
Impact of Soil Chemical Properties on the Growth Promotion Ability of Trichoderma ghanense, T. tomentosum and Their Complex on Rye in Different Land-Use Systems

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungi
2.2. Identification of Fungi
2.3. Trichoderma spp. Physiological Characteristics
2.4. Soil Sampling and Analysis
2.5. The Estimation of CM-Cellulase Activity of Soil
2.6. Design of Experiment
2.7. Statistical Analysis
3. Results
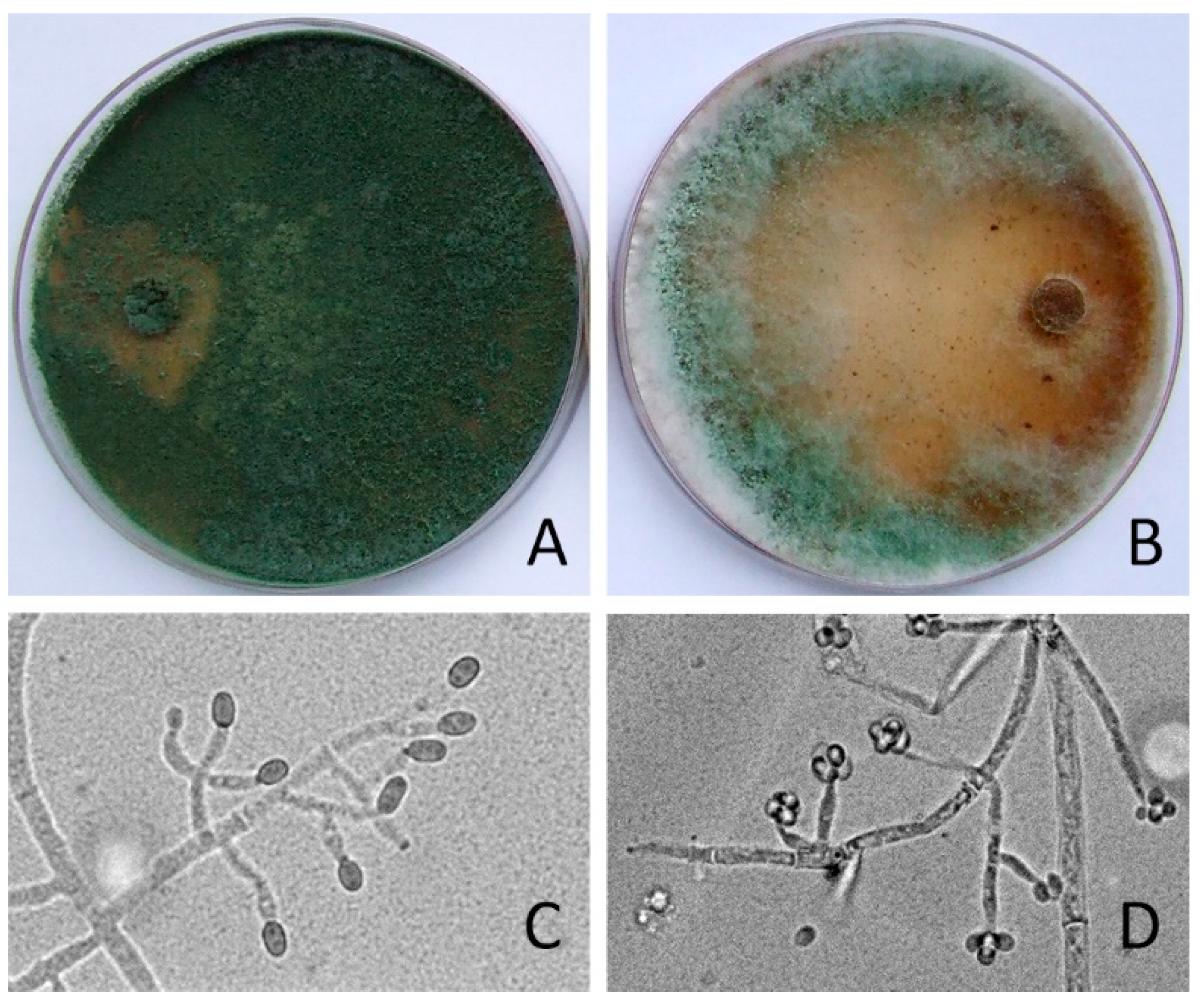
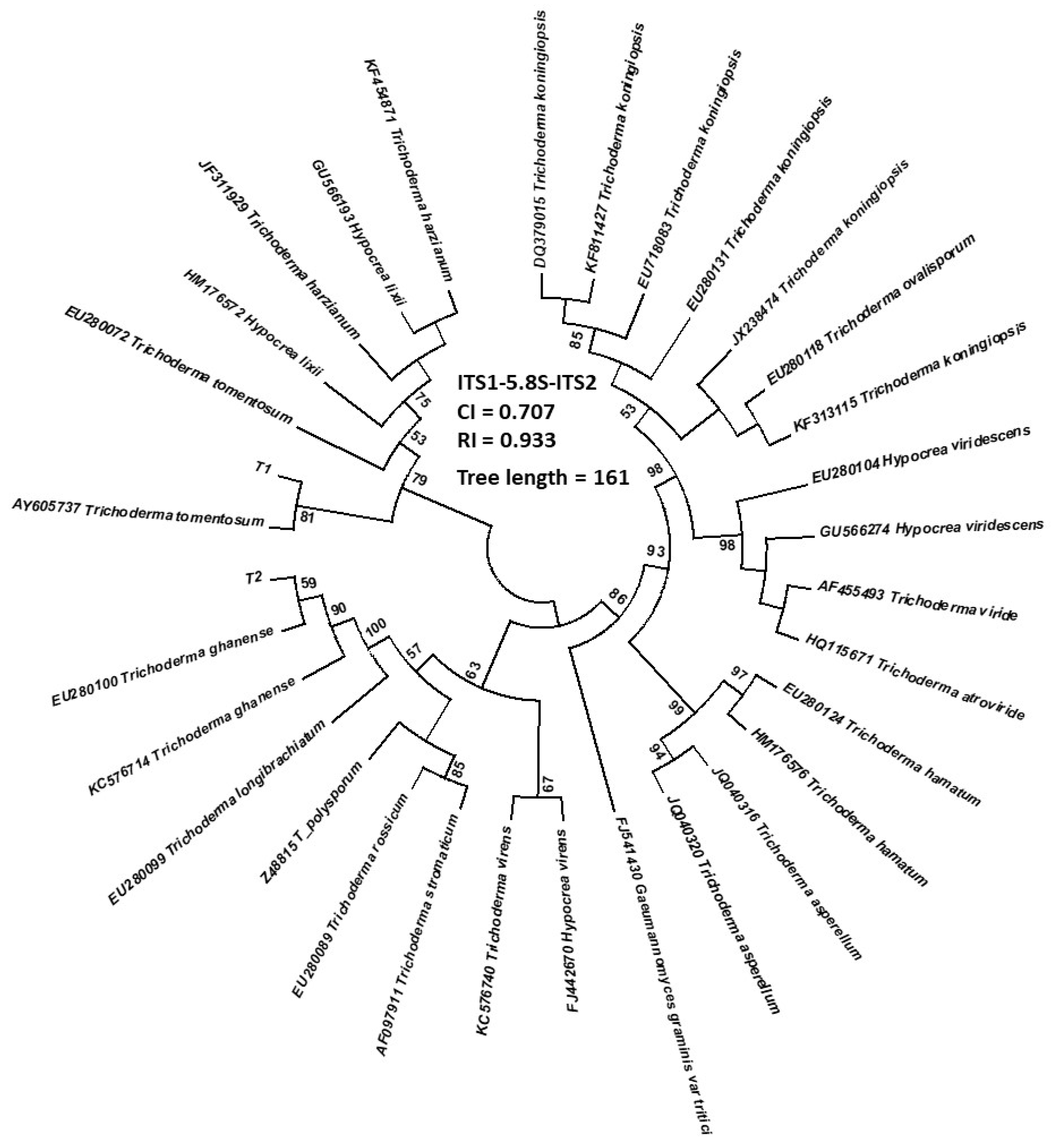
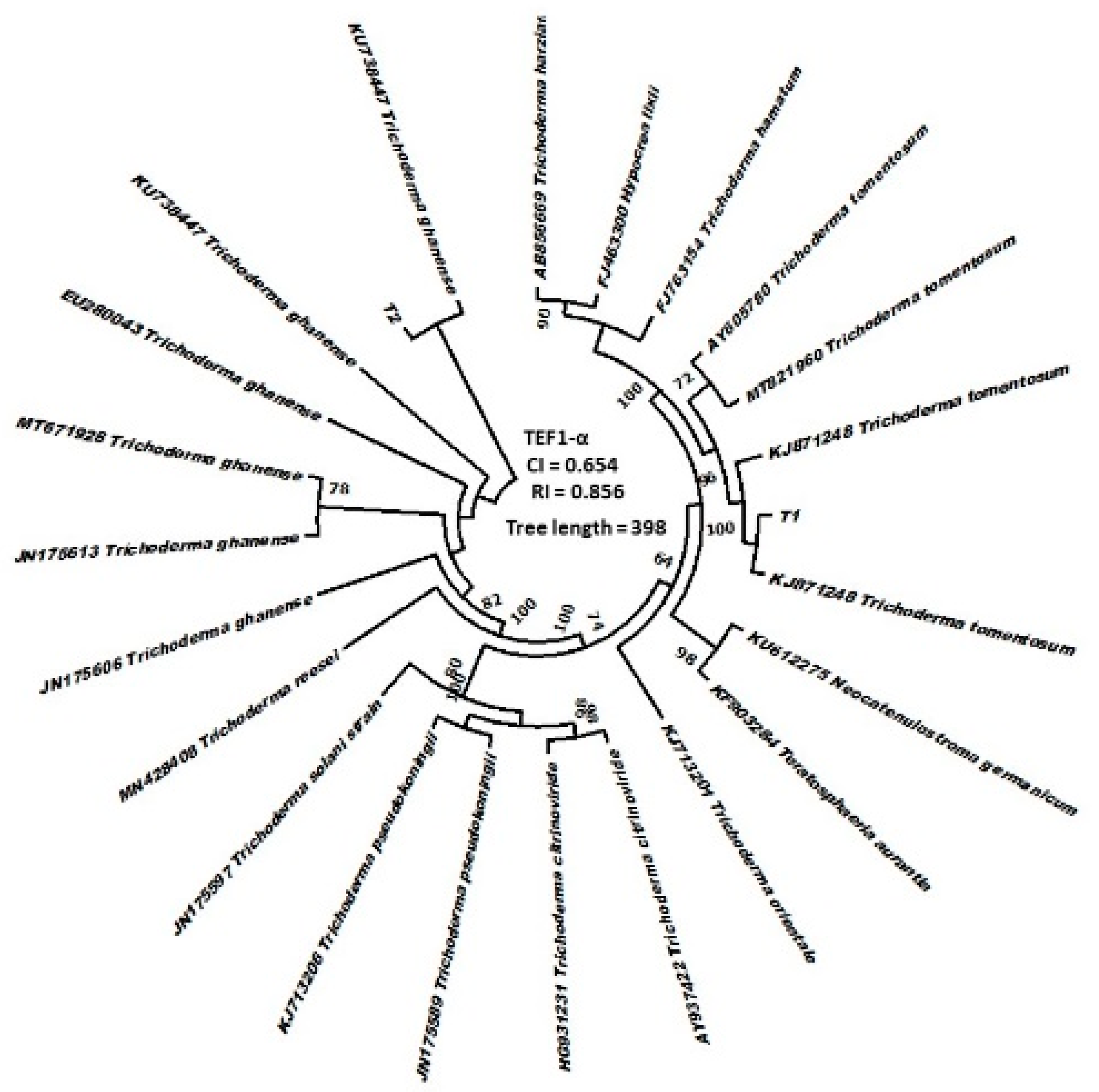
3.1. Identification of Trichoderma and Their Physiological Characteristics
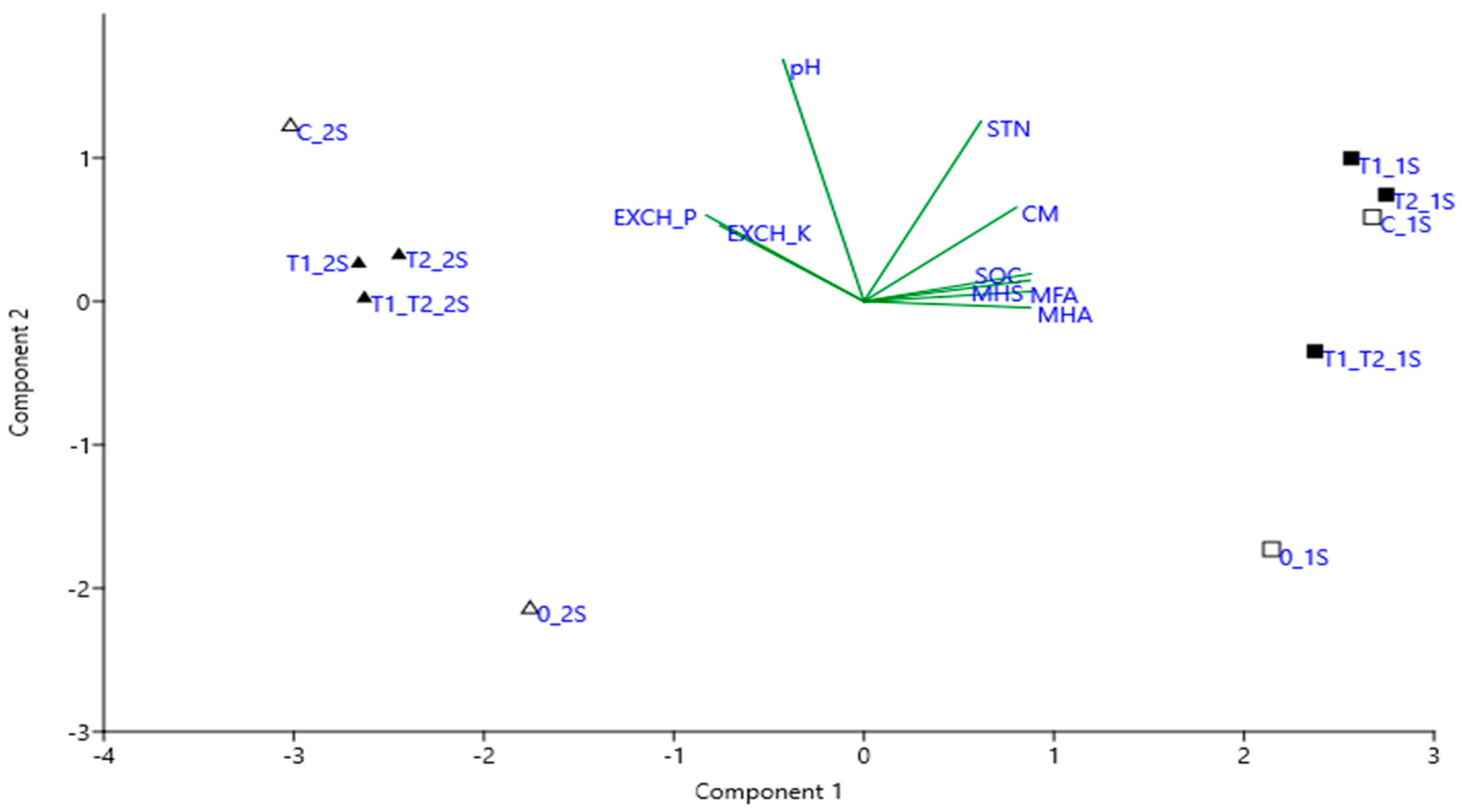
3.2. The Chemical Properties and CM-Cellulase Activity of Soil
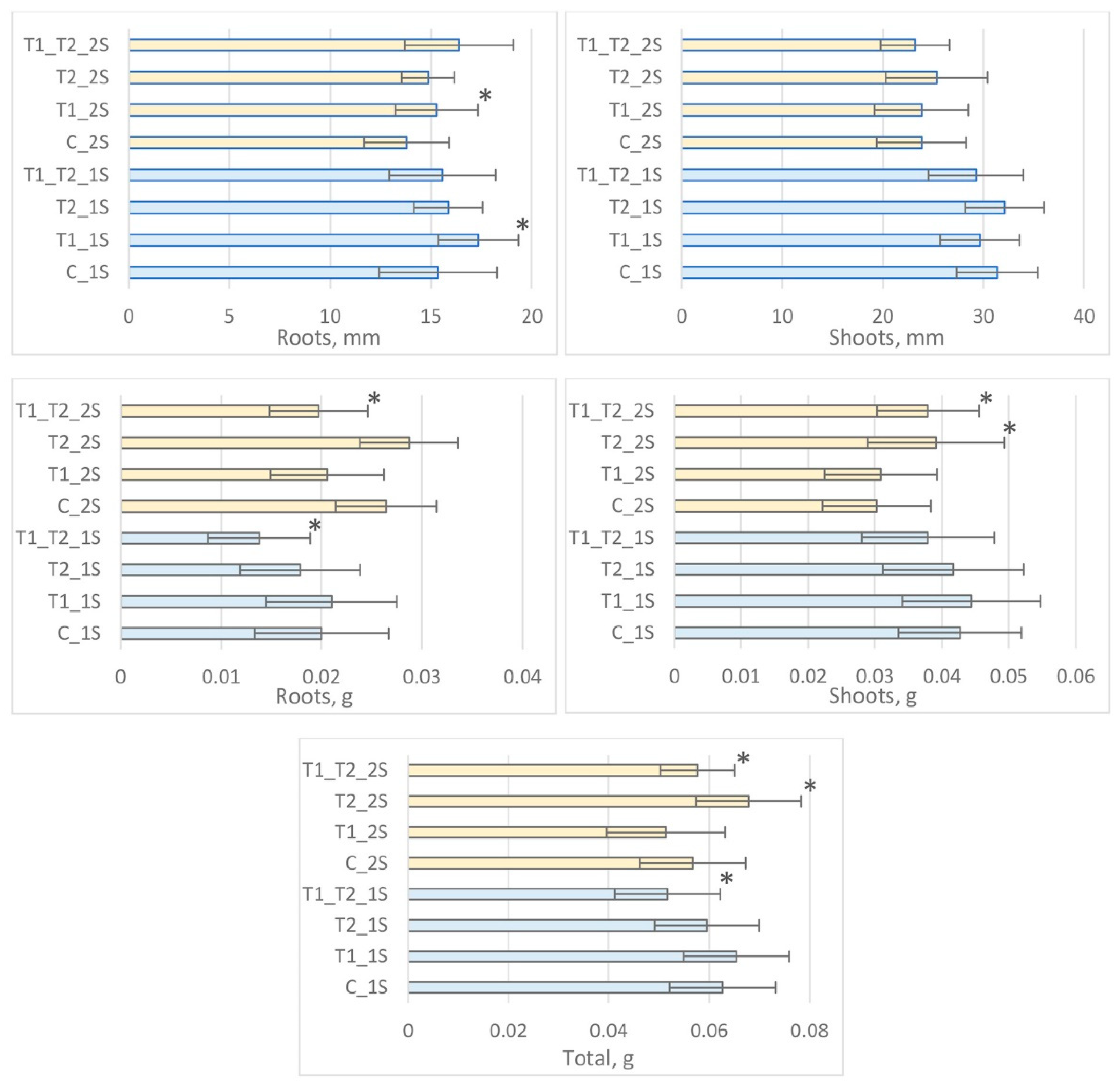
3.3. The Evaluation of Rye Seedling Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M. Impact of agrochemicals on soil microbiota and management: A Review. Land 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Lugtenberg, B.J.J.; Malfanova, N.; Kamilova, F.; Berg, G. Plant growth promotion by microbes. In Molecular Microbial Ecology of the Rhizosphere, 1st ed.; de Bruijn, F., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; Volume 2, pp. 559–573. [Google Scholar]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Aamir, M.; Rai Kumar, K.; Zehra, A.; Dubey Kumar, M.; Kumar, S.; Shukla, V.; Upadhyay, R.S. Microbial bioformulation-based plant biostimulants: A plausible approach toward next generation of sustainable agriculture. In Microbial Endophytes Functional Biology and Applications; Kumar, A., Radhakrishnan, E.K., Eds.; Woodhead Pub Ltd.: Cambridge, UK, 2020; pp. 195–225. [Google Scholar]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial biostimulants as response to modern agriculture needs: Composition, role and application of these innovative products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef]
- Saba, H.; Vibhash, D.; Manisha, M.; Prashant, K.S.; Farhan, H.; Tauseef, A. Trichoderma—A promising plant growth stimulator and biocontrol agent. Mycosphere 2014, 3, 524–531. [Google Scholar] [CrossRef]
- Nabi, S.U.; Malik, G.; Sharma, A. Trichoderma harzianum: A potential bioagent and plant growth promoter. EC Microbiol. 2018, 14, 147. [Google Scholar]
- Rajkonda, J.N.; Bhale, U.N. Influence of culture media on the growth of Trichoderma species. In Proceedings of the National Conference on Biodiversity: Prospects and Potentials at: Dept of Botany, S.M. Dnyandeo Mohekar Mahavidyala, Kalamb, Osmanabad, India, 28–29 December 2014; pp. 113–120. [Google Scholar]
- Asad, S.A.; Tabassum, A.; Hameed, A.; Hassan, F.U.; Afzal, A.; Khan, S.A.; Ahmed, R.; Shahzad, M. Determination of lytic enzyme activities of indigenous Trichoderma isolates from Pakistan. Braz. J. Microbiol. 2015, 46, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Cherkupally, R.; Amballa, H.; Reddy, B.N. In vitro screening for enzymatic activity of Trichoderma species for biocontrol potential. Ann. Plant Sci. 2017, 6, 1784–1789. [Google Scholar] [CrossRef]
- Novy, V.; Nielsen, F.; Seiboth, B.; Nidetzky, B. The influence of feedstock characteristics on enzyme production in Trichoderma reesei: A review on productivity, gene regulation and secretion profiles. Biotechnol. Biofuels 2019, 12, 238. [Google Scholar] [CrossRef]
- Hasan, S. Potential of Trichoderma sp. in bioremediation: A review. Int. J. Appl. Eng. Res. 2016, 3, 776–779. [Google Scholar]
- Banaay, C.G.B.; Cuevas, V.C.; Cruz, C.M.V. Trichoderma ghanense promotes plant growth and controls disease caused by Pythium arrhenomanes in seedlings of aerobic rice variety Apo. Philipp. Agric. Sci. 2012, 95, 175–184. [Google Scholar]
- Talla, S.G.; Raju, A.S.R.; Karri, S.; Kumar, Y.S. Production and antagonistic effect of Trichoderma spp. on pathogenic microorganisms (Botrytis cinerea, Fusarium oxysporum, Macrophomina phasealina and Rhizoctonia solani). Afr. J. Biotechnol. 2015, 14, 668–675. [Google Scholar]
- Herrera-Jiménez, E.; Alarcón, A.; Larsen, J.; Ferrera-Cerrato, R.; Cruz-Izquierdo, S.; Ferrera-Rodríguez, M.R. Comparative effects of two indole-producing Trichoderma strains and two exogenous phytohormones on the growth of Zea mays L., with or without tryptophan. J. Soil Sci. Plant Nut. 2018, 18, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defence system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwhibi, M.S.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar]
- Khoshmanzar, E.; Aliasgharzad, N.; Neyshabouri, M.R.; Khoshru, B.; Arzanlou, M.; Asgari Lajayer, B. Effects of Trichoderma isolates on tomato growth and inducing its tolerance to water-deficit stress. Int. J. Environ. Sci. Technol. 2020, 17, 869–878. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M.; Lopez, A.; Martos, V.; Reyes, A.; Maksimovic, I.; Eichler-Löbermann, B.; Malusà, E. Unexploited potential of some biotechnological techniques for biofertilizer production and formulation. Appl. Microbiol. Biotechnol. 2015, 99, 4983–4996. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Caporale, A.G.; Vitaglione, P.; Troise, A.D.; Pigna, M.; Ruocco, M. Influence of three different soil types on the interaction of two strains of Trichoderma harzianum with Brassica rapa subsp. sylvestris cv. esculenta, under soil mineral fertilization. Geoderma 2019, 350, 11–18. [Google Scholar] [CrossRef]
- Germida, J.J.; de Freitas, J.R. Cultural methods for soil and root-associated microorganisms. In Soil Sampling and Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press Taylor & Francis Group: Sound Parkway, NW, USA, 2007; pp. 341–354. [Google Scholar]
- Chaverri, P.; Castlebury, L.A.; Barrie, E.O.; Gary, J.S. Hypocrea/Trichoderma: Species with conidiophore elongations and green conidia. Mycologia 2003, 95, 1100–1140. [Google Scholar] [CrossRef]
- Samuels, G.J.; Ismaiel, A.; Mulaw, T.B.; Szakacs, G.; Druzhinina, I.S.; Kubicek, C.P.; Jaklitsch, W.M. The Longibrachiatum Clade of Trichoderma: A revision with new species. Fungal Divers. 2012, 55, 77–108. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, S.S., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Nikolcheva, L.G.; Bärlocher, F. Taxon-specific fungal primers reveal unexpectedly high diversity during leaf decomposition in a stream. Mycol. Prog. 2004, 3, 41–49. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Bilai, V.I. Methods of Experimental Mycology; Naukova Dumka: Kiev, Ukraine, 1982; p. 550. [Google Scholar]
- Ravin, H.A.; Harward, M.D. Rapid test for hepatolenticular degeneration. Lancet 1965, 267, 726–727. [Google Scholar] [CrossRef]
- Jermakov, A.I.; Arasimovitsh, V.V.; Jarosh, N.P.; Peruanskij, U.A.; Lukovnikova, G.A.; Ikonnikova, M.I. Methods of Plant Biochemical Research; Agropromizdat: Leningrad, Russia, 1987; p. 430. [Google Scholar]
- Chudiakova, C.K. Determination of the lignin content in fodder and green crop. Selskochoziaistvenaja Biol. 1984, 8, 120–124. [Google Scholar]
- Nikitin, B.A. Method for soil humus determination. Agric. Chem. 1999, 3, 156–158. [Google Scholar]
- Šlepetienė, A.; Kadžiulienė, Ž.; Feizienė, D.; Liaudanskienė, I.; Amalevičiūtė-Volungė, K.; Šlepetys, J. The distribution of organic carbon, its forms and macroelements in agricultural soils. Zemdirbyste 2020, 107, 291–300. [Google Scholar] [CrossRef]
- Ponomareva, V.V.; Plotnikova, T.A. Humus and Soil Formation; Nauka: Leningrad, Russia, 1980. [Google Scholar]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer-Verlag: Berlin/Heidelberg, Germany, 1996; p. 416. [Google Scholar]
- ISO 11269-2:2012 Soil Quality—Determination of Pollutants on Soil Flora. Part 2: Effect of Contaminated Soil on the Emergence and Early Growth of Higher Plants. Available online: www.iso.org/standard/51382.html (accessed on 14 January 2022).
- Singh, D.P.; Singh, H.B.; Prabha, R. Microbial Inoculants in Sustainable Agricultural Productivity; Functional Applications; Springer: New Delhi, India, 2016; Volume 2, p. 308. [Google Scholar]
- Muniappan, V.; Muthukumar, T.M. Influence of crop species and edaphic factors on the distribution and abundance of Trichoderma in Alfisol soils of southern India. Acta Bot. Croat. 2014, 73, 37–50. [Google Scholar] [CrossRef] [Green Version]
- van Elsas, J.D.; Jansson, J.K.; Trevors, J.T. Modern Soil Microbiology, 2nd ed.; CRC Press Taylor and Francis Group Boca Raton: Boca Raton, FL, USA, 2006; p. 698. [Google Scholar]
- Debnath, S.; Chakraborty, G.; Dutta, S.S.; Chaudhuri, S.R.; Das, P.; Saha, A.K. Potential of Trichoderma species as biofertilizer and biological control on Oryza sativa L. cultivation. Biotecnol. Veg. 2020, 20, 1–6. [Google Scholar]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 2020, 10, 2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.; Liu, Z.; Liu, B.; Wang, Y.; Wang, J. The effect of Trichoderma biofertilizer on the quality of flowering Chinese cabbage and the soil environment. Sci. Hortic. 2020, 262, 109069. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Del Mar Alguacil, M.; Pascual, J.A.; van Wees, S.C.M. Phytohormone profiles induced by Trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef]
- Hajieghrari, B.; Mohammadi, M. Growth-promoting activity of indigenous Trichoderma isolates on wheat seed germination, seedling growth and yield. Aust. J. Crop Sci. 2016, 10, 1339–1347. [Google Scholar] [CrossRef]
- Tančić-Živanov, S.; Medić-Pap, S.; Prvulovic, D. Effect of Trichoderma spp. on Growth Promotion and Antioxidative Activity of Pepper Seedlings. Braz. Arch. Biol. Technol. 2020, 63, e20180659. [Google Scholar] [CrossRef]
- Cooper, R.J.; Liu, C.; Fisher, D.S. Influence of humic substances on rooting and nutrient content of creeping bentgrass. Crop Sci. 1998, 38, 1639–1644. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. 2014, 1, 3. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Properties | Grassland | Arable Soil | Methods |
|---|---|---|---|
| Soil organic carbon (SOC), % | 2.49 | 1.43 | Spectrophotometric measurement method at 590 nm using glucose as a standard after wet combustion [39]. |
| Humus, % | 4.30 | 2.46 | Calculated using conversion factor (1.724) from SOC. |
| Total N (Nt), % | 0.115 | 0.080 | Kjeldahl method using a spectrophotometric measurement at 655 nm. |
| C/N | 21.70 | 17.90 | Calculation as ratio of SOC to Nt. |
| Exchangeable P2O5, mg/kg | 46.90 | 97.70 | A-L method. |
| Exchangeable K2O, mg/kg | 98.81 | 162.90 | A-L method. |
| Labile water-soluble carbon, CH2O, g/kg | 0.299 | 0.106 | Determined by IR detection after UV-catalyzed persulfate oxidation [40]. |
| Mobile humic substances, (MHS) % | 0.383 | 0.194 | Extracted with 0.1 M NaOH [41]. |
| Mobile humic acids, % | 0.168 | 0.092 | Determined in 0.1 M NaOH extract. |
| Mobile fulvic acids, % | 0.215 | 0.102 | Determined in 0.1 M NaOH extract. |
| pH | 6.92 | 6.83 | Determined by the potentiometric method in 1 M KCl (1:2.5, w/v) extract. |
| CM-cellulase activity, µg GE g−1 · dm · 24 h−1 | 456.70 | 100.40 | Determined by the spectrophotometric method [42]. |
| Strain | Sequence Area Characterization |
|---|---|
| T. tomentosum T1 |  |
| T. tomentosum AY605737 | |
| T. ghanense T2 |  |
| T. ghanense EU280100 |
| Strain | Sequence Area Characterization |
|---|---|
| T. tomentosum T1 |  |
| T. tomentosum KJ871248 | |
| T. tomentosum T2 |  |
| T. ghanense EU280043 |
| Physiological Characteristics | T. ghanense | T. tomentosum |
|---|---|---|
| Growth (diameter, mm) on PDA at 7 °C | 5.0 ± 0.0 | 5.0 ± 0.0 |
| 15 °C | 37.0 ± 1.7 * | 43.2 ± 2.4 * |
| 26 °C | 46.6 ± 2.3 * | 45.1 ± 1.7 * |
| 35 °C | 56.2 ± 2.3 * | 50.0 ± 0.0 * |
| Growth (diameter, mm) on CzDA at 26 °C and pH 4 | 55.0 ± 0.0 * | 73.7 ± 0.5 * |
| 5 | 61.0 ± 1.7 * | 67.6 ± 0.6 * |
| 6 | 53.3 ± 2.8 * | 63.0 ± 0.0 * |
| 7 | 51.6 ± 2.8 * | 42.0 ± 1.0 * |
| 8 | 51.0 ± 1.7 * | 36.0 ± 0.0 * |
| Decomposition of cellulose, % | 32.7 ± 1.3 * | 25.6 ± 0.9 * |
| Decomposition of lignin, % | 11.0 ± 0.8 * | 14.2 ± 0.7 * |
| Enzymatic activity: | ||
| endoglucanase, % | 120.6 ± 2.8 * | 10.8 ± 1.4 * |
| peroxidase, a.u./g | 106.0 ± 1.4 * | 90.0 ± 4.2 * |
| tyrosinase, c.u./g | 0.033 ± 0.0 | 0.72 ± 0.0 |
| laccase, a.u./g | 0.004 ± 0.0 | 0.004 ± 0.0 |
| Chemical Properties of Soil | Variants | Control—Non Inoculated Soil | ||
|---|---|---|---|---|
| T. ghanense | T. tomentosum | T. ghanense + T. tomentosum | ||
| 1S | 1S | 1S | 1S | |
| SOC, % | 2.72 ± 0.02 | 2.68 ± 0.06 | 2.74 ± 0.08 | 2.75 ± 0.18 |
| Humus, % | 4.69 ± 0.07 | 4.61 ± 0.11 | 4.73 ± 0.11 | 4.74 ± 0.01 |
| Total N, % | 0.208 ± 0.001 | 0.215 ± 0.003 * | 0.207 ± 0.008 | 0.204 ± 0.004 |
| C/N | 13.1 | 12.5 | 13.2 | 13.5 |
| Exchangeable P2O5, mg/kg | 71.90 ± 0.57 | 71.60 ± 0.00 * | 71.60 ± 0.70 | 69.30 ± 0.28 |
| Exchangeable K2O, mg/kg | 124.50 ± 0.41 * | 128.50 ± 0.13 * | 124.20 ± 0.08 * | 135.49 ± 0.14 |
| Labile water-soluble carbon, CH2O, g/kg | 0.268 ± 0.019 | 0.275 ± 0.026 | 0.314 ± 0.012 | 0.291 ± 0.019 |
| Mobile humic substances, % | 0.406 ± 0.08 | 0.434 ± 0.004 | 0.422 ± 0.014 | 0.426 ± 0.013 |
| Mobile humic acids, % | 0.190 ± 0.001 | 0.179 ± 0.004 * | 0.198 ± 0.004 * | 0.188 ± 0.003 |
| Mobile fulvic acids, % | 0.216 ± 0.012 * | 0.255 ± 0.008 | 0.224 ± 0.003 | 0.238 ± 0.004 |
| pH | 7.08 ± 0.01 * | 7.02 ± 0.01 * | 7.02 ± 0.014 | 7.00 ± 0.00 |
| CM-cellulase activity, µg GE g−1 · dm · 24h−1 | 896.00 ± 3.46 | 930.20 ± 1.91 * | 849.40 ± 7.20* | 897.60 ± 2.622 |
| Chemical Properties of Soil | Variants | Control—Non Inoculated Soil | ||
|---|---|---|---|---|
| T. ghanense | T. tomentosum | T. ghanense + T. tomentosum | ||
| 2S | 2S | 2S | 2S | |
| SOC, % | 1.39 ± 0.05 | 1.43 ± 0.01 | 1.33 ± 0.01 | 1.38 ± 0.05 |
| Humus, % | 2.40 ± 0.06 | 2.46 ± 0.01 | 2.30 ± 0.06 | 2.38 ± 0.11 |
| Total N, % | 0.105 ± 0.001 | 0.109 ± 0.001 * | 0.114 ± 0.001 | 0.123 ± 0.004 |
| C/N | 13.2 | 13.1 | 11.7 | 11.2 |
| Exchangeable P2O5, mg/kg | 134.40 ± 1.20 | 130.60 ± 1.13 | 129.60 ± 0.14 | 127.40 ± 2.69 |
| Exchangeable K2O, mg/kg | 163.40 ± 0.68 * | 155.70 ± 0.70 * | 149.90 ± 0.68 * | 178.30 ± 1.34 |
| Labile water-soluble carbon, CH2O, g/kg | 0.194 ± 0.015 | 0.197 ± 0.013 | 0.182 ± 0.019* | 0.199 ± 0.021 |
| Mobile humic substances, % | 0.146 ± 0.006 | 0.151 ± 0.004 | 0.144 ± 0.006 | 0.157 ± 0.003 |
| Mobile humic acids, % | 0.050 ± 0.001 | 0.061 ± 0.002 | 0.051 ± 0.001 * | 0.059 ± 0.000 |
| Mobile fulvic acids, % | 0.096 ± 0.001 | 0.090 ± 0.004 | 0.093 ± 0.004 | 0.098 ± 0.001 |
| pH | 7.13 ± 0.01 * | 7.15 ± 0.00 | 7.13 ± 0.01 * | 7.28 ± 0.00 |
| CM-cellulase activity, µg GE g−1 · dm · 24 h−1 | 114.80 ± 3.79 * | 324.60 ± 3.47 * | 332.80 ± 4.30 * | 114.50 ± 3.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bridžiuvienė, D.; Raudonienė, V.; Švedienė, J.; Paškevičius, A.; Baužienė, I.; Vaitonis, G.; Šlepetienė, A.; Šlepetys, J.; Kačergius, A. Impact of Soil Chemical Properties on the Growth Promotion Ability of Trichoderma ghanense, T. tomentosum and Their Complex on Rye in Different Land-Use Systems. J. Fungi 2022, 8, 85. https://doi.org/10.3390/jof8010085
Bridžiuvienė D, Raudonienė V, Švedienė J, Paškevičius A, Baužienė I, Vaitonis G, Šlepetienė A, Šlepetys J, Kačergius A. Impact of Soil Chemical Properties on the Growth Promotion Ability of Trichoderma ghanense, T. tomentosum and Their Complex on Rye in Different Land-Use Systems. Journal of Fungi. 2022; 8(1):85. https://doi.org/10.3390/jof8010085
Chicago/Turabian StyleBridžiuvienė, Danguolė, Vita Raudonienė, Jurgita Švedienė, Algimantas Paškevičius, Ieva Baužienė, Gintautas Vaitonis, Alvyra Šlepetienė, Jonas Šlepetys, and Audrius Kačergius. 2022. "Impact of Soil Chemical Properties on the Growth Promotion Ability of Trichoderma ghanense, T. tomentosum and Their Complex on Rye in Different Land-Use Systems" Journal of Fungi 8, no. 1: 85. https://doi.org/10.3390/jof8010085
APA StyleBridžiuvienė, D., Raudonienė, V., Švedienė, J., Paškevičius, A., Baužienė, I., Vaitonis, G., Šlepetienė, A., Šlepetys, J., & Kačergius, A. (2022). Impact of Soil Chemical Properties on the Growth Promotion Ability of Trichoderma ghanense, T. tomentosum and Their Complex on Rye in Different Land-Use Systems. Journal of Fungi, 8(1), 85. https://doi.org/10.3390/jof8010085