
Screening and Identification of Indigenous Entomopathogenic Fungal Isolates from Agricultural Farmland Soils in Nile Delta, Egypt

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
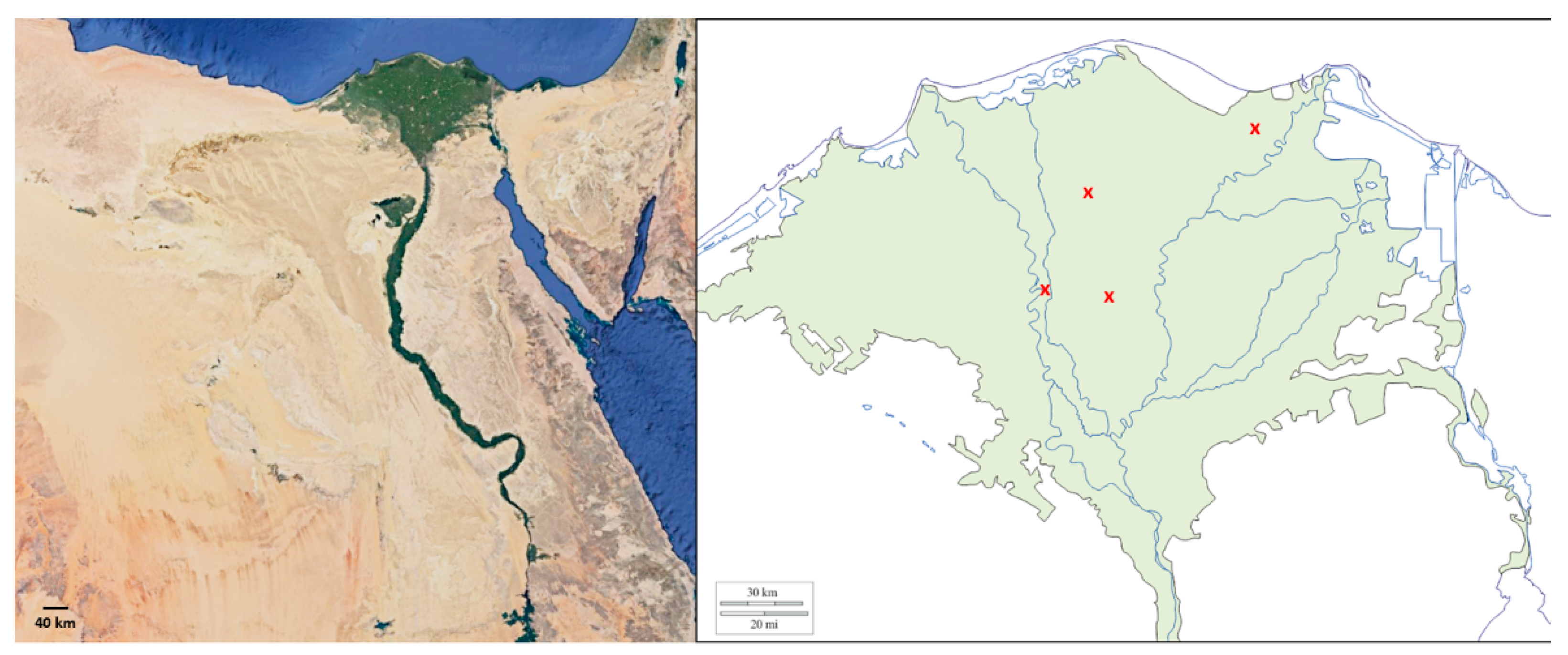
2.1. Soil Sampling
2.2. Entrapment of Entomopathogenic Fungi by Insect Baiting
2.3. Pathogenicity Screening
2.4. Molecular Identification and Phylogenetic Analysis
2.5. Second Round of Bioassays
2.6. Thermotolerance and Conidial Germination Assay
2.7. Statistical Analysis
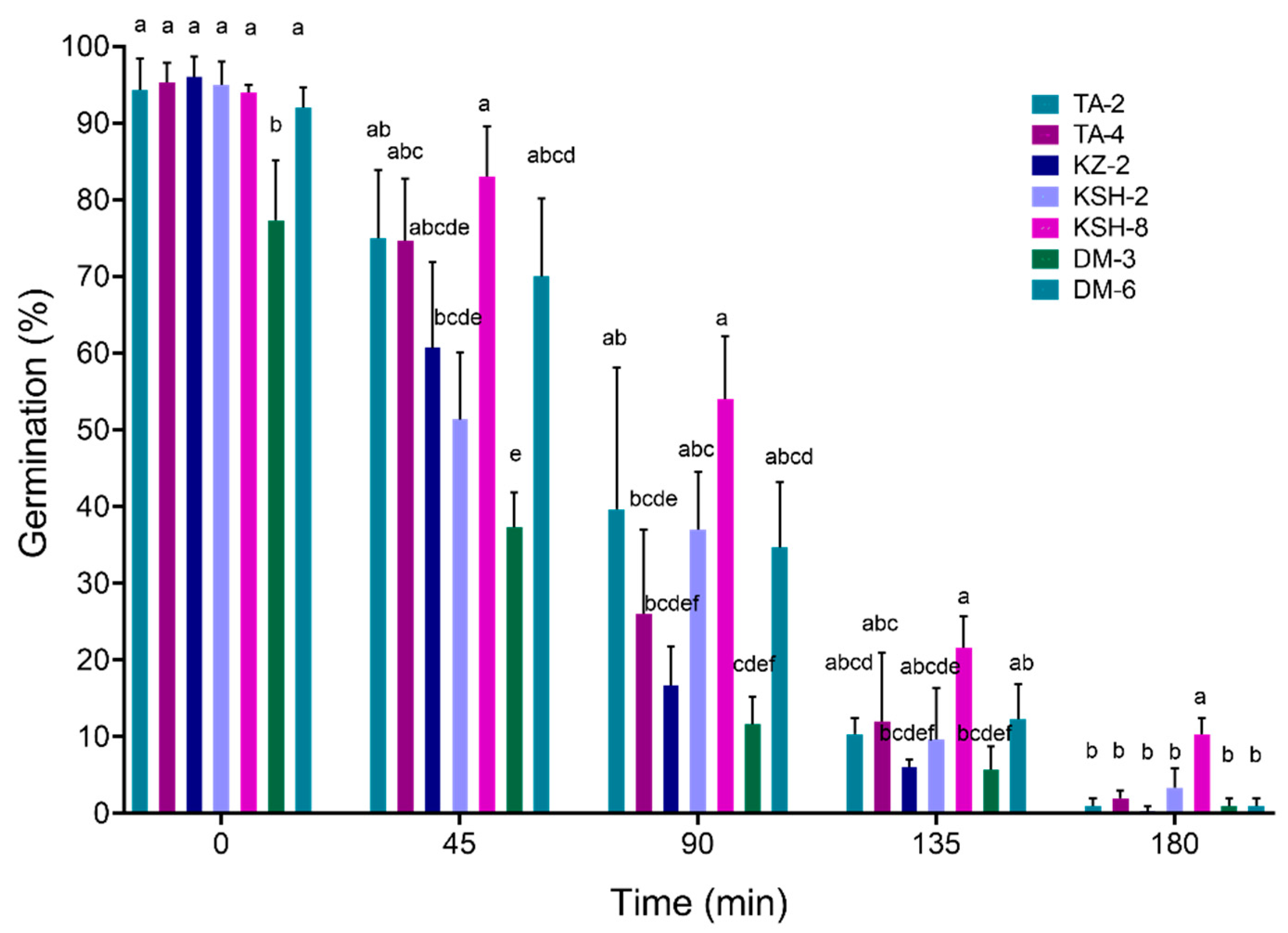
3. Results
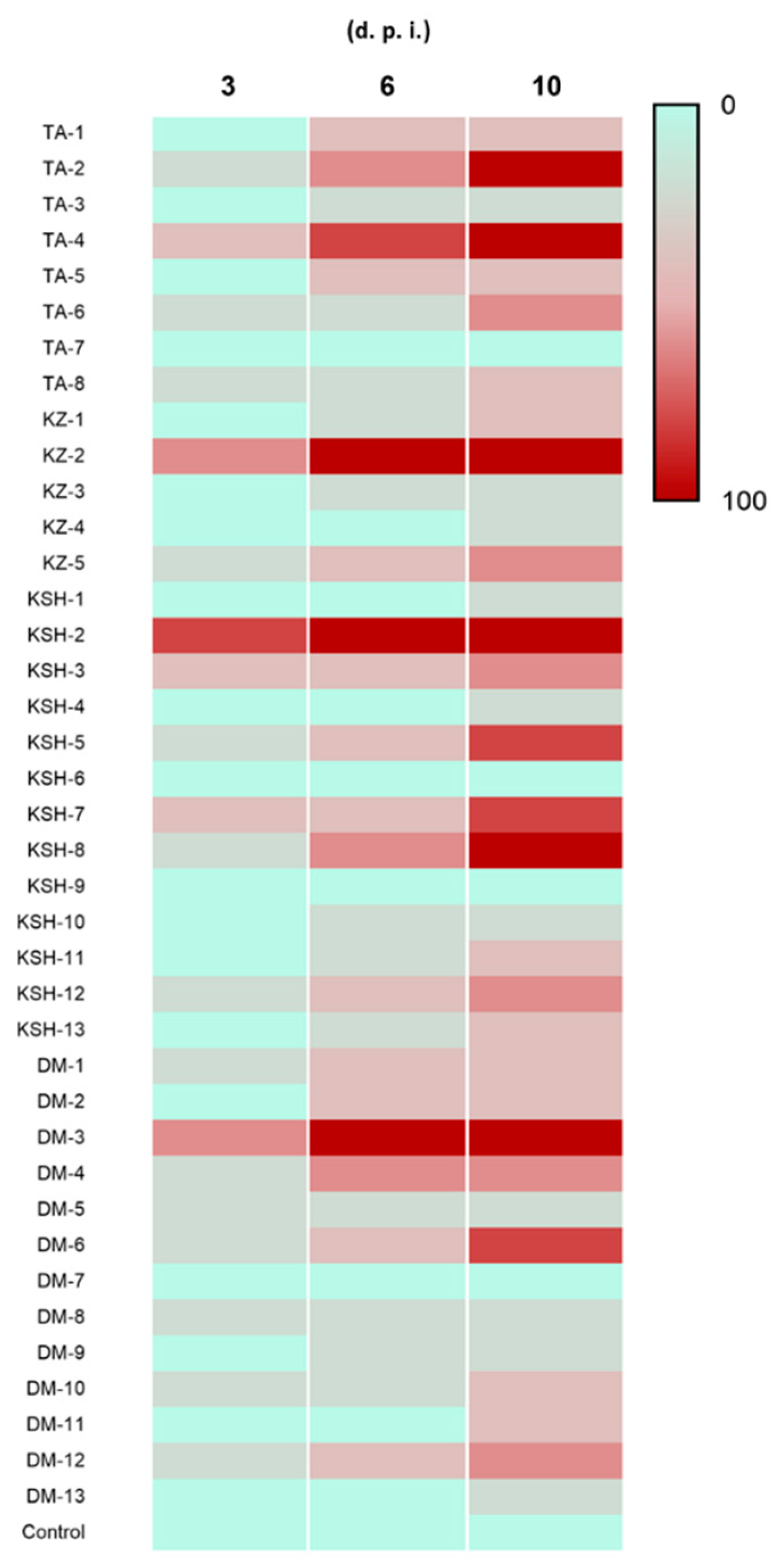
3.1. Isolation and First Pathogenicity Screening of Insect Associated Fungal Isolates
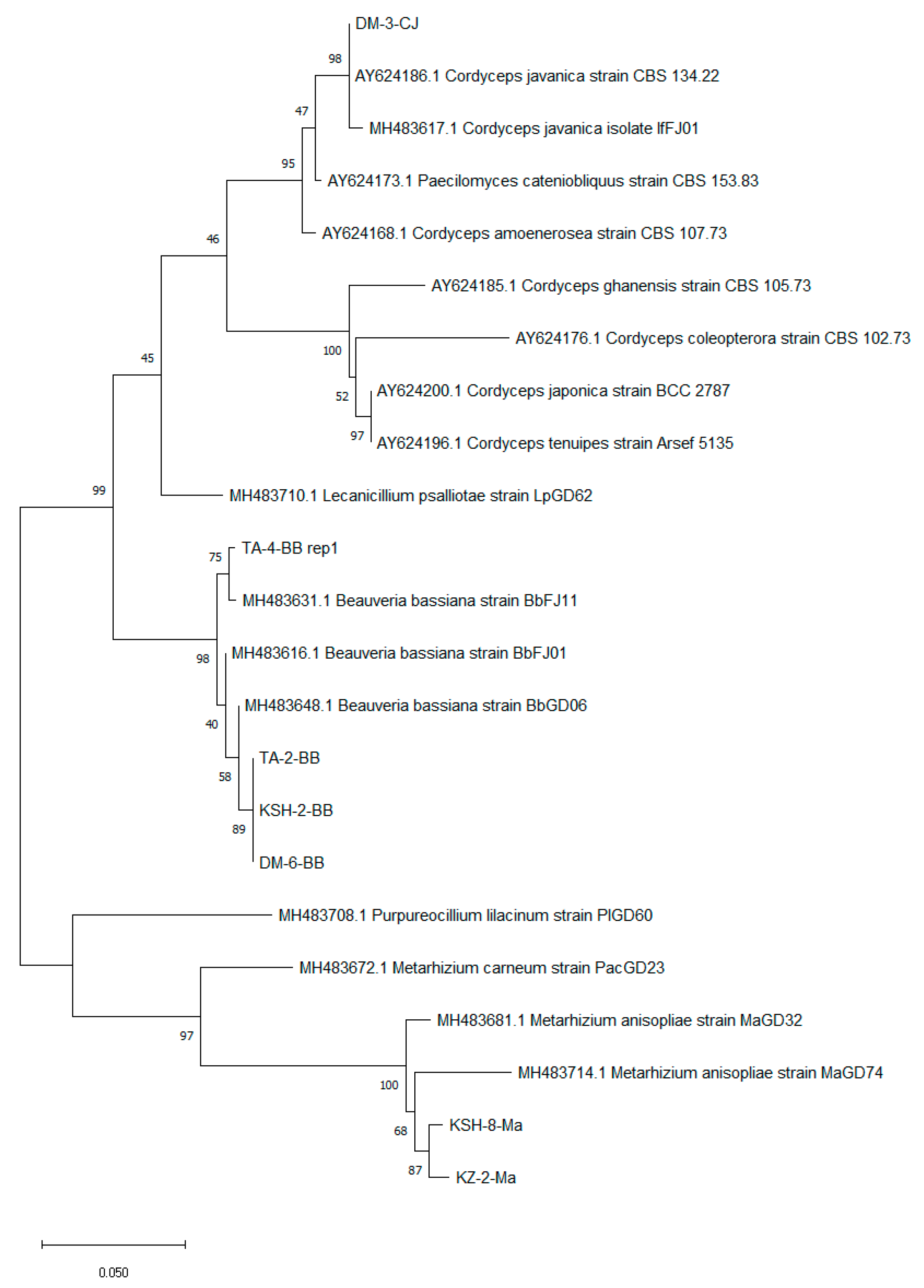
3.2. Molecular Identification and Phylogenetic Analysis
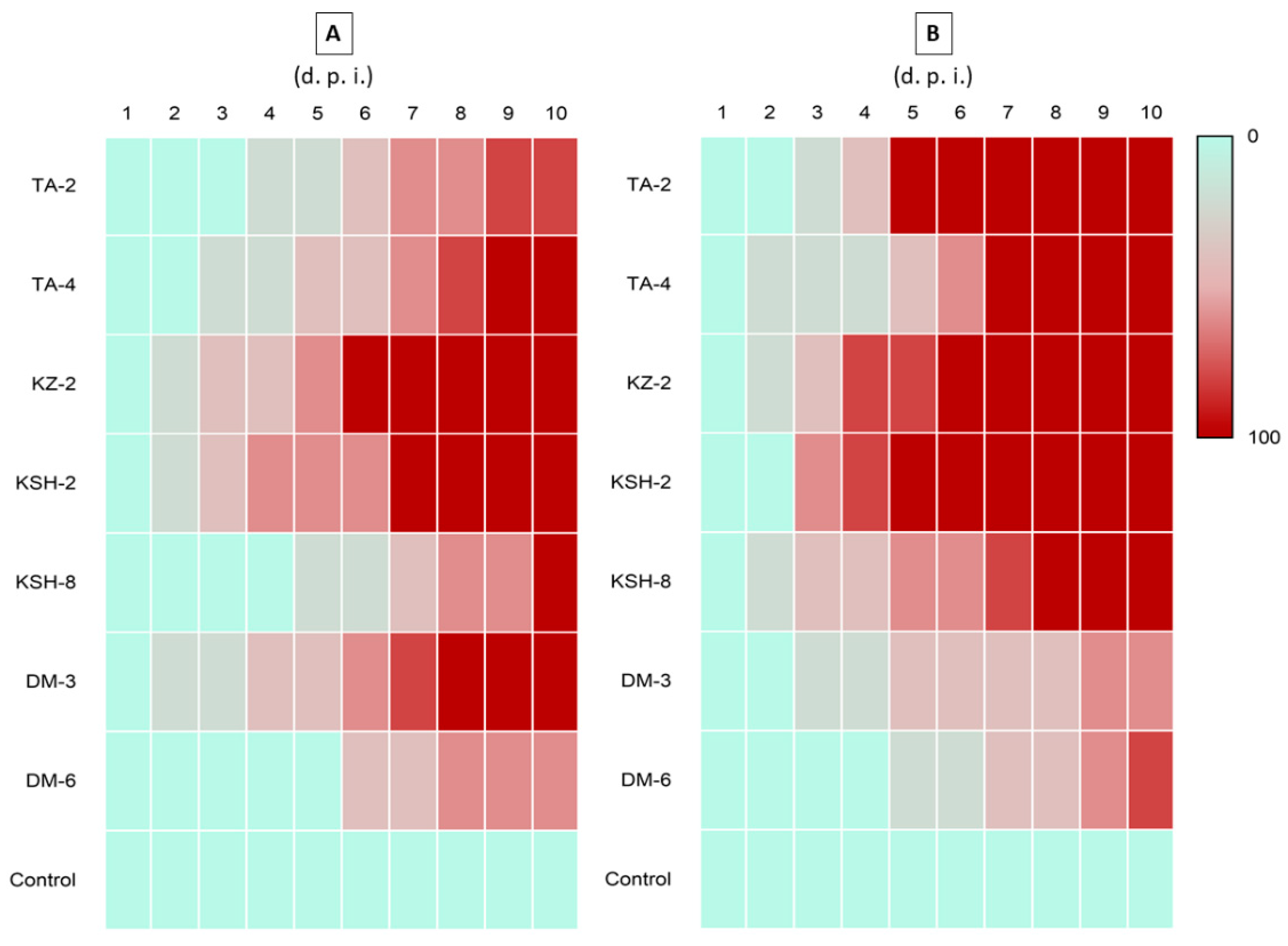
3.3. Second Round of Pathogenicity against S. litura and T. molitor
3.4. Thermotolerance Assay
4. Discussion
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect. Pests 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Crop Losses and the Economic Impact of Insect Pests on Brazilian Agriculture. Crop. Prot. 2014, 56, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Kooner, R.; Arora, R. Insect Pests and Crop Losses. In Breeding Insect Resistant Crops for Sustainable Agriculture; Arora, R., Sandhu, S., Eds.; Springer: Singapore, 2017; pp. 45–66. [Google Scholar]
- DiBartolomeis, M.; Kegley, S.; Mineau, P.; Radford, R.; Klein, K. An Assessment of Acute Insecticide Toxicity Loading (AITL) of Chemical Pesticides Used on Agricultural Land in the United States. PLoS ONE 2019, 14, e0220029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ara, Z.G.; Haque, A.R. A Comprehensive Review on Synthetic Insecticides: Toxicity to Pollinators, Associated Risk to Food Security, and Management Approaches. J. Biosyst. Eng. 2021, 46, 254–272. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic Entomopathogenic Fungi: A Valuable Biological Control Tool against Plant Pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-Fungal-Interactions: A Detailed Review on Entomopathogenic Fungi Pathogenicity to Combat Insect Pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic Fungi: Unconventional Applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Coombes, C.A.; Hill, M.P.; Moore, S.D.; Dames, J.F. Entomopathogenic Fungi as Control Agents of Thaumatotibia leucotreta in Citrus Orchards: Field Efficacy and Persistence. BioControl 2016, 61, 729–739. [Google Scholar] [CrossRef]
- Konopická, J.; Bohatá, A.; NERMUT, J.; Jozová, E.; Mráček, Z.; Palevsky, E.; Zemek, R. Efficacy of Soil Isolates of Entomopathogenic Fungi against the Bulb Mite, Rhizoglyphus robini (Acari: Acaridae). Syst. Appl. Acarol. 2021, 26, 1149–1167. [Google Scholar] [CrossRef]
- Wakil, W.; Kavallieratos, N.G.; Ghazanfar, M.U.; Usman, M.; Habib, A.; El-Shafie, H.A.F. Efficacy of Different Entomopathogenic Fungal Isolates against Four Key Stored-Grain Beetle Species. J. Stored Prod. Res. 2021, 93, 101845. [Google Scholar] [CrossRef]
- Sevim, A.; Demir, I.; Höfte, M.; Humber, R.A.; Demirbag, Z. Isolation and Characterization of Entomopathogenic Fungi from Hazelnut-Growing Region of Turkey. BioControl 2010, 55, 279–297. [Google Scholar] [CrossRef]
- Niu, X.; Xie, W.; Zhang, J.; Hu, Q. Biodiversity of Entomopathogenic Fungi in the Soils of South China. Microorganisms 2019, 7, 311. [Google Scholar] [CrossRef] [Green Version]
- Cherry, A.J.; Abalo, P.; Hell, K. A Laboratory Assessment of the Potential of Different Strains of the Entomopathogenic Fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to Control Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in Stored Cowpea. J. Stored Prod. Res. 2005, 41, 295–309. [Google Scholar] [CrossRef]
- El Gohary, R. Agriculture, Industry, and Wastewater in the Nile Delta. Int. J. Sci. Res. Agric. Sci. 2015, 22, 159–172. [Google Scholar]
- Radwan, T.M.; Blackburn, G.A.; Whyatt, J.D.; Atkinson, P.M. Dramatic Loss of Agricultural Land Due to Urban Expansion Threatens Food Security in the Nile Delta, Egypt. Remote Sens. 2019, 11, 332. [Google Scholar] [CrossRef] [Green Version]
- Fishar, M.R. Nile Delta (Egypt). In The Wetland Book: II: Distribution, Description, and Conservation; Finlayson, C.M., Milton, G.R., Prentice, R.C., Davidson, N.C., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 1251–1260. [Google Scholar] [CrossRef]
- Köberl, M.; Wagner, P.; Müller, H.; Matzer, R.; Unterfrauner, H.; Cernava, T.; Berg, G. Unraveling the Complexity of Soil Microbiomes in a Large-Scale Study Subjected to Different Agricultural Management in Styria. Front. Microbiol. 2020, 11, 1052. [Google Scholar] [CrossRef]
- Ekesi, S.; Maniania, N.K.; Lux, S.A. Effect of Soil Temperature and Moisture on Survival and Infectivity of Metarhizium anisopliae to Four Tephritid Fruit Fly Puparia. J. Invertebr. Pathol. 2003, 83, 157–167. [Google Scholar] [CrossRef]
- Kessler, P.; Enkerl, J.; Schweize, C.; Keller, S. Survival of Beauveria brongniartii in the Soil after Application as a Biocontrol Agent against the European Cockchafer Melolontha melolontha. BioControl 2004, 49, 563–581. [Google Scholar] [CrossRef]
- El-Husseini, M.M.; El-Heneidy, A.H.; Awadallah, K.T. Natural Enemies Associated with Some Economic Pests in Egyptian Agro-Ecosystems. Egypt. J. Biol. Pest Control 2018, 28, 78. [Google Scholar] [CrossRef]
- Hussein, K.A.; Abdel-Rahman, M.A.A.; Abdel-Mallek, A.Y.; El-Maraghy, S.S.; Joo, J.H. Climatic Factors Interference with the Occurrence of Beauveria bassiana and Metarhizium anisopliae in Cultivated Soil. Afr. J. Biotechnol. 2010, 9, 7674–7682. [Google Scholar] [CrossRef]
- Taylor, W.W.; Radcliffe, F.; van Peenen, P.F.D. The Isolation of Pathogenic Fungi from the Soils of Egypt, the Sudan, and Ethiopia. Sabouraudia J. Med. Vet. Mycol. 1964, 3, 235–238. [Google Scholar] [CrossRef]
- Sahar, S.A.; Entesar, N.S.H.; Magdy, A.A.; Afaf, A.A.; Manal, E.A.E. Isolation of Entomopathogenic Fungi and Efficacy as a Biological Control Agent on Red Spider Mite Tetranychus urticae (Acari: Tetranychidae). Egypt. J. Plant Prot. Res. Inst. 2019, 2, 759–769. [Google Scholar]
- Atia, M.; Farid, A.; Zaki, M.M. The Isolation of Pathogenic Fungi and Actinomycetes from Soil in Egypt. Sabouraudia J. Med. Vet. Mycol. 1981, 19, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, A.H.; Moustafa, A.F. A Survey of Egyptian Soil Fungi with Special Reference to Aspergillus, Penicillium and Penicillium-Related Genera. Trans. Br. Mycol. Soc. 1970, 54, 35–44. [Google Scholar] [CrossRef]
- Zimmermann, G. The ‘Galleria Bait Method’ for Detection of Entomopathogenic Fungi in Soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Firacative, C.; Khan, A.; Duan, S.; Ferreira-Paim, K.; Leemon, D.; Meyer, W. Rearing and Maintenance of Galleria mellonella and Its Application to Study Fungal Virulence. J. Fungi 2020, 6, 130. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, A.; Wang, M.; Zhang, X.; Wang, C.; Qiu, Z.; Wang, W.; Wang, H.; Liu, J. Meta-Analysis and Evaluation by Insect-Mediated Baiting Reveal Different Patterns of Hypocrealean Entomopathogenic Fungi in the Soils from Two Regions of China. Front. Microbiol. 2020, 11, 1133. [Google Scholar] [CrossRef]
- Keyser, C.A.; De Fine Licht, H.H.; Steinwender, B.M.; Meyling, N.V. Diversity within the Entomopathogenic Fungal Species Metarhizium Flavoviride Associated with Agricultural Crops in Denmark. BMC Microbiol. 2015, 15, 249. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.W.; Hyde, K.D.; Ho, W.W.H. Single Spore Isolation of Fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- David, W.A.L.; Ellaby, S.; Taylor, G. Rearing Spodoptera exempta on Semi-Synthetic Diets and on Growing Maize. Entomol. Exp. Appl. 1975, 18, 226–237. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Manoch, L.; Samson, R.A. On the Relationships of Paecilomyces Sect. Isarioidea Species. Mycol. Res. 2005, 109, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Alves, S.B.; Roberts, D.W.; Fan, M.; Delalibera, I.; Tang, J.; Lopes, R.B.; Faria, M.; Rangel, D.E.N. Biological Control of Insects in Brazil and China: History, Current Programs and Reasons for Their Successes Using Entomopathogenic Fungi. Biocontrol Sci. Technol. 2010, 20, 117–136. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.A.; Pell, J.K. Entomopathogenic Fungi as Biological Control Agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, B.; Poornesha, B.; Shylesha, A.N. Effect of Entomopathogenic Fungi against Invasive Pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in Maize. Egypt. J. Biol. Pest Control 2020, 30, 100. [Google Scholar] [CrossRef]
- El Kichaoui, A.Y.; Bara’a, A.; El Hindi, M. Isolation, Molecular Identification and under Lab Evaluation of the Entomopathogenic Fungi M. Anisopliae and B. Bassiana against the Red Palm Weevil R. ferrugineus in Gaza Strip. Adv. Microbiol. 2017, 7, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.C.; Wu, S.S.; Liu, Y.C.; Yang, Y.H.; Tsai, Y.F.; Li, Y.H.; Tseng, C.T.; Tang, L.C.; Nai, Y.S. Construction and Selection of an Entomopathogenic Fungal Library from Soil Samples for Controlling Spodoptera litura. Front. Sustain. Food Syst. 2021, 5, 15. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, M.R.; Kim, S.; Lee, S.J.; Park, S.E.; Nai, Y.S.; Lee, G.S.; Shin, T.Y.; Kim, J.S. Tenebrio molitor-Mediated Entomopathogenic Fungal Library Construction for Pest Management. J. Asia. Pac. Entomol. 2018, 21, 196–204. [Google Scholar] [CrossRef]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha Arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef]
- Bidochka, M.J.; Kasperski, J.E.; Wild, G.A.M. Occurrence of the Entomopathogenic Fungi Metarhizium anisopliae and Beauveria bassiana in Soils from Temperate and Near-Northern Habitats. Can. J. Bot. 1998, 76, 1198–1204. [Google Scholar] [CrossRef]
- McGuire, A.V.; Northfield, T.D. Tropical Occurrence and Agricultural Importance of Beauveria bassiana and Metarhizium anisopliae. Front. Sustain. Food Syst. 2020, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atef, M.M.; Ebrahim, S.A.A.; Eladawy, A.M.M. Biodiversity and Occurrence of Entomopathogenic Agents in Suez Canal Area-Egypt. Egypt. J. Agric. Res. 2018, 96, 445–464. [Google Scholar]
- Ou, D.; Zhang, L.H.; Guo, C.F.; Chen, X.S.; Ali, S.; Qiu, B.L. Identification of a New Cordyceps javanica Fungus Isolate and Its Toxicity Evaluation against Asian Citrus Psyllid. Microbiologyopen 2019, 8, e00760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, P.B.; Duren, E.B.; Qureshi, J.A.; Adair, R.C.; Adair, M.M.; Cave, R.D. Field Efficacy of Cordyceps javanica, White Oil and Spinetoram for the Management of the Asian Citrus Psyllid, Diaphorina citri. Insects 2021, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Baja, F.; Poitevin, C.G.; Araujo, E.S.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Infection of Beauveria bassiana and Cordyceps javanica on Different Immature Stages of Duponchelia fovealis Zeller (Lepidoptera: Crambidae). Crop. Prot. 2020, 138, 105347. [Google Scholar] [CrossRef]
- Kang, B.R.; Han, J.H.; Kim, J.J.; Kim, Y.C. Dual Biocontrol Potential of the Entomopathogenic Fungus, Isaria javanica, for Both Aphids and Plant Fungal Pathogens. Mycobiology 2018, 46, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Xie, W.; Cai, W.; Thaochan, N.; Hu, Q. Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential. J. Fungi 2021, 7, 984. [Google Scholar] [CrossRef]
- Kassem, H.S.; Bello, A.R.S.; Alotaibi, B.M.; Aldosri, F.O.; Straquadine, G.S. Climate Change Adaptation in the Delta Nile Region of Egypt: Implications for Agricultural Extension. Sustainability 2019, 11, 685. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.H.; Shin, T.Y.; Lee, J.Y.; Choi, C.J.; Woo, S.D. Screening and Evaluation of Acaropathogenic Fungi against the Bulb Mite Rhizoglyphus Robini. J. Asia. Pac. Entomol. 2021, 24, 991–996. [Google Scholar] [CrossRef]
- da Cruz Couceiro, J.; Fatoretto, M.B.; Demétrio, C.G.B.; Meyling, N.V.; Delalibera, Í. UV-B Radiation Tolerance and Temperature-Dependent Activity Within the Entomopathogenic Fungal Genus Metarhizium in Brazil. Front. Fungal Biol. 2021, 2, 6. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfiky, A. Screening and Identification of Indigenous Entomopathogenic Fungal Isolates from Agricultural Farmland Soils in Nile Delta, Egypt. J. Fungi 2022, 8, 54. https://doi.org/10.3390/jof8010054
Alfiky A. Screening and Identification of Indigenous Entomopathogenic Fungal Isolates from Agricultural Farmland Soils in Nile Delta, Egypt. Journal of Fungi. 2022; 8(1):54. https://doi.org/10.3390/jof8010054
Chicago/Turabian StyleAlfiky, Alsayed. 2022. "Screening and Identification of Indigenous Entomopathogenic Fungal Isolates from Agricultural Farmland Soils in Nile Delta, Egypt" Journal of Fungi 8, no. 1: 54. https://doi.org/10.3390/jof8010054
APA StyleAlfiky, A. (2022). Screening and Identification of Indigenous Entomopathogenic Fungal Isolates from Agricultural Farmland Soils in Nile Delta, Egypt. Journal of Fungi, 8(1), 54. https://doi.org/10.3390/jof8010054