
Melanization of Candida auris Is Associated with Alteration of Extracellular pH

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Candida auris Strains and Media
2.2. C. auris Melanization in Liquid Media
2.3. Melanin Extraction and Electron Paramagnetic Resonance (EPR)
2.4. Extracellular Melanin Isolation
2.5. Supernatant Melanization Activity
2.6. Light Microscopy
2.7. Transmission Electron Microscopy (TEM)
2.8. Scanning Electron Microscopy (SEM)
2.9. Extracellular Melanin Add-Back
2.10. Cell Wall Disruption Assays
2.11. Preparation of Fungal Cells for ssNMR Analysis
2.12. Solid-State NMR Spectroscopy
2.13. pH Measurements and Supernatant pH Alteration
2.14. Ammonia Quantification
2.15. Cell Surface Hydrophobicity (CSH)
2.16. Oxidative Stress
2.17. Killing by Bone Marrow-Derived Macrophages
2.18. Galleria Mellonella Infection
3. Results
3.1. Temperature Dependence of Melanization on Candida auris Strains
3.2. Electron Paramagnetic Resonance (EPR)
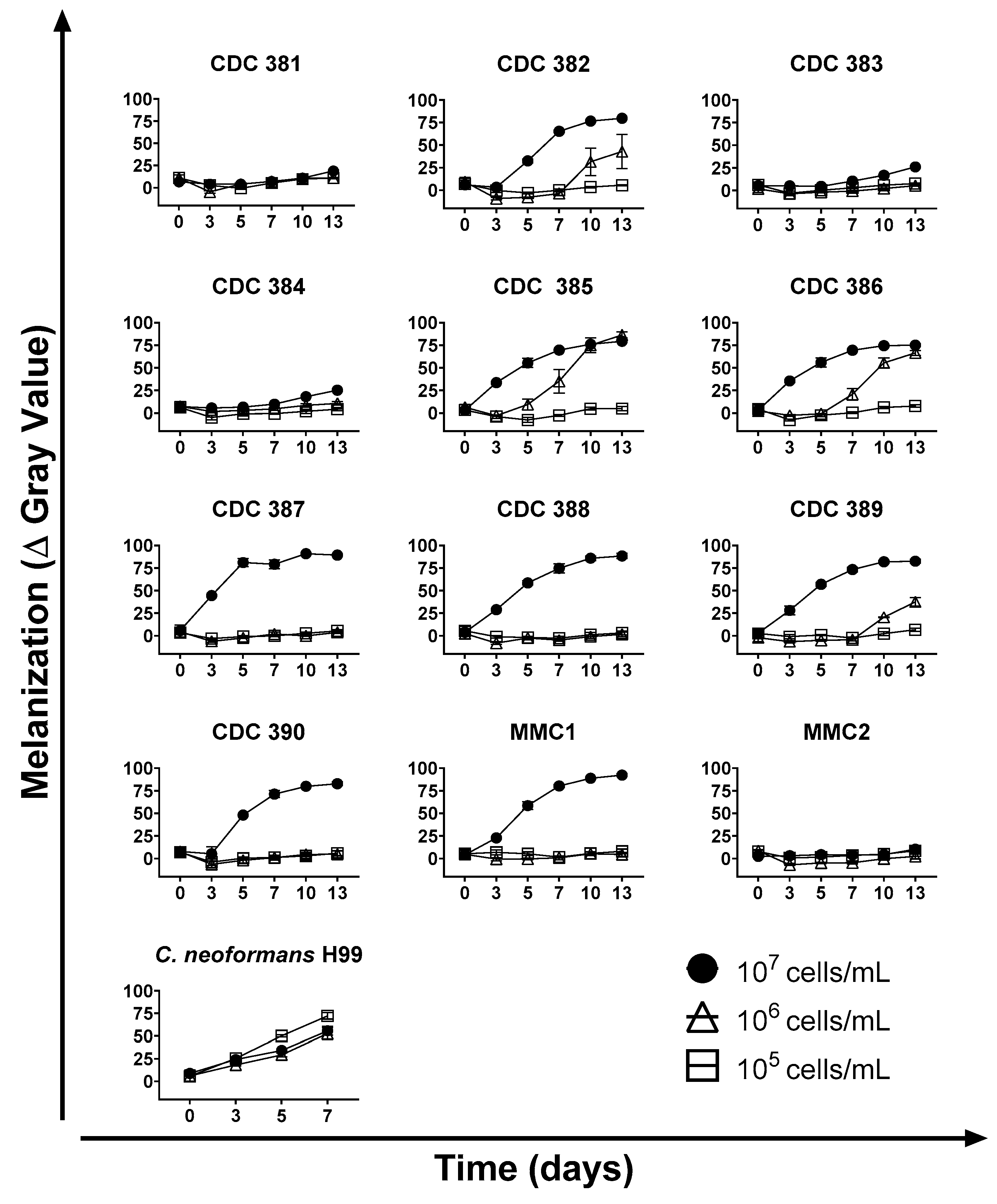
3.3. Time and Cell Density
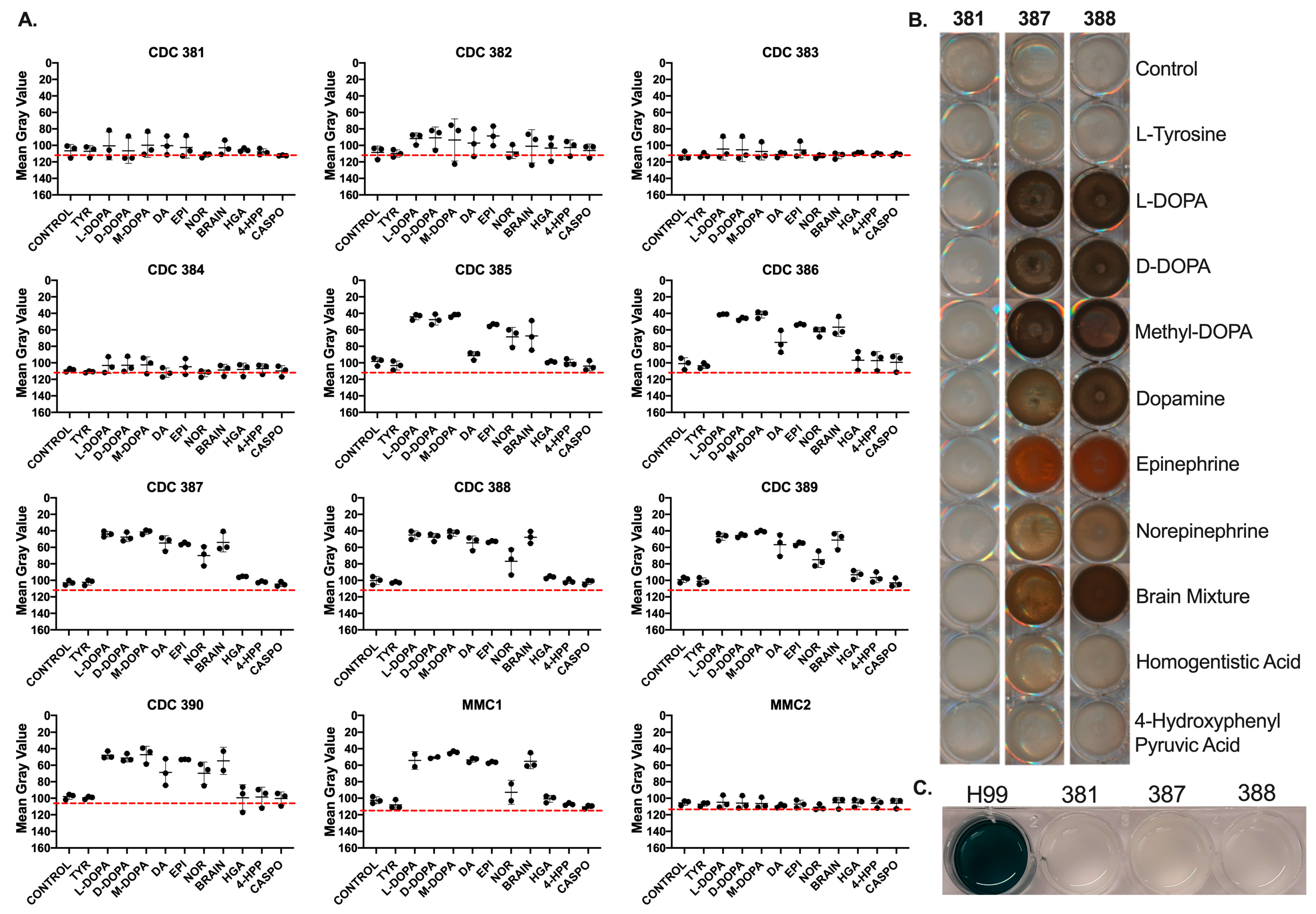
3.4. Candida auris Strains Melanize Using Substrates Associated with DOPA Melanin
3.5. Melanin Is Primarily Found in the Supernatant
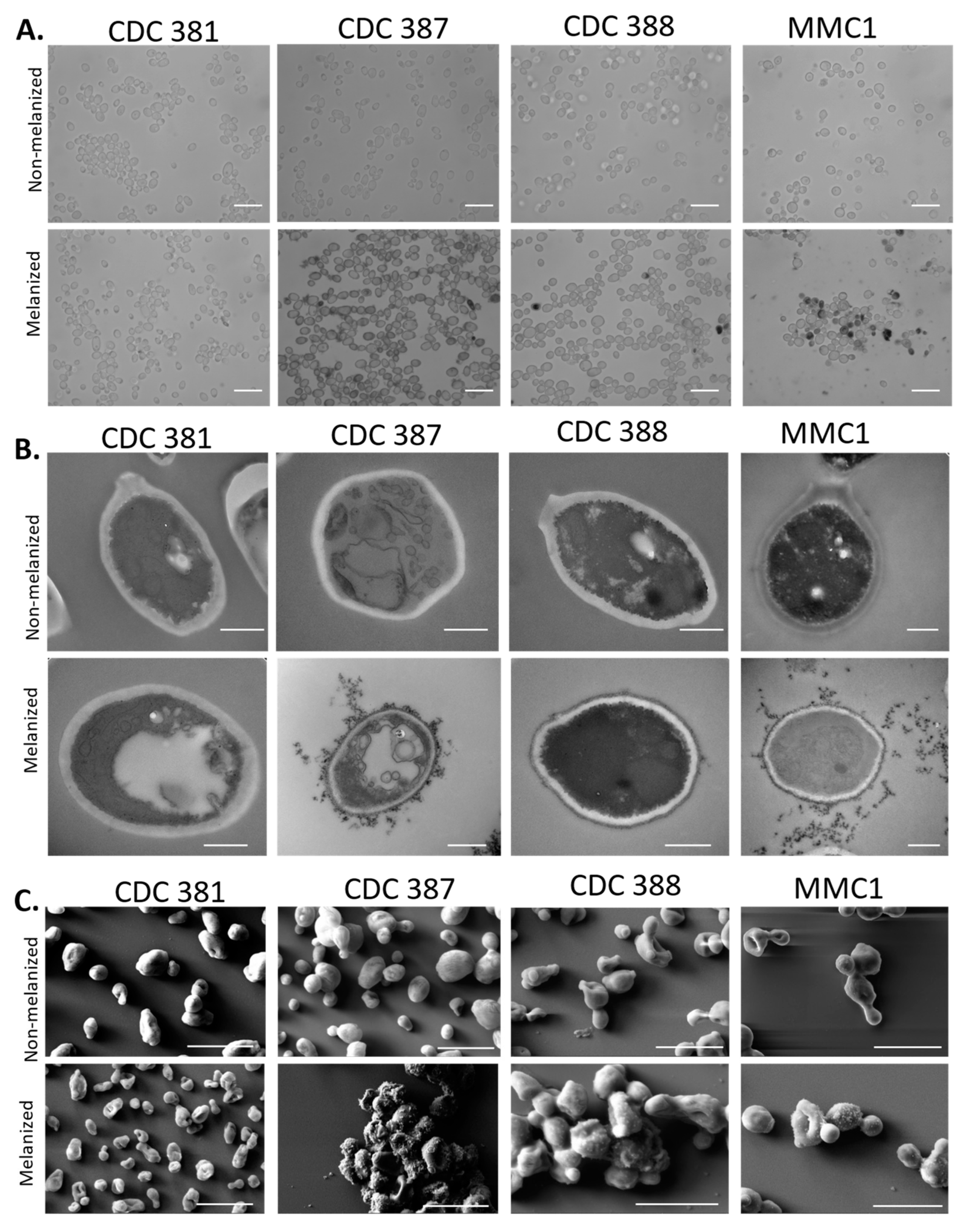
3.6. Cell-Bound Melanin Is Localized to the Periphery of the Cell Wall
3.7. Effects of Altering Cell Wall Structures on the Melanization of C. auris
3.8. Melanization Affects the Hydrophobicity of Some C. auris Strains
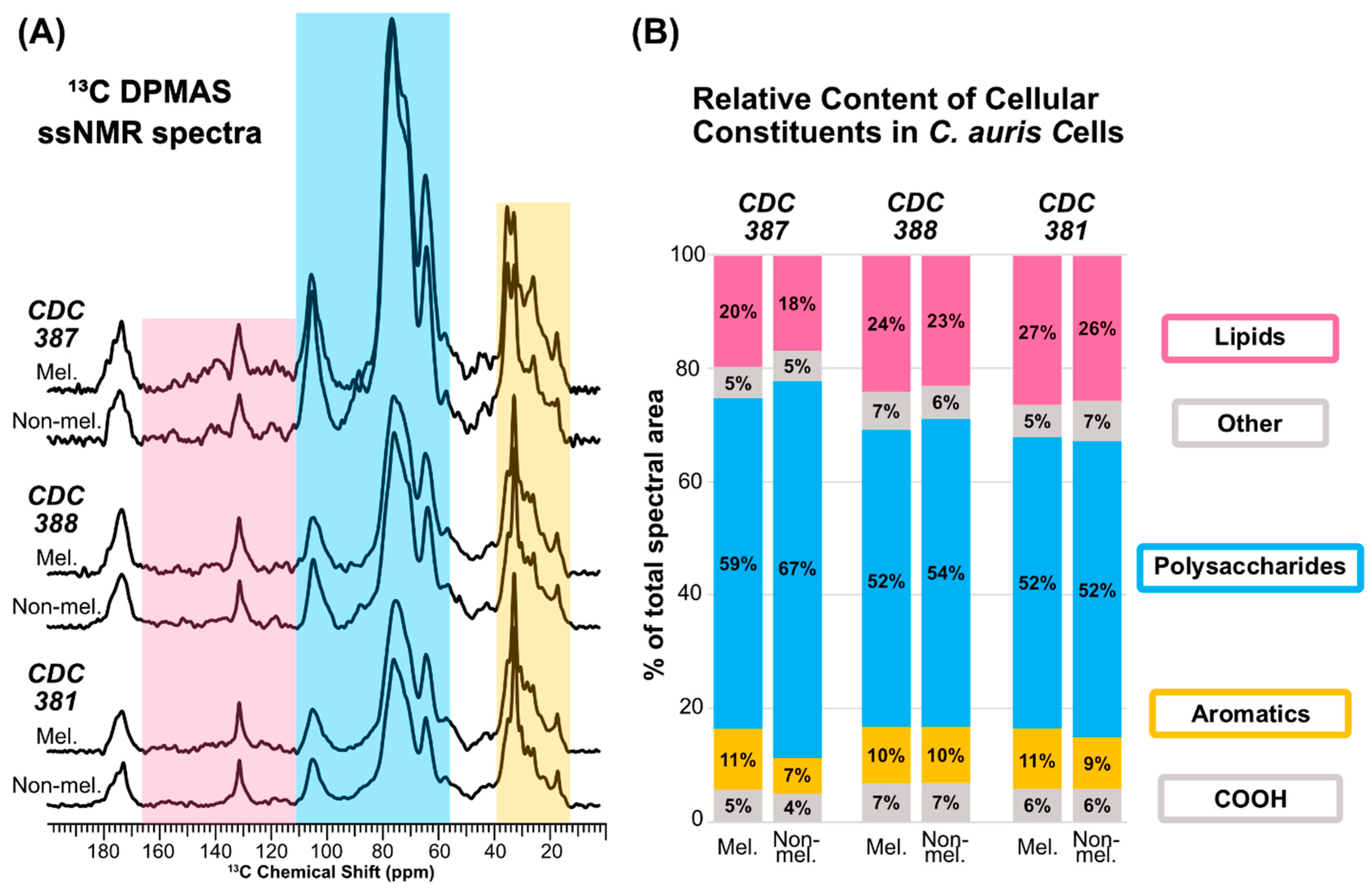
3.9. Melanin Adherence Is Correlated with a Higher Cell-Wall Polysaccharide Content
3.10. Supernatant Melanization Is Resistant to Denaturing Conditions
3.11. Melanization Genes in the C. auris Genome
3.12. Supernatant Neutral pH Correlates with Ability of C. auris Strains to Melanize
3.13. Melanin Protects C. auris from Oxidative Damage
3.14. Melanin Does Not Protect C. auris against Macrophage Killing
3.15. Melanin Does Not Affect C. auris Virulence during Galleria Mellonella In Vivo Infection
4. Discussion
4.1. Characterization of Melanin Production and Localization
4.2. Mechanism of Melanin Production
4.3. Implications of C. auris Melanization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10, e01397-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, P.; Singh, P.; Wang, Y.; Yadav, A.; Pawar, K.; Singh, A.; Padmavati, G.; Xu, J.; Chowdhary, A. Environmental Isolation of Candida auris from the Coastal Wetlands of Andaman Islands, India. mBio 2021, 12, e03181-20. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Alfouzan, W. Candida auris: Epidemiology, Diagnosis, Pathogenesis, Antifungal Susceptibility, and Infection Control Measures to Combat the Spread of Infections in Healthcare Facilities. Microorganisms 2021, 9, 807. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.-C. First Three Reported Cases of Nosocomial Fungemia Caused by Candida auris. J. Clin. Microbiol. 2011, 49, 3139–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida aurissp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.V.; Nett, J.E. Candida auris Infection and Biofilm Formation: Going Beyond the Surface. Curr. Clin. Microbiol. Rep. 2020, 7, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bustos, V.; Cabanero-Navalon, M.D.; Ruiz-Saurí, A.; Ruiz-Gaitán, A.C.; Salavert, M.; Tormo, M.Á.; Pemán, J. What Do We Know about Candida auris? State of the Art, Knowledge Gaps, and Future Directions. Microorganisms 2021, 9, 2177. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.; da Silva, L.B.; Gerfen, G.; Araújo, G.R.D.S.; Frases, S.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Candida glabrata produces a melanin-like pigment that protects against stress conditions encountered during parasitism. Futur. Microbiol. 2021, 16, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.A.; Gómez, B.L.; Mora-Montes, H.M.; Mackenzie, K.S.; Munro, C.A.; Brown, A.J.P.; Gow, N.A.R.; Kibbler, C.C.; Odds, F.C. Melanin Externalization in Candida albicans Depends on Cell Wall Chitin Structures. Eukaryot. Cell 2010, 9, 1329–1342. [Google Scholar] [CrossRef] [Green Version]
- Morris-Jones, R.; Gomez, B.L.; Diez, S.; Uran, M.; Morris-Jones, S.D.; Casadevall, A.; Nosanchuk, J.D.; Hamilton, A.J. Synthesis of Melanin Pigment by Candida albicans In Vitro and during Infection. Infect. Immun. 2005, 73, 6147–6150. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Xu, Y.; Gao, N.; Chen, J. Wor1-regulated ferroxidases contribute to pigment formation in opaque cells of Candida albicans. FEBS Open Bio. 2021, 11, 598–621. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.F.Q.; Casadevall, A. The Role of Melanin in Fungal Pathogenesis for Animal Hosts. In Fungal Physiology and Immunopathogenesis [Internet]; Rodrigues, M.L., Ed.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–30. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Sun, N.; Xiao, D.; Pan, J.; Wang, Y.; Zhu, X. A copper-responsive factor gene CUF1 is required for copper induction of laccase in Cryptococcus neoformans. FEMS Microbiol. Lett. 2009, 296, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauch, R.M.; Cunha, V.D.O.; Dias, A.L.T. The copper interference with the melanogenesis of Cryptococcus neoformans. Rev. Inst. Med. Trop. S Paulo. 2013, 55, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero, R.J.B.; Camacho, E.; Casadevall, A. Melanization in Cryptococcus neoformans Requires Complex Regulation. mBio 2020, 11, e03313-19. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Kendall, T.; Yip, P.; Davy, A.; Sefcik, J.; Sutter, J.U. Influence of Ions and pH on the Formation of Solid- and Liquid-like Melanin. ACS Omega. 2020, 5, 25059–25068. [Google Scholar] [CrossRef]
- Eslami, M.; Zare, H.R.; Namazian, M. Thermodynamic Parameters of Electrochemical Oxidation of l-DOPA: Experimental and Theoretical Studies. J. Phys. Chem. B 2012, 116, 12552–12557. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O’Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; et al. The structural unit of melanin in the cell wall of the fungal pathogen Cryptococcus neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef]
- Franzen, A.J.; Cunha, M.M.; Miranda, K.; Hentschel, J.; Plattner, H.; da Silva, M.B.; Salgado, C.G.; de Souza, W.; Rozental, S. Ultrastructural characterization of melanosomes of the human pathogenic fungus Fonsecaea pedrosoi. J. Struct. Biol. 2008, 162, 75–84. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the Deacetylated Form of Chitin, Is Necessary for Cell Wall Integrity in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Walton, F.J.; Idnurm, A.; Heitman, J. Novel gene functions required for melanization of the human pathogen Cryptococcus neoformans. Mol. Microbiol. 2005, 57, 1381–1396. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T. Chemical Composition of Wild-type and Mutant Aspergillus nidulans Cell Walls. The Nature of Polysaccharide and Melanin Constituents. J. Gen. Microbiol. 1970, 63, 75–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrissian, C.; Camacho, E.; Kelly, J.E.; Wang, H.; Casadevall, A.; Stark, R.E. Solid-state NMR spectroscopy identifies three classes of lipids in Cryptococcus neoformans melanized cell walls and whole fungal cells. J. Biol. Chem. 2020, 295, 15083–15096. [Google Scholar] [CrossRef] [PubMed]
- Chrissian, C.; Lin, C.P.C.; Camacho, E.; Casadevall, A.; Neiman, A.M.; Stark, R.E. Unconventional Constituents and Shared Molecular Architecture of the Melanized Cell Wall of C. neoformans and Spore Wall of S. cerevisiae. J. Fungi. 2020, 6, 329. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Nosanchuk, J.D.; Webber, J.B.W.; Emerson, R.J.; Camesano, A.T.A.; Casadevall, A. Microstructure of Cell Wall-Associated Melanin in the Human Pathogenic Fungus Cryptococcus neoformans. Biochemistry 2005, 44, 3683–3693. [Google Scholar] [CrossRef]
- Jacobson, E.S.; Tinnell, S.B. Antioxidant function of fungal melanin. J. Bacteriol. 1993, 175, 7102–7104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alviano, D.S.; Franzen, A.J.; Travassos, L.R.; Holandino, C.; Rozental, S.; Ejzemberg, R.; Alviano, C.S.; Rodrigues, M.L. Melanin from Fonsecaea pedrosoi Induces Production of Human Antifungal Antibodies and Enhances the Antimicrobial Efficacy of Phagocytes. Infect. Immun. 2004, 72, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doering, T.L.; Nosanchuk, J.D.; Roberts, W.K.; Casadevall, A. Melanin as a potential cryptococcal defence against microbicidal proteins. Med. Mycol. 1999, 37, 175–181. [Google Scholar] [CrossRef]
- Rosas, Á.L.; Casadevall, A. Melanization decreases the susceptibility of Cryptococcus neoformans to enzymatic degradation. Mycopathologia 2001, 151, 53–56. [Google Scholar] [CrossRef]
- Stappers, M.H.T.; Clark, A.E.; Aimanianda, V.; Bidula, S.; Reid, D.M.; Asamaphan, P.; Brown, G.D. Recognition of DHN-melanin by MelLec, is required for protective immunity to Aspergillus. Nature 2018, 555, 382–386. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Rosas, A.L.; Casadevall, A. The antibody response to fungal melanin in mice. J. Immunol. 1998, 160, 6026–6031. [Google Scholar] [PubMed]
- Rosas, A.L.; Nosanchuk, J.D.; Casadevall, A. Passive Immunization with Melanin-Binding Monoclonal Antibodies Prolongs Survival of Mice with Lethal Cryptococcus neoformans Infection. Infect. Immun. 2001, 69, 2286–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panel Details | Antimicrobial Resistance Isolate Bank | Antibiotic/Antimicrobial Resistance | CDC [Internet]. Available online: https://wwwn.cdc.gov/ARIsolateBank/Panel/PanelDetail?ID=2 (accessed on 19 January 2022).
- Zamith-Miranda, D.; Heyman, H.M.; Couvillion, S.P.; Cordero, R.J.B.; Rodrigues, M.L.; Nimrichter, L.; Casadevall, A.; Amatuzzi, R.F.; Alves, L.R.; Nakayasu, E.S.; et al. Comparative Molecular and Immunoregulatory Analysis of Extracellular Vesicles from Candida albicans and Candida auris. mSystems 2021, 6, e00822-21. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.; Prados-Rosales, R.; Itin, B.; Casadevall, A.; Stark, R.E. Solid-state NMR Reveals the Carbon-based Molecular Architecture of Cryptococcus neoformans Fungal Eumelanins in the Cell Wall. J. Biol. Chem. 2015, 290, 13779–13790. [Google Scholar] [CrossRef] [Green Version]
- Vij, R.; Danchik, C.; Crawford, C.; Dragotakes, Q.; Casadevall, A. Variation in Cell Surface Hydrophobicity among Cryptococcus neoformans Strains Influences Interactions with Amoebas. mSphere 2020, 5, e00310-20. [Google Scholar] [CrossRef] [PubMed]
- Williamson, B.; Wilk, A.; Guerrero, K.D.; Mikulski, T.D.; Elias, T.N.; Sawh, I.; Healey, K.R. Impact of Erg11 Amino Acid Substitutions Identified in Candida auris Clade III Isolates on Triazole Drug Susceptibility. Antimicrob. Agents Chemother. 2022, 66, e01624-21. [Google Scholar] [CrossRef]
- Ademe, M.; Girma, F. Candida auris: From Multidrug Resistance to Pan-Resistant Strains. Infect. Drug Resist. 2020, 13, 1287–1394. [Google Scholar]
- Baker, R.P.; Chrissian, C.; Stark, R.E.; Casadevall, A. Cryptococcus neoformans melanization incorporates multiple catecholamines to produce polytypic melanin. J. Biol. Chem. 2021, 298, 101519. [Google Scholar] [CrossRef]
- Frases, S.; Salazar, A.; Dadachova, E.; Casadevall, A. Cryptococcus neoformans Can Utilize the Bacterial Melanin Precursor Homogentisic Acid for Fungal Melanogenesis. Appl. Environ. Microbiol. 2007, 73, 615–621. [Google Scholar] [CrossRef]
- Perez-Dulzaides, R.; Camacho, E.; Cordero, R.J.B.; Casadevall, A. Cell-wall dyes interfere with Cryptococcus neoformans melanin deposition. Microbiology 2018, 164, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; Chrissian, C.; Cordero, R.J.B.; Liporagi-Lopes, L.; Stark, R.E.; Casadevall, A. N-acetylglucosamine affects Cryptococcus neoformans cell-wall composition and melanin architecture. Microbiology 2017, 163, 1540–1556. [Google Scholar] [CrossRef] [PubMed]
- Herth, W. Calcofluor white and Congo red inhibit chitin microfibril assembly of Poterioochromonas: Evidence for a gap between polymerization and microfibril formation. J. Cell Biol. 1980, 87, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Elorza, M.V.; Rico, H.; Sentandreu, R. Calcofluor White Alters the Assembly of Chitin Fibrils in Saccharomyces cerevisiae and Candida albicans Cells. Microbiology 1983, 129, 1577–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raut, J.; Rathod, V.; Karuppayil, S.M. Cell Surface Hydrophobicity and Adhesion: A Study on Fifty Clinical Isolates of Candida albicans. Nippon Ishinkin Gakkai Zasshi 2010, 51, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danchik, C.; Casadevall, A. Role of Cell Surface Hydrophobicity in the Pathogenesis of Medically-Significant Fungi. Front. Cell. Infect. Microbiol. 2021, 10, 594973. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Aisen, P.; Casadevall, A. Melanin, melanin “ghosts,” and melanin composition in Cryptococcus neoformans. Infect. Immun. 1996, 64, 2420–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidrich, D.; Corbellini, V.A.; Mendes, S.D.C.; Fernandes, E.K.; Lazzarotto, L.; Ribeiro, A.C.; Zanette, R.A.; Scroferneker, M.L. Melanin: Quantification and protection against oxidative stress in chromoblastomycosis agents. Med. Mycol. 2018, 57, 260–263. [Google Scholar] [CrossRef]
- Wolf, J.M.; Rivera, J.; Casadevall, A. Serum albumin disrupts Cryptococcus neoformans and Bacillus anthracis extracellular vesicles. Cell. Microbiol. 2012, 14, 762–773. [Google Scholar] [CrossRef]
- Coelho, C.; Vij, R.; Smith, D.Q.; Brady, N.R.; Hamacher-Brady, A.; Casadevall, A. Study of Microbial Extracellular Vesicles:Separation by Density Gradients, Protection Assays and Labelling for Live Tracking. Bio Protoc. 2020, 10, e3502. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Brown, L.C.; Maryam, M.; Vij, R.; Smith, D.F.; Burnet, M.C.; Kyle, J.E.; Heyman, H.M.; Ramirez, J.; Prados-Rosales, R.; et al. Listeria monocytogenes virulence factors, including listeriolysin O, are secreted in biologically active extracellular vesicles. J. Biol. Chem. 2019, 294, 1202–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Inglett, G.E. Molecular weight and ionic strength dependence of fluorescence intensity of the Calcofluor/β-glucan complex in flow-injection analysis. J. Food Compos. Anal. 2006, 19, 466–472. [Google Scholar] [CrossRef]
- Wu, J.; Deng, X.; Tian, B.; Wang, L.; Xie, B. Interactions between Oat β-Glucan and Calcofluor Characterized by Spectroscopic Method. J. Agric. Food Chem. 2008, 56, 1131–1137. [Google Scholar] [CrossRef]
- Vylkova, S.; Carman, A.J.; Danhof, H.A.; Collette, J.R.; Zhou, H.; Lorenz, M.C. The Fungal Pathogen Candida albicans Autoinduces Hyphal Morphogenesis by Raising Extracellular pH. mBio 2011, 2, e00055-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadorojny, C.; Saxton, S.; Finger, R. Spectrophotometric Determination of Ammonia. J. (Water Pollut. Control. Fed.) 1973, 45, 905–912. [Google Scholar]
- Pye, A.E. Activation of prophenoloxidase and inhibition of melanization in the haemolymph of immune Galleria mellonella larvae. Insect Biochem. 1978, 8, 117–123. [Google Scholar] [CrossRef]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Et Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Ezzati-Tabizi, R.; Talaei-Hassanloui, R.; Farrokhi, N.; Hossininaveh, V.; Alavi, M. Haemolymph phenoloxidase activity of larval Plodia interpunctella and Galleria mellonella in response to Beauveria bassiana and Pseudomonas fluorescens. Int. J. Agric. Innov. Res. 2013, 2, 217–220. [Google Scholar]
- Trevijano-Contador, N.; Herrero-Fernández, I.; García-Barbazán, I.; Scorzoni, L.; Rueda, C.; Rossi, S.A.; Zaragoza, O. Cryptococcus neoformans induces antimicrobial responses and behaves as a facultative intracellular pathogen in the non mammalian model Galleria mellonella. Virulence 2015, 6, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Bustos, V.; Pemán, J.; Ruiz-Gaitán, A.; Cabañero-Navalon, M.D.; Cabanilles-Boronat, A.; Fernández-Calduch, M.; Ruiz-Saurí, A. Host–pathogen interactions upon Candida auris infection: Fungal behaviour and immune response in Galleria mellonella. Emerg. Microbes Infect. 2022, 11, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bustos, V.; Ruiz-Saurí, A.; Ruiz-Gaitán, A.; Sigona-Giangreco, I.A.; Cabañero-Navalon, M.D.; Sabalza-Baztán, O.; Pemán, J. Characterization of the Differential Pathogenicity of Candida auris in a Galleria mellonella Infection Model. Microbiol. Spectrum. 2021, 9, e00013-21. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Youngchim, S.; Zamith-Miranda, D.; Nosanchuk, J. Fungal Melanin and the Mammalian Immune System. J. Fungi 2021, 7, 264. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.; Emery, H. Temperature regulation of the cryptococcal phenoloxidase. J. Med. Vet. Mycol. 1991, 29, 121–124. [Google Scholar] [CrossRef]
- Albuquerque, P.; Nicola, A.M.; Nieves, E.; Paes, H.C.; Williamson, P.R.; Silva-Pereira, I.; Casadevall, A. Quorum Sensing-Mediated, Cell Density-Dependent Regulation of Growth and Virulence in Cryptococcus neoformans. mBio 2014, 5, e00986-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, L.R.; Ntiamoah, P.; Gácser, A.; Casadevall, A.; Nosanchuk, J.D. Voriconazole Inhibits Melanization in Cryptococcus neoformans. Antimicrob. Agents Chemother. 2007, 51, 4396–4400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, S.D.; Bennett, J.E.; Kwon-Chung, K.J.; Perfect, J.R.; Williamson, P.R. Effect of the laccase gene CNLAC1, on virulence of Cryptococcus neoformans. J. Exp. Med. 1996, 184, 377–386. [Google Scholar] [CrossRef]
- Nakayasu, E.S.; Nosanchuk, J.D. Multi-omics Signature of Candida auris, an Emerging and Multidrug-Resistant Pathogen. mSystems 2019, 4, e00257-19. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.; Chow, F.W.-N.; Tsang, C.-C.; Liu, X.; Yao, W.; Chan, T.T.-Y.; Siu, G.K.-H.; Ho, A.Y.-M.; Luk, K.S.; Lau, S.K.-P.; et al. Induction of amphotericin B resistance in susceptible Candida auris by extracellular vesicles. Emerg. Microbes Infect. 2022, 11, 1900–1909. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Frases, S.; Nicola, A.M.; Rodrigues, M.L.; Casadevall, A. Vesicle-associated melanization in Cryptococcus neoformans. Microbiology 2009, 155, 3860–3867. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.S.; Coelho, C.; De Leon-Rodriguez, C.M.; Rossi, D.C.P.; Camacho, E.; Jung, E.H.; Kulkarni, M.; Casadevall, A. Cryptococcus neoformans urease affects the outcome of intracellular pathogenesis by modulating phagolysosomal pH. PLoS Pathog. 2018, 14, e1007144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vylkova, S.; Lorenz, M.C. Modulation of Phagosomal pH by Candida albicans Promotes Hyphal Morphogenesis and Requires Stp2p, a Regulator of Amino Acid Transport. PLoS Pathog. 2014, 10, e1003995. [Google Scholar] [CrossRef] [PubMed]
- Meena, B.; Rajan, L.A.; Vinithkumar, N.V.; Kirubagaran, R. Novel marine actinobacteria from emerald Andaman & Nicobar Islands: A prospective source for industrial and pharmaceutical byproducts. BMC Microbiol. 2013, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Singleton, D.R.; Fidel, P.L., Jr.; Wozniak, K.L.; Hazen, K.C. Contribution of cell surface hydrophobicity protein 1 (Csh1p) to virulence of hydrophobic Candida albicans serotype A cells. FEMS Microbiol. Lett. 2005, 244, 373–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazen, K.C.; Brawner, D.L.; Riesselman, M.H.; Jutila, M.A.; Cutler, J.E. Differential adherence of hydrophobic and hydrophilic Candida albicans yeast cells to mouse tissues. Infect. Immun. 1991, 59, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, Y.; Kagaya, K. Molecular bases of adhesion of Candida albicans. J. Med. Vet. Mycol. 1997, 35, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Tronchin, G.; Pihet, M.; Bezerra, L.M.L.; Bouchara, J.-P. Adherence mechanisms in human pathogenic fungi. Med. Mycol. 2008, 46, 749–772. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Finn, T. Multidrug-Resistant Candida haemulonii and C. auris, Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23, 195–203. [Google Scholar] [CrossRef]
- Rossato, L.; Colombo, A.L. Candida auris: What Have We Learned About Its Mechanisms of Pathogenicity? Front. Microbiol. 2018, 9, 3081. [Google Scholar] [CrossRef]
- Bravo Ruiz, G.; Lorenz, A. What do we know about the biology of the emerging fungal pathogen of humans Candida auris? Microbiol. Res. 2021, 242, 126621. [Google Scholar] [CrossRef] [PubMed]
- Mattoon, E.; Cordero, R.; Casadevall, A. Fungal Melanins and Applications in Healthcare, Bioremediation and Industry. J. Fungi 2021, 7, 488. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Casadevall, A. Impact of Melanin on Microbial Virulence and Clinical Resistance to Antimicrobial Compounds. Antimicrob. Agents Chemother. 2006, 50, 3519–3528. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Jain, K.; Wang, Y.; Pawar, K.; Kaur, H.; Sharma, K.K.; Tripathy, V.; Singh, A.; Xu, J.; Chowdhary, A. Candida auris on Apples: Diversity and Clinical Significance. mBio 2022, 13, e00518-22. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Alias | Clade | Melanin |
|---|---|---|---|
| CDC 381 | B11220 | Clade II (East Asian) | No |
| CDC 382 | B11109 | Clade I (South Asian) | Partially |
| CDC 383 | B11221 | Clade III (African) | No |
| CDC 384 | B11222 | Clade III (African) | No |
| CDC 385 | B11244 | Clade IV (South American) | Yes |
| CDC 386 | B11245 | Clade IV (South American) | Yes |
| CDC 387 | B8441 | Clade I (South Asian) | Yes |
| CDC 388 | B11098 | Clade I (South Asian) | Yes |
| CDC 389 | B11203 | Clade I (South Asian) | Yes |
| CDC 390 | B11205 | Clade I (South Asian) | Yes |
| CDC 931 | B11243 | Clade IV (South American) | Yes |
| CDC 1097 | IFRC2087 | Clade V (Iranian) | Yes |
| CDC 1101 | B18678 | Clade II (East Asian) | No |
| CDC 1102 | B17835 | Clade III (African) | No |
| CDC 1103 | B18683 | Clade III (African) | No |
| CDC 1104 | B18017 | Clade IV (South American) | Yes |
| MMC1 | N/A | Clade I (South Asian) | Yes |
| MMC2 | N/A | Unknown | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, D.F.Q.; Mudrak, N.J.; Zamith-Miranda, D.; Honorato, L.; Nimrichter, L.; Chrissian, C.; Smith, B.; Gerfen, G.; Stark, R.E.; Nosanchuk, J.D.; et al. Melanization of Candida auris Is Associated with Alteration of Extracellular pH. J. Fungi 2022, 8, 1068. https://doi.org/10.3390/jof8101068
Smith DFQ, Mudrak NJ, Zamith-Miranda D, Honorato L, Nimrichter L, Chrissian C, Smith B, Gerfen G, Stark RE, Nosanchuk JD, et al. Melanization of Candida auris Is Associated with Alteration of Extracellular pH. Journal of Fungi. 2022; 8(10):1068. https://doi.org/10.3390/jof8101068
Chicago/Turabian StyleSmith, Daniel F. Q., Nathan J. Mudrak, Daniel Zamith-Miranda, Leandro Honorato, Leonardo Nimrichter, Christine Chrissian, Barbara Smith, Gary Gerfen, Ruth E. Stark, Joshua D. Nosanchuk, and et al. 2022. "Melanization of Candida auris Is Associated with Alteration of Extracellular pH" Journal of Fungi 8, no. 10: 1068. https://doi.org/10.3390/jof8101068
APA StyleSmith, D. F. Q., Mudrak, N. J., Zamith-Miranda, D., Honorato, L., Nimrichter, L., Chrissian, C., Smith, B., Gerfen, G., Stark, R. E., Nosanchuk, J. D., & Casadevall, A. (2022). Melanization of Candida auris Is Associated with Alteration of Extracellular pH. Journal of Fungi, 8(10), 1068. https://doi.org/10.3390/jof8101068