. [Google Scholar]
- Blackwell, B.A.; Gilliam, J.T.; Savard, M.E.; Miller, D.; Duvick, J.P. Oxidative deamination of hydrolysed fumonisin B1 (AP1) by cultures of Exophiala spinifera. Nat. Toxins 1999, 7, 31–38. [Google Scholar] [CrossRef]
- Cavaglieri, L.; Orlando, J.; Etcheverry, M. In vitro influence of bacterial mixtures on Fusarium verticillioides growth and fumonisin B1 production: Effect of seeds treatment on maize root colonization. Lett. Appl. Microbiol. 2005b, 41, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Nesci, A.; Castillo, C.; Etcheverry, M. Impact of bacterial biological control agents on fumonisin B1 content and Fusarium verticillioides infection of field-grown maize. Biol. Control 2010, 53, 258–266. [Google Scholar] [CrossRef]
- Al-Masri, S.A.; El- Safty, S.M.S.; Nada, S.A.; Amra, H.A. Saccharomyces cerevisiae and Probiotic Bacteria Potentially Inhibit Fumonisin B1 Production in Vitro and in vivo. J. Am. Sci. 2011, 7, 198–205. [Google Scholar]
- Cavaglieri, L.; Passone, A.; Etcheverry, M. Screening procedures for selecting rhizobacteria with biocontrol effects upon F. vericillioides growth and fumonisin B1 production. Res. Microbiol. 2004, 155, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Fareid, M.A. Biocontrol of Fusarium Moulds and Fumonisin B1 Production. World Rural Obs. 2011, 3, 58–61. [Google Scholar]
- Dalie, D.K.D.; Deschamps, A.M.; Atanasova-Penichon, V.; Richard-Forget, F. Potential of Pediococcus pentosaceus (L006) Isolated from Maize Leaf to Suppress Fumonisin-Producing Fungal Growth. J. Food Prot. 2010, 73, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Gergolet, D.G.; Pizzolitto, R.P.; Vazquex, C.; Usseglio, V.L.; Zunino, M.P.; Dambolena, J.S.; Zygadlo, J.A.; Merlo, C. Effects of the volatile organic compounds produced by Enterococcus spp. Strains isolated from maize grainsilod on Fusarium verticillioides growth and fumonisin B1 production. J. Stored Prod. Res. 2021, 93, 101825. [Google Scholar] [CrossRef]
- Calistru, C.; McLean, M.; Berjak, P. In vitro studies on the potential for biological control of Aspergillus flavus and Fusarium moniliforme by Trichoderma species. A study of the production of extracellular metabolites by Trichoderma species. Mycopathologia 1997, 137, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Yates, I.E.; Meredith, F.; Smart, W.; Bacon, C.W.; Jaworski, A.J. Trichoderma viride Suppresses Fumonisin B1 Production by Fusarium moniliforme. J. Food Prot. 1999, 62, 1326–1332. [Google Scholar] [CrossRef]
- Torres, A.; Ramirez, M.L.; Arroyo, M.; Chulze, S.; Magan, N. Potential use of antioxidants for control of growth and fumonisin production by Fusarium verticillioides and Fusarium proliferatum on whole maize grain. Int. J. Food Microbiol. 2002, 83, 319–324. [Google Scholar] [CrossRef]
- Coma, V.; Portes, E.; Gardrat, C.; Richard-Forget, F.; Castellan, A. In vitro inhibitory effect of tetrahydrocurcuminoids on Fusarium proliferatum growth and fumonisin B1 biosynthesis. Food Addit. Cont. 2011, 28, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Kapoor, I.P.S.; Singh, P.; de Heluani, C.S.; de Lampasona, M.P.; Catalan, C.A.N. Chemistry, antioxidant and an-timicrobial investigations on essential oil and oleoresins of Zingiber officinale. Food Chem. Toxicol. 2008, 46, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Menniti, A.; Gregori, R.; Neri, F. Activity of natural compounds on Fusarium verticillioides and fumonisin production in stored maize kernels. Int. J. Food Microbiol. 2010, 136, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xie, W.; Gong, Z. Antifungal activity of a chitin binding proteins from Ginkgo biloba. FEBS Lett. 2000, 478, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Samapundo, S.; De Meulenaer, B.; Osei-Nimoh, D.; Lamboni, L.Y.; Debevere, J.; Devlieghere, F. Can phenolic compounds be used for the protection of corn from fungal invasion and mycotoxin contamination during storage? Food Microbiol. 2007, 24, 465–473. [Google Scholar] [CrossRef]
- Beekrum, S.; Govinden, R.; Padayachee, T.; Odhav, B. Naturally occurring phenols: A detoxification strategy for fumonisin B1. Food Addit. Contam. 2003, 20, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.; Bainbridge, B.W.; Heala, J.B.; Perez, A.E.; Jimenez, D.R.M. A simple PCR based method for the detection of the chickpea-wilt pathogen Fusarium oxysporum f.sp. ciceris in artificial and natural soils. Eur. J. Plant Pathol. 1999, 105, 251–259. [Google Scholar] [CrossRef]
- Deepa, N.; Nayaka, C.S.; Udaya Shankar, A.C.; Krishna Kumar, V.; Niranjana, S.R.; Prakash, H.S.; Raghavendra, M.P. De-tection and management of seed-borne toxigenic Fusarium verticillioides by plant alkaloids. J. Mycol. Plant Pathol. 2012, 42, 161–166. [Google Scholar]
- Achimon, F.; Brito, V.D.; Pizzolitto, R.P.; Sanchez, A.R.; Gomez, E.A.; Zydadlor, J.A. Chemical composition and antifungal properties of commercial essential oils against the maize pythopathogenic fungus Fusarium verticillioides. Rev. Argent. Microbiol. 2021, in press. [Google Scholar]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.J.; Jubair, A.F.; Mohammadali, M.T. Antifungal activity of Tagetes erecta extract and Trichoderma harzianum on the pathogenic fungus Fusarium verticillioides. Plant Arch. 2020, 20, 185–188. [Google Scholar]
- Hinton, D.M.; Bacon, C.W. Enterobacter cloacae is an endophytic symbiont of corn. Mycopathologia 1995, 129, 117–125. [Google Scholar] [CrossRef]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in Biological Control: From Basic Research to Field Implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lee, K.M.; Cho, W.K.; Park, J.Y.; Kim, K.H. Differential contributions of RNA interference components in response to distinct Fusarium graminearum virus infectios. J. Virol. 2018, 92, 1756–1772. [Google Scholar] [CrossRef] [Green Version]
- Luongo, L.; Galli, M.; Corazza, L.; Meekes, E.; Haas, L.; Plas, L.C. Potential of fungal antagonists for bio-control of Fusarium spp. in wheat and maize through competition in crop debris. Biocontrol Sci. Technol. 2005, 15, 229–242. [Google Scholar] [CrossRef]
- Bluhm, B.H.; Kim, H.; Butchko, R.A.E.; Woloshuk, C.P. Involvement of ZFR1 of Fusarium verticillioides in kernel colonization and the regulation of FST1, a putative sugar transporter gene required for fumonisin biosynthesis on maize kernels. Mol. Plant Pathol. 2008, 9, 203–211. [Google Scholar] [CrossRef]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Etcheverry, M.; Torres, A.; Ramirez, M.; Chulze, S.; Magan, N. In vitro control of growth and fumonisin production by Fusarium verticillioides and F. proliferatum using antioxidants under different water availability and temperature regimes. J. Appl. Microbiol. 2002, 92, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Farnochi, M.; Torres, A.; Magan, N.; Chulze, S. Effect of antioxidants and competing mycoflora on Fusarium verticillioides and F. proliferatum populations and fumonisin production on maize grain. J. Stored Prod. Res. 2004, 41, 211–219. [Google Scholar] [CrossRef]
- Reynoso, M.M.; Torres, A.M.; Ramirez, M.L.; Rodríguez, M.I.; Chulze, S.N.; Magan, N. Efficacy of antioxidant mixtures on growth, fumonisin production and hydrolytic enzyme production by Fusarium verticillioides and F. proliferatum in vitro on maize-based media. Mycol. Res. 2002, 106, 1093–1099. [Google Scholar] [CrossRef]
- Roshan, A.B.; Mohana, D.C. Antimicrobial and antimycotoxigenic activities of (3s)-3,6,7-trihydroxy-α-tetralone isolated from endophytic Phoma moricola. Pharma Innov. J. 2021, 10, 852–860. [Google Scholar]
- Seepe, H.A.; Lodama, K.E.; Sutherland, R.; Nxumalo, W.; Amoo, S.O. In Vivo Antifungal Activity of South African Medicinal Plant Extracts against Fusarium pathogens and Their Phytotoxicity Evaluation. Plants 2020, 9, 1668. [Google Scholar] [CrossRef]
- Srichana, D.; Phumruang, A.; Chongkid, B. Inhibition Effect of Betel Leaf Extract on the Growth of Aspergillus flavus and Fusarium verticillioides Thammasat. Ind. J. Sci. Technol. 2009, 14, 74–77. [Google Scholar]
- Yooussef, M.M.; Pham, Q.; Achar, P.N.; Sreenivasa, M.Y. Antifungal activity of essential oils on Aspergillus parasiticus isolated from peanuts. J. Plant. Prot. Res. 2016, 56, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Hell, K.; Cardwell, K.; Setamou, M.; Poehling, H.-M. The influence of storage practices on aflatoxin contamination in maize in four agroecological zones of Benin, west Africa. J. Stored Prod. Res. 2000, 36, 365–382. [Google Scholar] [CrossRef]
- Dambolena, J.; Zygadlo, J.A.; Rubinstein, H. Antifumonisin activity of natural phenolic compounds: A structure–property–activity relationship study. Int. J. Food Microbiol. 2011, 145, 140–146. [Google Scholar] [CrossRef]
- Ferrari, S.; Vairo, D.; Ausubel, F.M.; Cervone, F.; De Lorenzo, G. Tandemly duplicated Arabidopsis genes that encode polyga-lacturonase-inhibiting proteins are regulated coordinately by different signal transduction pathways in response to fungal in-fection. Plant Cell Online 2003, 15, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Srichana, D.; Taengtip, R.; Kondo, S. Antimicrobial activity of Gynostemma pentaphyllum extracts against fungi producing aflatoxin and fumonisin and bacteria causing diarrheal disease. Southeast Asian J. Trop. Med. Public Health 2011, 42, 704. [Google Scholar] [PubMed]
- Sebayang, A.; Mirsam, H.; Pakki, S.; Azrai, M.; Muis, A. Control of Fusarium verticillioides on corn with a combination of B. subtilis TM3 formulation and botanical pesticides. Saudi J. Bio. Sci. 2021, in press. [Google Scholar] [CrossRef]
- Soliman, K.; Badeaa, R. Effect of oil extracted from some medicinal plants on different mycotoxigenic fungi. Food Chem. Toxicol. 2002, 40, 1669–1675. [Google Scholar] [CrossRef]
- López, A.G.; Theumer, M.G.; Zygadlo, J.A.; Rubinstein, H.R. Aromatic plants essential oils activity on F. verticillioides fumonsin B1 production in corn grains. Mycopathologia 2004, 158, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Egido, J.; Marin, S. Inhibitory effect of cinnamon, clove, lemongrass, oregano and pal-marose essential oils on growth and fumonisin B1 production by Fusarium proliferatum in maize grain. Int. J. Food Microbiol. 2003, 89, 145–154. [Google Scholar] [CrossRef]
- Nagendra Prasad, M.N.; Shankara Bhat, S.; Sreenivasa, M.Y. Antifungal activity of essential oils against Phomopsis azadirachtae-the causative agent of die-back disease of neem. J. Agric. Technol. 2010, 6, 27–133. [Google Scholar]
- Sreenivasa, M.Y.; Dass, R.S.; Charithraj, A.P.; Premila, N.A.; Janardhana, G.R. Assessment of the growth inhibiting effect of some plant essential growth inhibiting effect of some plant essential oils on different Fusarium species isolated from Sorghum and maize grains. J. Plant Dis. Protect. 2011, 118, 208–213. [Google Scholar] [CrossRef]
- López-Meneses, A.K.; Plascencia-Jatomea, M.; Lizardi-Mendoza, J.; Rosas-Burgos, E.C.; Luque-Alcaraz, A.G.; Cortez-Rocha, M.O. Anti-fungal and antimycotoxigenic activity of essential oils from Eucalyptus globulus, Thymus capitatus and Schinus molle. Food Sci. Technol. 2015, 35, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Houicher, A.; Hamdi, M.; Hechachna, H.; Özogul, F. Chemical composition and antifungal activity of Anacyclus valentinus essential oil from Algeria. Food Biosci. 2018, 25, 28–31. [Google Scholar] [CrossRef]
- Zabka, M.; Pavela, R.; Slezakova, L. Antifungal effect of Pimenta dioica essential oil against dangerous pathogenic and tox-inogenic fungi. Ind. Crops Prod. 2009, 30, 250–253. [Google Scholar] [CrossRef]
- Xing, F.; Hua, H.; Selvaraj, J.N.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Growth inhibition and morphological alterations of Fusarium verticillioides by cinnamon oil and cinnamaldehyde. Food Control 2014, 46, 343–350. [Google Scholar] [CrossRef]
- Sharifzadeh, A.; Javan, A.J.; Shokri, H.; Abbaszadeh, S.; Keykhosravy, K. Evaluation of antioxidant and antifungal proper-ties of the traditional plants against foodborne fungal pathogens. J. Mycol. Med. 2016, 26, e11–e17. [Google Scholar] [CrossRef]
- Davari, M.; Ezazi, R. Chemical composition and antifungal activity of the essential oil of Zhumeria majdae, Heracleum persicum and Eucalyptus sp. against some important phytopathogenic fungi. J. Mycol. Med. 2017, 27, 463–468. [Google Scholar] [CrossRef]
- Dambolena, J.; Zunino, M.; López, A.G.; Rubinstein, H.; Zygadlo, J.A.; Mwangi, J.W.; Thoithi, G.N.; Kibwage, I.; Mwalukumbi, J.M.; Kariuki, S.T. Essential oils composition of Ocimum basilicum L. and Ocimum gratissimum L. from Kenya and their inhibitory effects on growth and fumonisin production by Fusarium verticillioides. Innov. Food Sci. Emerg. Technol. 2010, 11, 410–414. [Google Scholar] [CrossRef]
- Ingok, A.M.; Devoglu, D.; Dikmetas, N.; Karbancioglu-Guler, F.; Capanoglu, E. Antibacterial, antifungal, antimycotoxigenic, antioxidant activities of essential oils: An updated review. Molecules 2020, 25, 4711. [Google Scholar] [CrossRef]
- Duvick, J. Prospects for reducing fumonisin contamination of maize through genetic modification. Environ. Health Persp. 2001, 109, 337–342. [Google Scholar]
- Cleveland, T.E.; Dowd, P.F.; Desjardins, A.E.; Bhatnagar, D.; Cotty, P.J. United States Department of Agriculture? Agricultural Research Service research on pre-harvest prevention of mycotoxins and mycotoxigenic fungi in US crops. Pest. Manag. Sci. 2003, 59, 629–642. [Google Scholar] [CrossRef]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional genomic analysis of constitutive and inducible defense responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance. BMC Genom. 2014, 15, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 Maize Genome: Complexity, Diversity, and Dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandra, L.; Luca, P.; Adriano, M. Differential gene expression in kernels and skills of maize lines with contrasting levels of ear rot resistance after Fusarium verticillioides infection. J. Plant Physiol. 2010, 167, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Maschietto, V.; Marocco, A.; Malachova, A.; Lanubile, A. Resistance to Fusarium verticillioides and fumonisin accumulation in maize inbred lines involves an earlier and enhanced expression of lipoxygenase (LOX) genes. J. Plant Physiol. 2015, 188, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Butròn, A.; Santiago, R.; Mansilla, P.; Pintos-Varela, C.; Ordás, A.; Malvar, R.A. Maize (Zea mays L.) Genetic factors for pre-venting fumonisin contamination. J. Agric. Food Chem. 2006, 54, 6113–6117. [Google Scholar] [CrossRef]
- Dowd, P.F. Biotic and abiotic factors limiting efficacy of Bt corn in indirectly reducing mycotoxin levels in commercial fields. J. Econ. Èntomol. 2001, 94, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.G.; Campbell, K.W.; Pilcher, C.D.; DeGooyer, T.A.; Robinson, A.E.; McMillen, B.L.; Spangler, S.M.; Riordan, S.G.; Rice, L.G.; Richard, J.L. Lower Fumonisin Mycotoxin Levels in the Grain of Bt Corn Grown in the United States in 2000−2002. J. Agric. Food Chem. 2004, 52, 1390–1397. [Google Scholar] [CrossRef]
- International Programme on Chemical Safety (IPCS). Environmental Health Criteria 217: Microbial Pest Control Agent Bacillus Thuringiensis; World Health Organisation (WHO): Geneva, Switzerland, 1999. [Google Scholar]
- Wu, F. Mycotoxin Reduction in Bt Corn: Potential Economic, Health, and Regulatory Impacts. Transgenic Res. 2006, 15, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Daou, R.; Joubrane, K.; Maroun, R.G.; Khabbaz, L.R.; Ismail, A.; El Khoury, A. Mycotoxins: Factors influencing production and control strategies. AIMS Agric. Food 2021, 6, 416–447. [Google Scholar] [CrossRef]


| Serial Number | Crop | Disease | Toxins |
|---|---|---|---|
| 1. | Corn/Maize (Zea mays) | Fusarium ear rot, stalk rot, kernel rot, root rot, seed rot, seedling blight, seedling root rot | FB1, FB2, and FB3 |
| 2. | Cultivated wild rice (Zizania palustris) | Scab | FB1, FB2 |
| 3. | Oats (Avens sativa) | Fusarium foot rot, snow mold, seedling blight, head blight | FB1, FB2 |
| 4. | Pearl millet (Pennisetum glauccum) | Top rot | FB2, FB1 |
| 5. | Rice (Oryza sativa) | Seedling blight, water mold, root rot, pecky rice (kernel spotting) | FB1, FB2, FB3 |
| 6. | Sorghum (Sorghum bicolor) | Damping off and seed rot, Fusarium wilt head blight, root and stalk rot, twisted top, seedling blight, seed rot | FB2, FB1 |
| 7. | Sugarcane (Saccharum spp.) | Fusarium stem rot, pokkah baeng, wilt; | FB1, FB2 |
| 8. | Wheat (Triticum spp.) | Black point (kernel smudge), head blight (scab), root, crown, and foot rot, pink snow mold; | FB1, FB2 |
| Serial Number | Plant Parts Treated with Biocontrol Agents | Test Organisms | Targeted Feature | Percent Inhibition |
|---|---|---|---|---|
| 1. | Maize plant | Bacillus subtilis | Colonization | 28–78% |
| 2. | Maize seedling stalk | Bacillus mojavensis | Colonization | 24–58% |
| 3. | Seed | Bacillus amyloliquifaciens | Fungal growth | >82% |
| 4. | Seed | Microbacterium oleovarans | Maize infection | |
| 5. | Seed | Enterobacter hormacchei | Fungal growth | |
| 6. | Maize roots | Enterobacter cloacae | Colonization in roots | <50% |
| 7. | Maize stalk | Clonostachys rosae | Colonization | 50% |
| 8. | Rhizoplane and endorhizosphere region | Arthrobacter globiformis | Root colonization | 69–80% |
| 9. | Azotobacter armeniacus | Root colonization | 56–75% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
N, D.; Achar, P.N.; Sreenivasa, M.Y. Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review. J. Fungi 2021, 7, 776. https://doi.org/10.3390/jof7090776
N D, Achar PN, Sreenivasa MY. Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review. Journal of Fungi. 2021; 7(9):776. https://doi.org/10.3390/jof7090776
Chicago/Turabian StyleN, Deepa, Premila N. Achar, and Marikunte Y. Sreenivasa. 2021. "Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review" Journal of Fungi 7, no. 9: 776. https://doi.org/10.3390/jof7090776
APA StyleN, D., Achar, P. N., & Sreenivasa, M. Y. (2021). Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review. Journal of Fungi, 7(9), 776. https://doi.org/10.3390/jof7090776