
New Species of Nectriaceae (Hypocreales) from China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Morphological Studies
2.2. DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analyses
3. Results
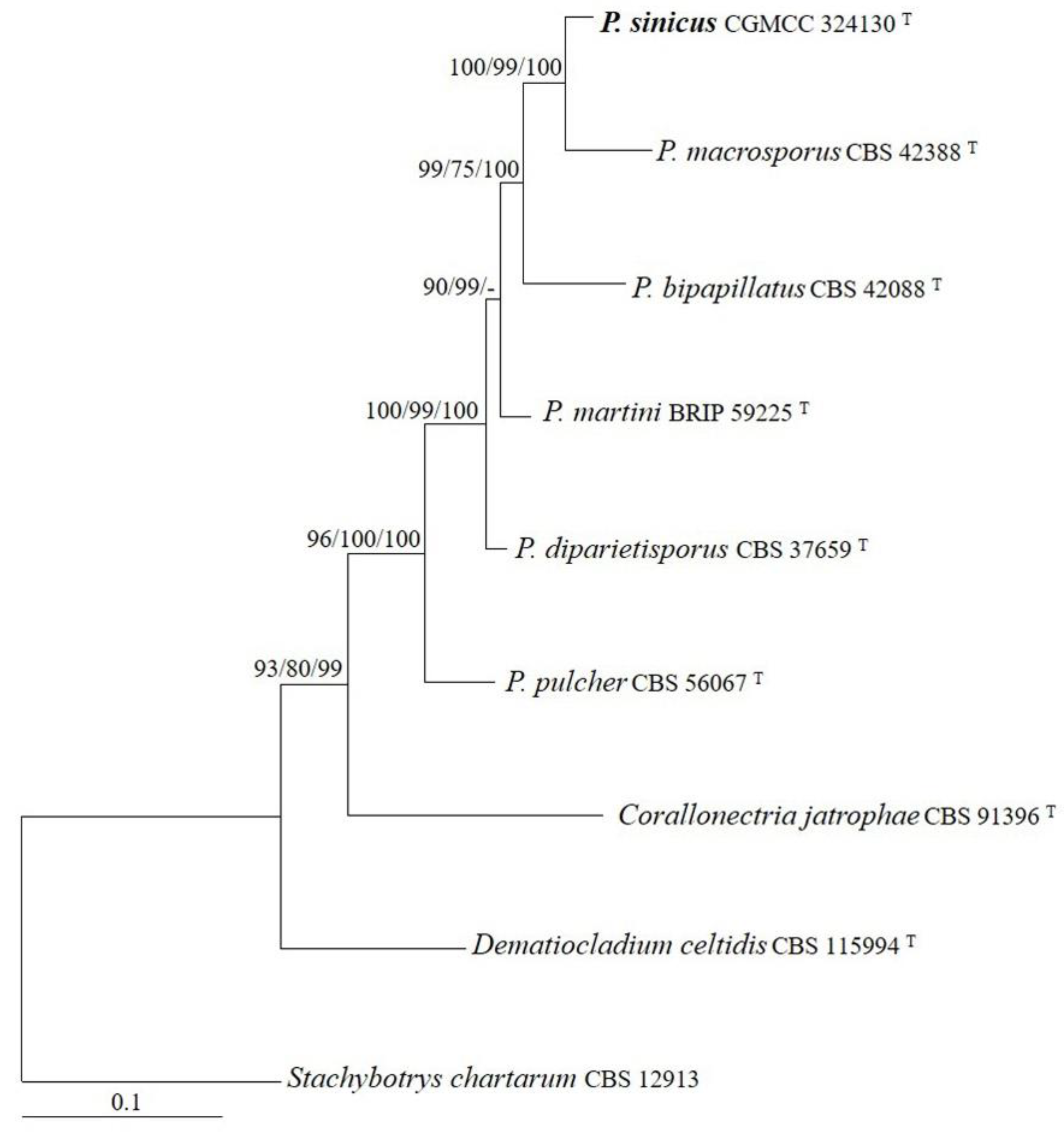
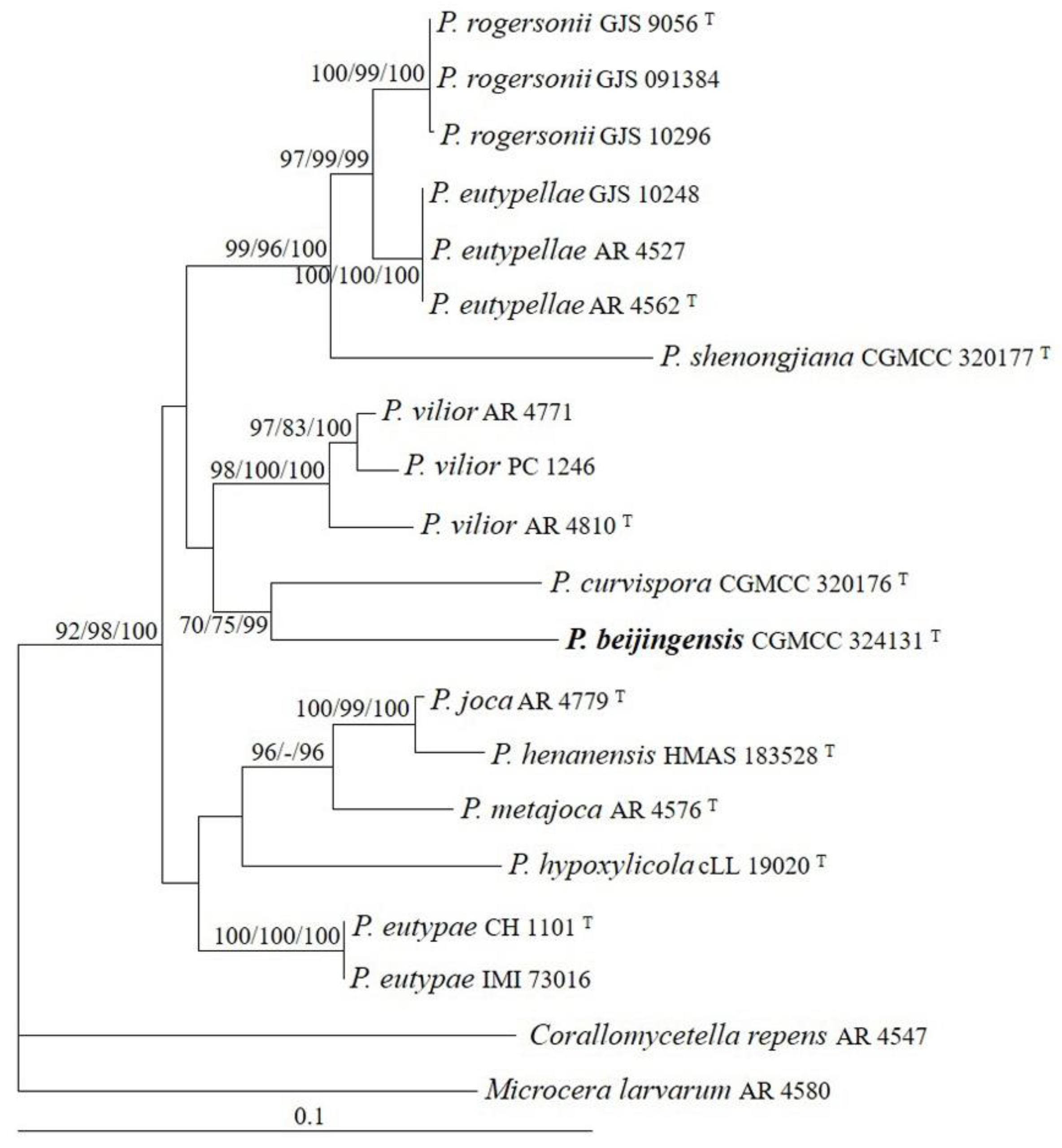
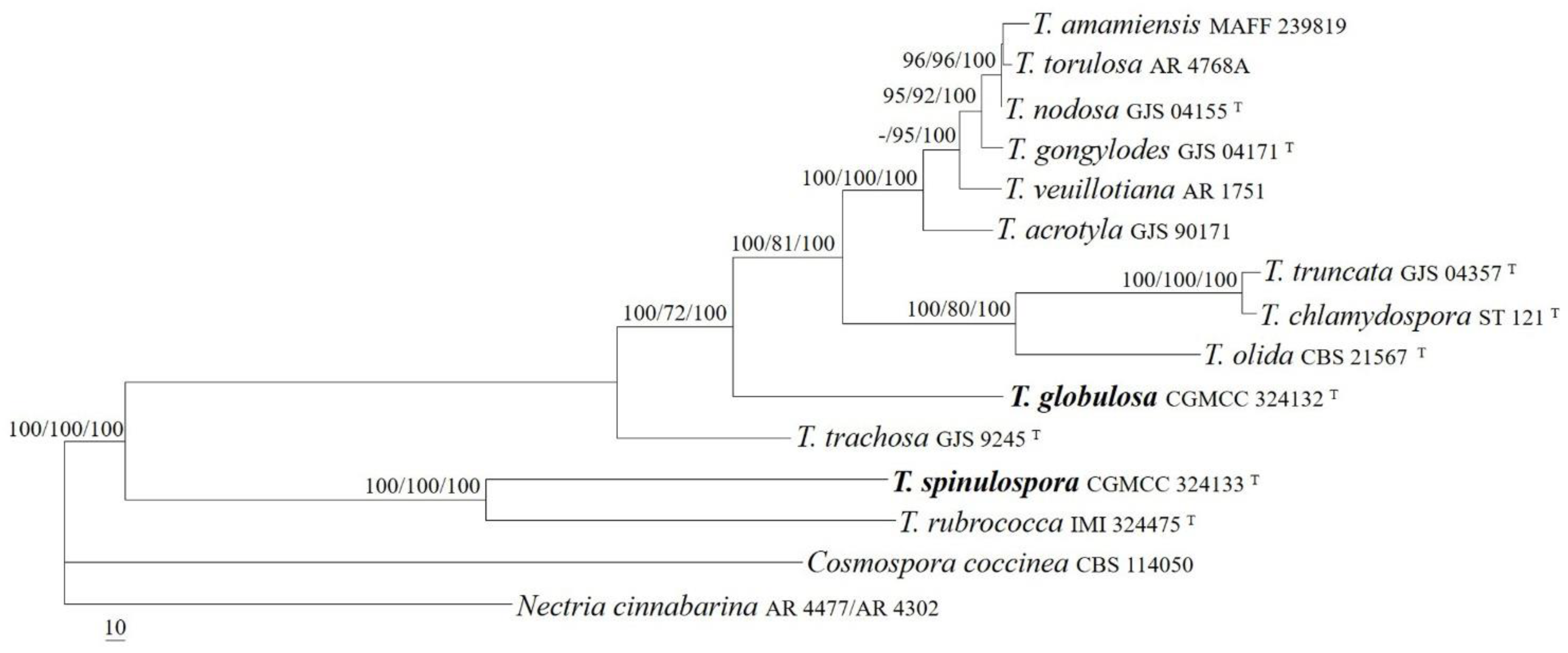
3.1. Phylogeny
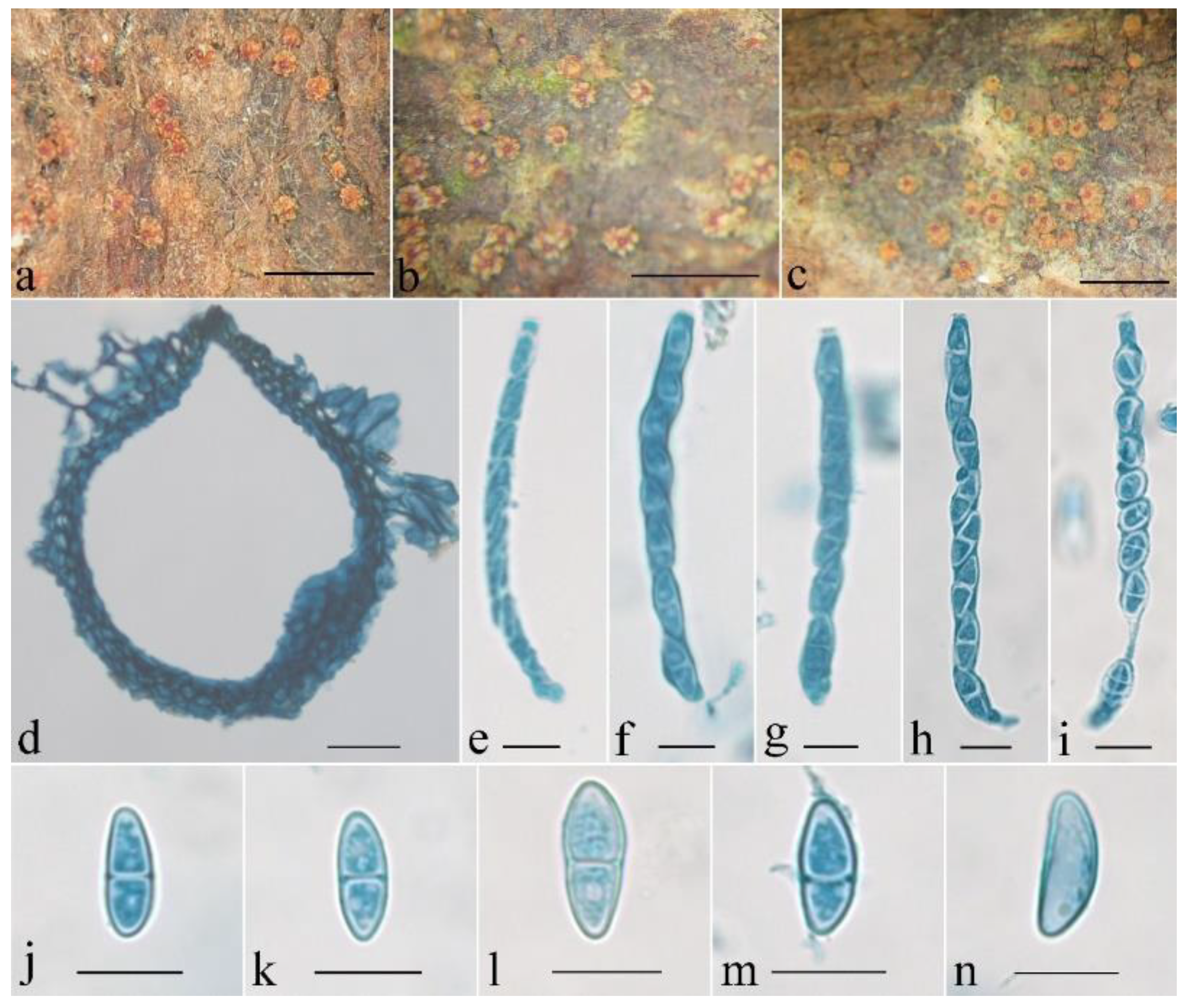
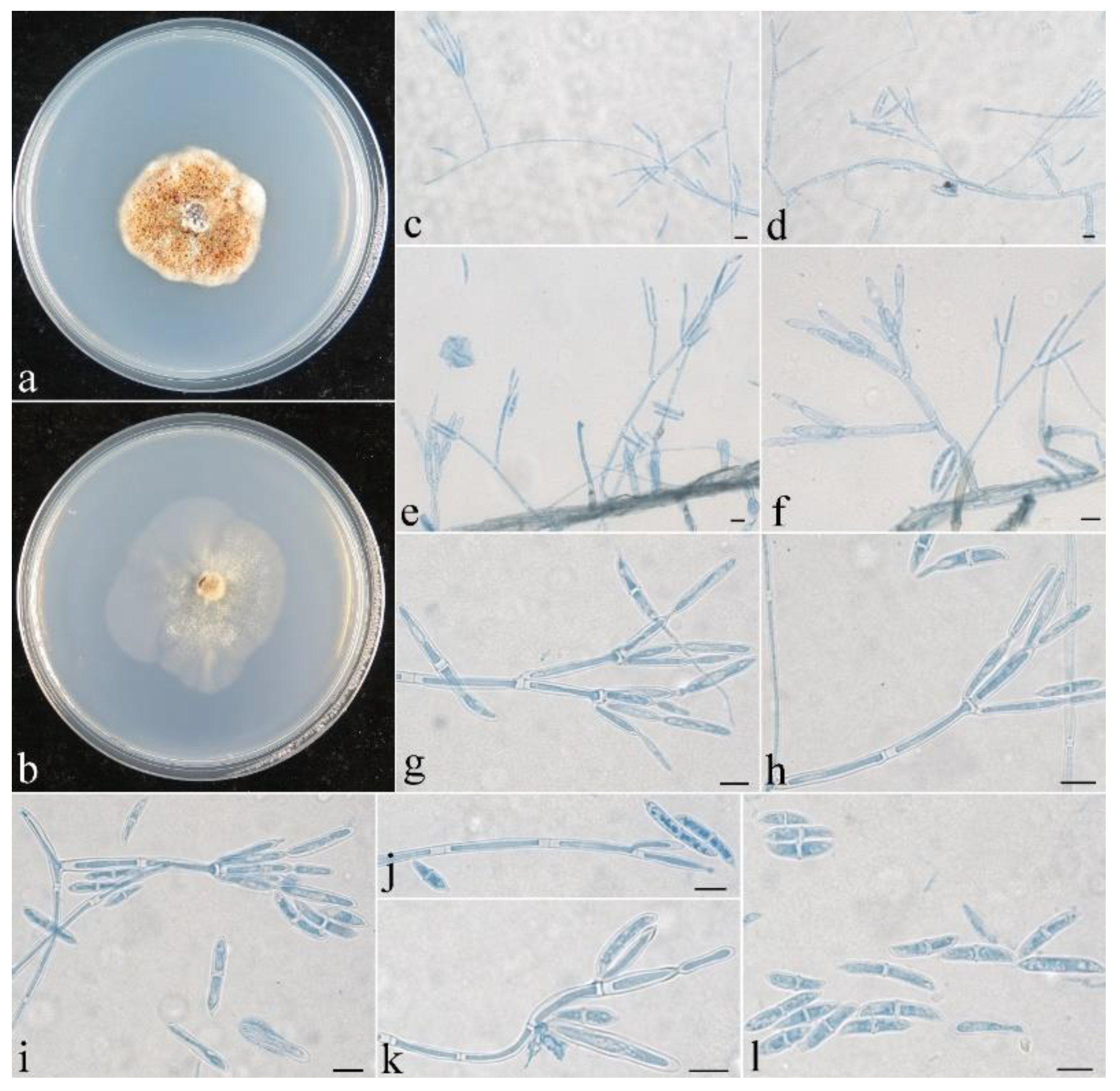
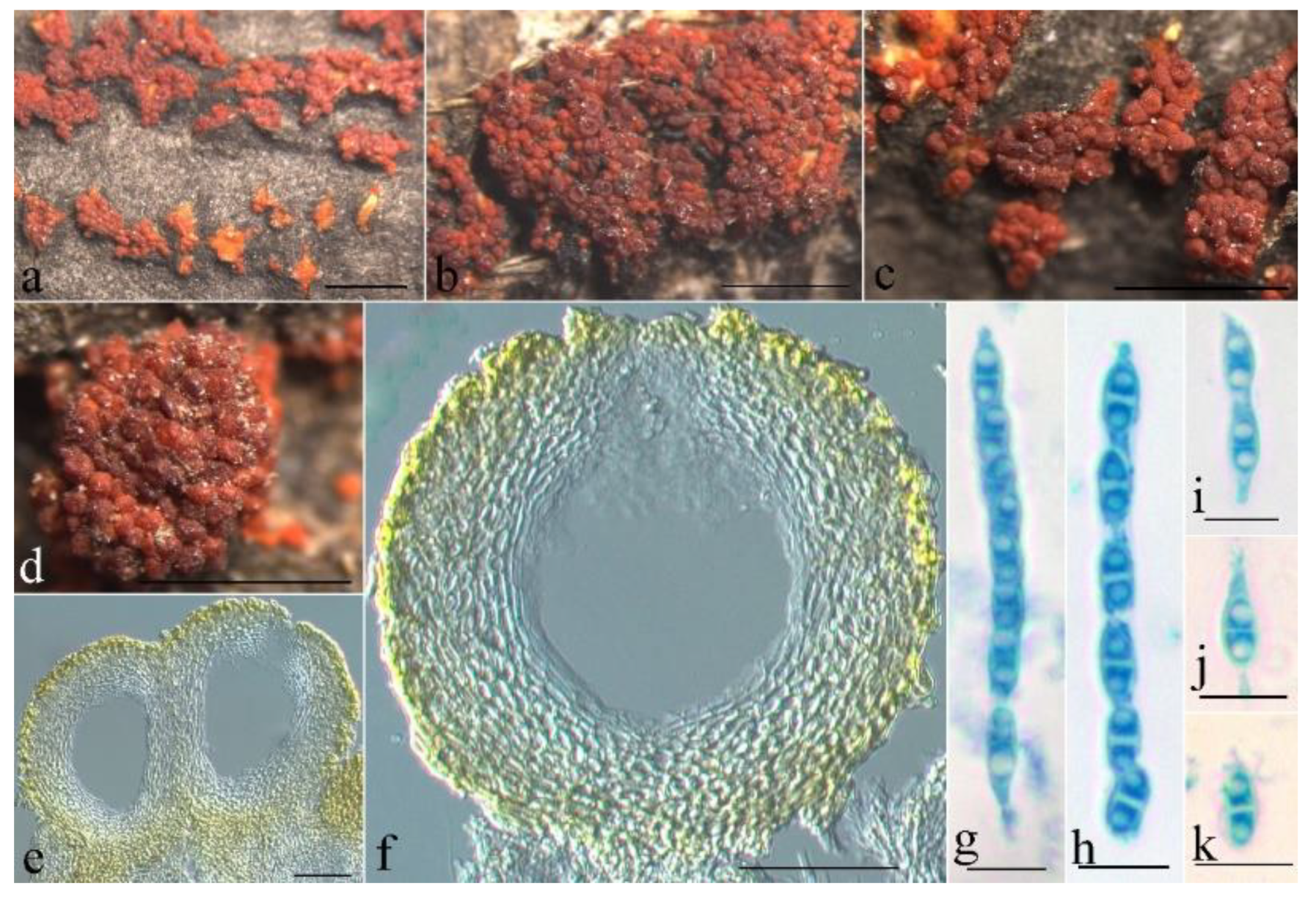
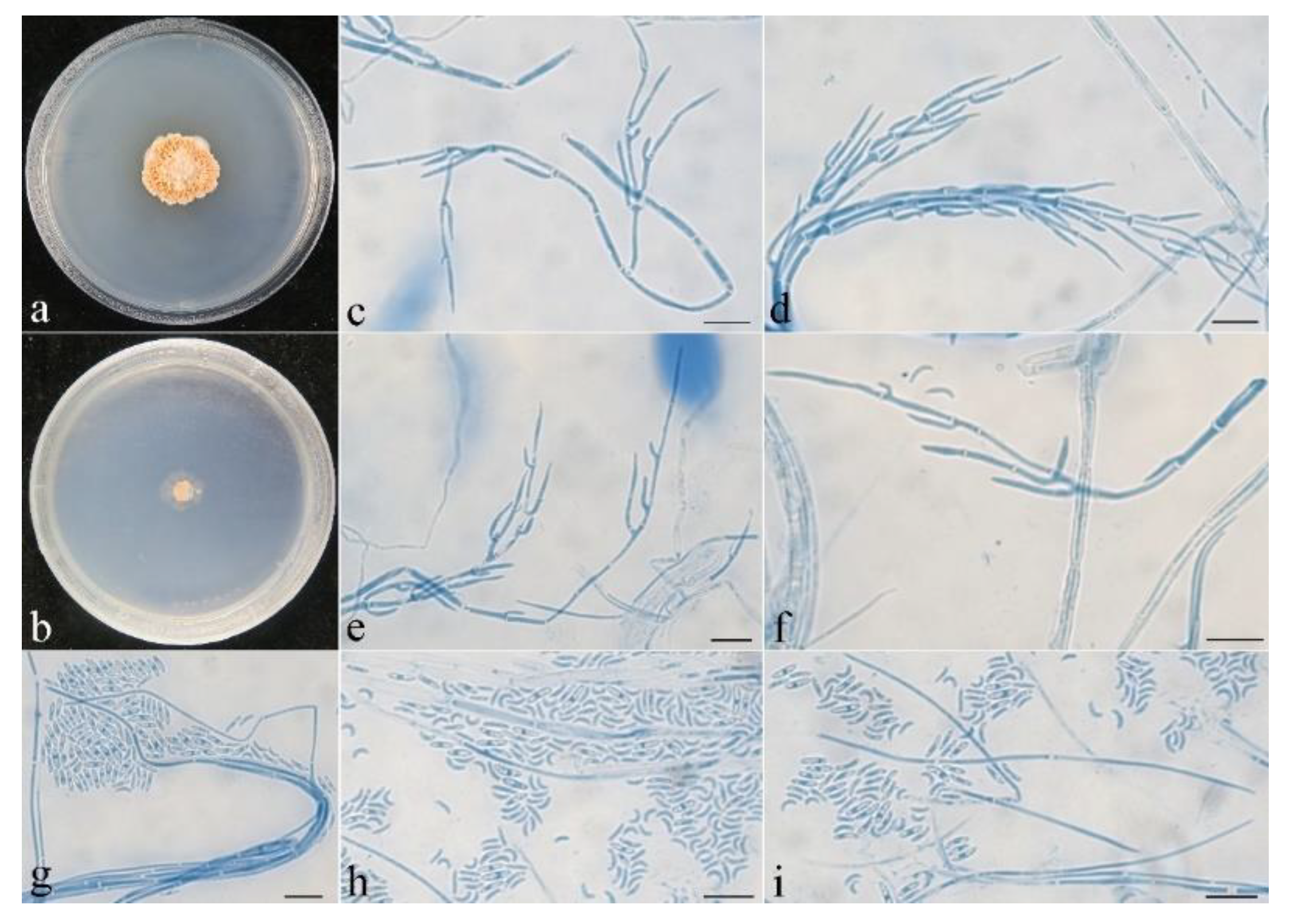
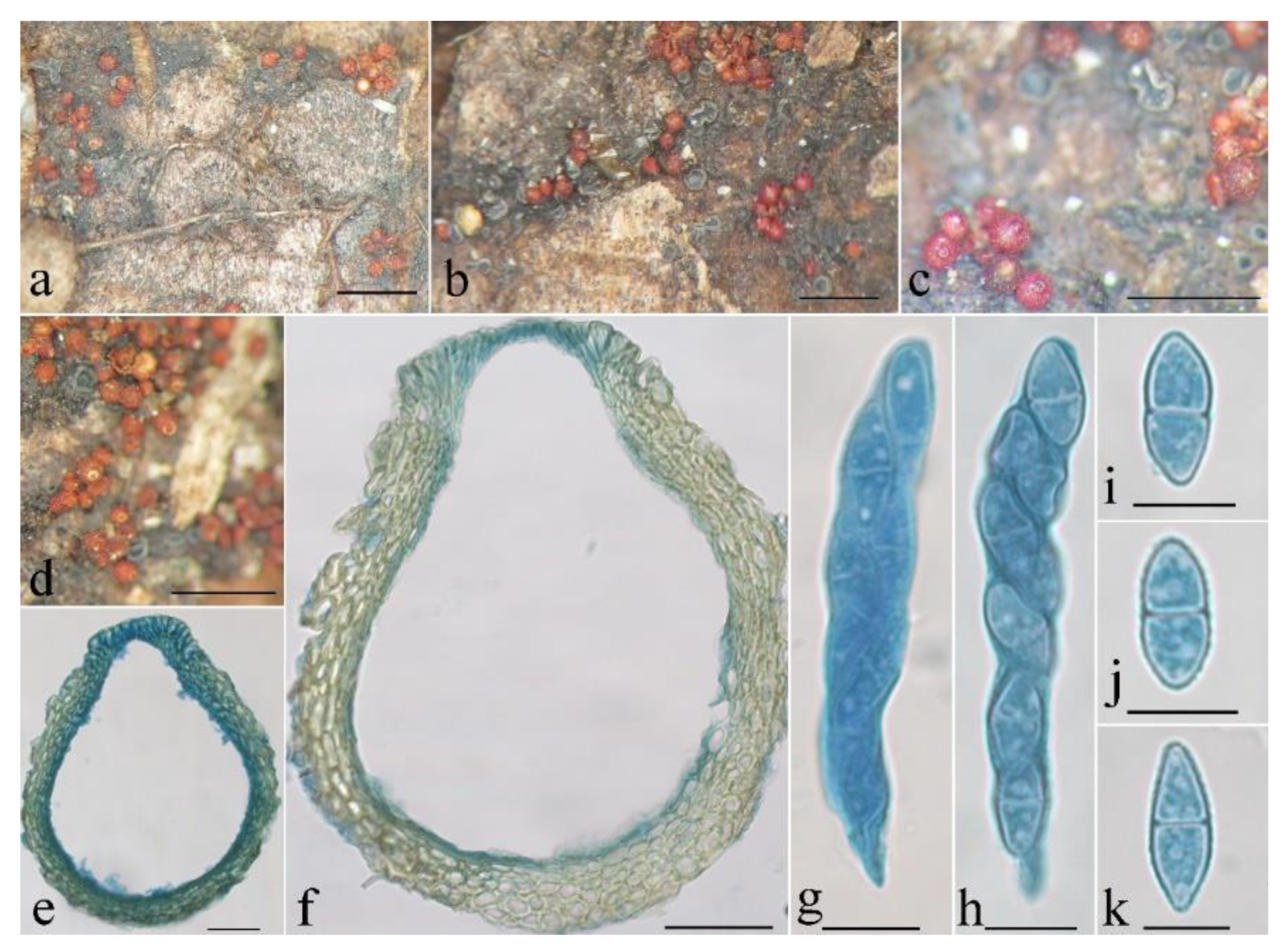
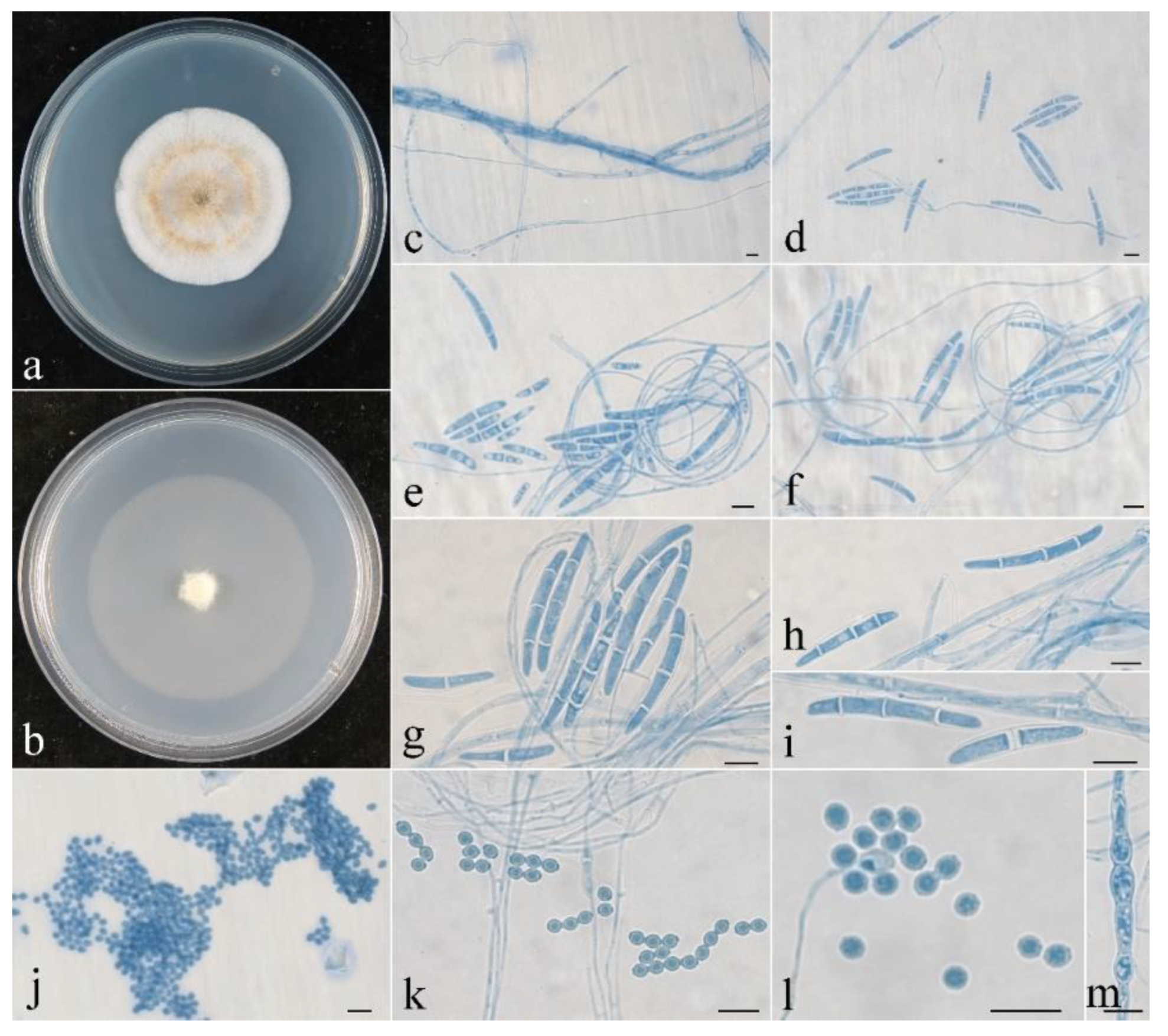
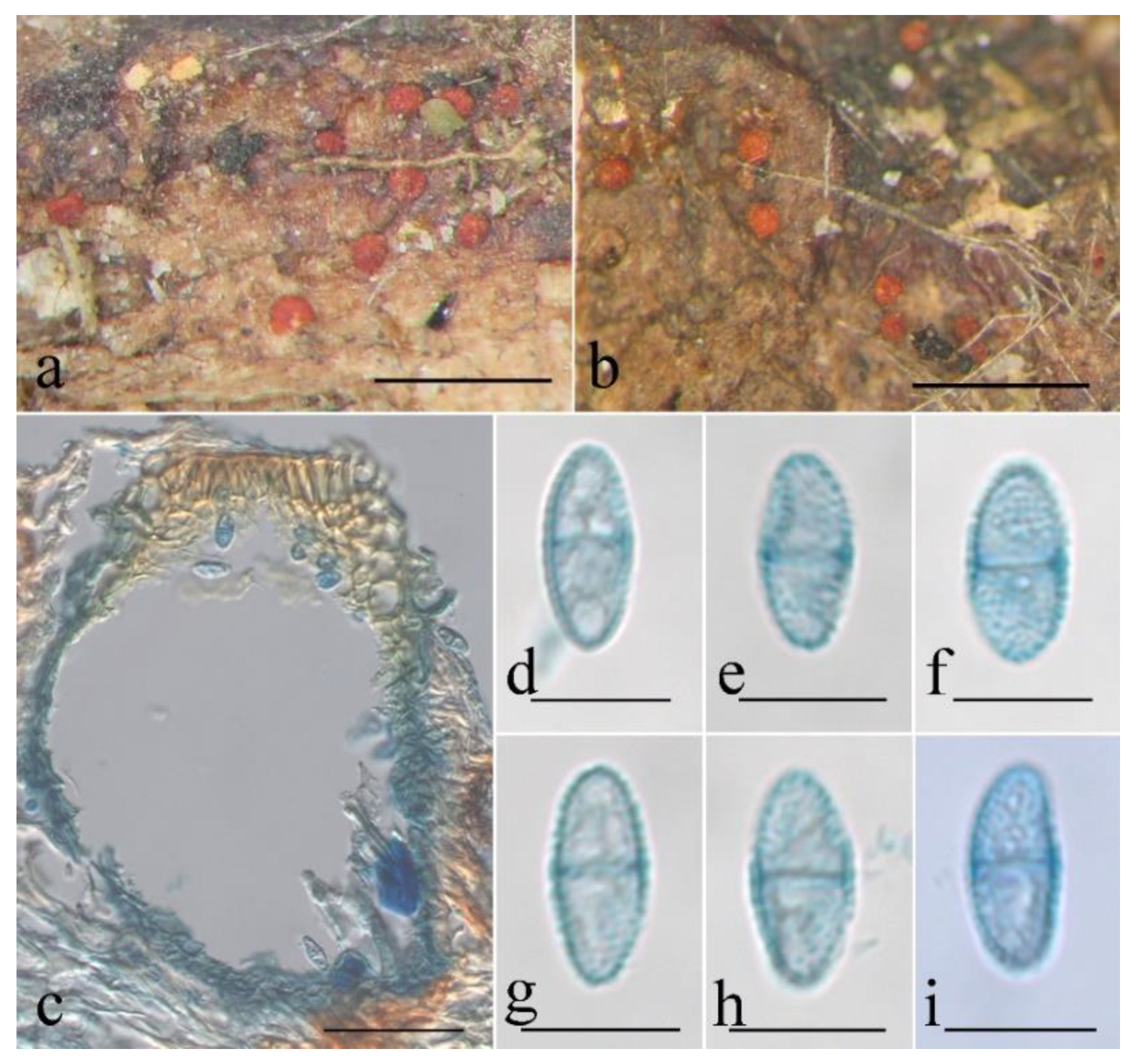
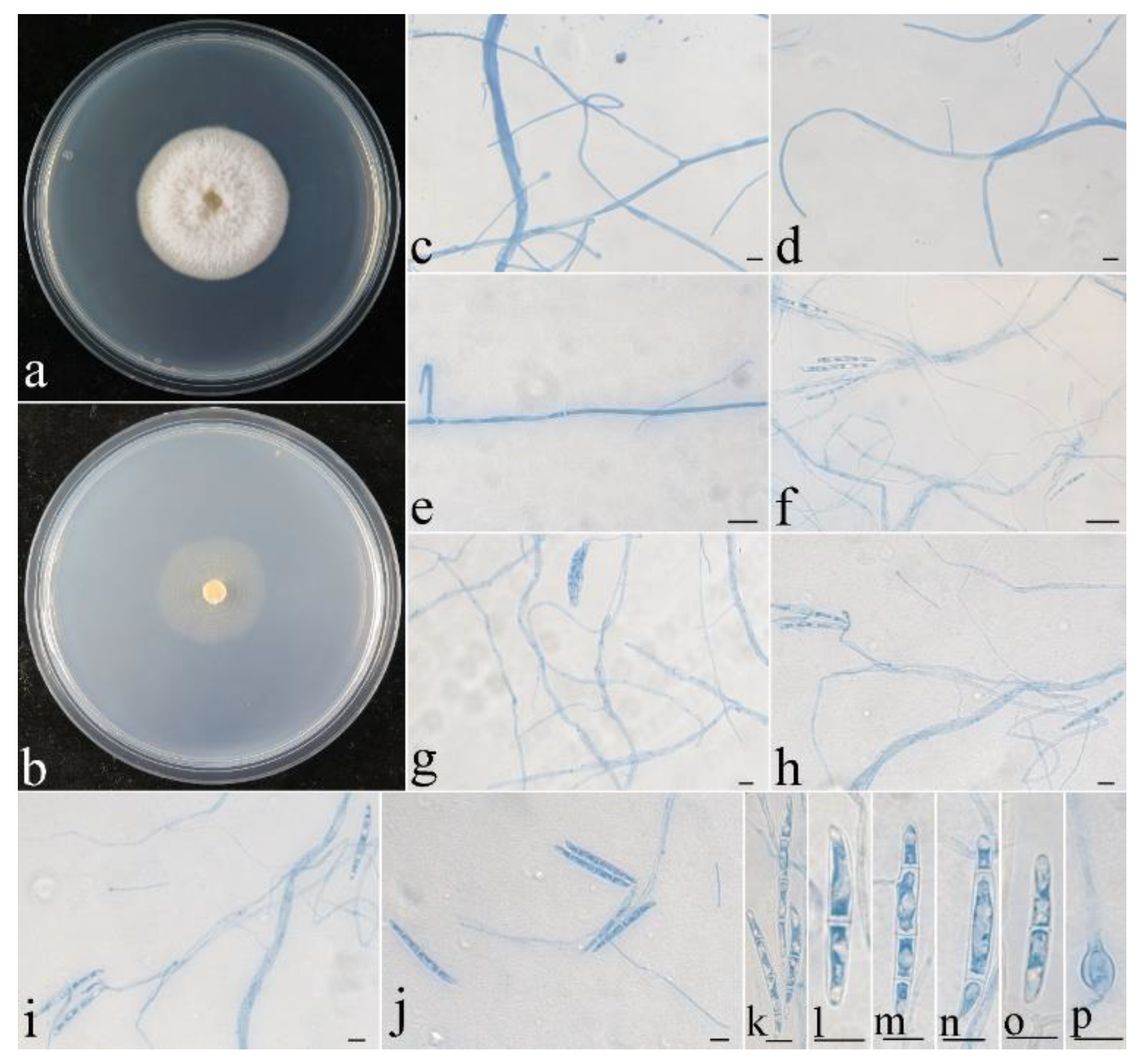
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–260. [Google Scholar]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- van Emden, J.H. Penicillifer, a new genus of Hyphomycetes from soil. Acta Bot. Neerl. 1968, 17, 54–58. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.S.; Rossman, A.Y.; Samuels, G.J.; Chaverri, P. Pseudocosmospora, a new genus to accommodate Cosmospora vilior and related species. Mycologia 2013, 105, 1287–1305. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.Q.; Zhuang, W.Y. Eight new combinations of Bionectriaceae and Nectriaceae. Mycosystema 2017, 36, 278–281. [Google Scholar]
- Zeng, Z.Q.; Zhuang, W.Y. Our current understanding of the genus Pseudocosmospora (Hypocreales, Nectriaceae) in China. Mycol. Prog. 2021, 20, 419–429. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J. Pseudocosmospora hypoxylicola (Nectriaceae), a new species from the French Alps. Ascomycete.org 2020, 12, 6–10. [Google Scholar]
- Chaverri, P.; Salgado, C.; Hirooka, Y.; Rossman, A.Y.; Samuels, G.J. Delimitation of Neonectria and Cylindrocarpon (Nectriaceae, Hypocreales, Ascomycota) and related genera with Cylindrocarpon-like anamorphs. Stud. Mycol. 2011, 68, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Salazar, C.; Rossman, A.Y.; Chaverri, P. The genus Thelonectria (Nectriaceae, Hypocreales, Ascomycota) and closely related species with Cylindrocarpon-like asexual states. Fungal Divers. 2016, 80, 411–455. [Google Scholar] [CrossRef]
- Salgado-Salazar, C.; Rossman, A.; Samuels, G.J.; Capdet, M.; Chaverri, P. Multigene phylogenetic analyses of the Thelonectria coronata and T. veuillotiana species complexes. Mycologia 2012, 104, 1325–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado-Salazar, C.; Rossman, A.Y.; Samuels, G.J.; Hirooka, Y.; Sanchez, R.M.; Chaverri, P. Phylogeny and taxonomic revision of Thelonectria discophora (Ascomycota, Hypocreales, Nectriaceae) species complex. Fungal Divers. 2015, 70, 1–29. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Four new taxa of Ilyonectria and Thelonectria (Nectriaceae) revealed by morphology and combined ITS and β-tubulin sequence data. Phytotaxa 2013, 85, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.Q.; Zhuang, W.Y. The genera Rugonectria and Thelonectria (Hypocreales, Nectriaceae) in China. MycoKeys 2019, 55, 101–120. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Nouri, M.T.; Trouillas, F.P. Taxonomy and multi-locus phylogeny of cylindrocarpon-like species associated with diseased roots of grapevine and other fruit and nut crops in California. Fungal Syst. Evol. 2019, 4, 59–75. [Google Scholar] [CrossRef]
- Braun, U.; Bensch, K. Annotated list of taxonomic novelties published in “Fungi Rhenani Exsiccati” Supplementi Fasc. 1 to 5, issued by K. W. G. L. Fuckel between 1865 and 1867. Schlechtendalia 2020, 37, 80–133. [Google Scholar]
- Nirenberg, H.I. Studies on the morphologic and biologic differentiation in Fusarium section Liseola. Mitt. Biol. Bundesanst. Land-Forstw. 1976, 169, 1–117. [Google Scholar]
- Samuels, G.J.; Dodd, S.; Lu, B.S.; Petrini, O.; Schroers, H.J.; Druzhinina, I.S. The Trichoderma koningii aggregate species. Stud. Mycol. 2006, 56, 67–133. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analyzed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA Polymerase II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgin, D.G. The ClustalX windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4883. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP 4.0b10: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Mol. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comp. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Brayford, D.; Samuels, G.J. Some didymosporous species of Nectria with nonmicroconidial Cylindrocarpon anamorphs. Mycologia 1993, 85, 612–637. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code); Koeltz Scientific Books: Berlin/Heidelberg, Germany, 2018; pp. 1–254. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs No 4; Published by the author; Kobe, Japan, 1985; pp. 1–68. [Google Scholar]
- Zhang, X.M.; Zhuang, W.Y. Phylogeny of some genera in the Nectriaceae (Hypocreales, Ascomycetes) inferred from 28S nrDNA partial sequences. Mycosystema 2006, 25, 15–22. [Google Scholar]
- Luo, J.; Zhuang, W.Y. Chaetopsinectria (Nectriaceae, Hypocreales), a new genus with Chaetopsina anamorphs. Mycologia 2010, 102, 976–984. [Google Scholar] [CrossRef]
- Samuels, G.J.; Doi, Y.; Rogerson, C.T. Contributions toward a mycobiota of Indonesia: Hypocreales. Mem. N. Y. Bot. Gard. 1990, 59, 6–108. [Google Scholar]
- Samuels, G.J.; Brayford, D. Species of Nectria (sensu lato) with red perithecia and striate ascospores. Sydowia 1994, 46, 75–161. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| ITS | LSU | rpb2 | tef1 | ||
| Corallonectria jatrophae | CBS 91396 T | NR153873 | KM231611 | KM232298 | KM231863 |
| Dematiocladium celtidis | CBS 115994 T | AY793430 | AY793438 | - | KM231864 |
| P. bipapillatus | CBS 42088 T | KM231740 | KM231608 | KM232295 | KM231860 |
| P. diparietisporus | CBS 37659 T | NR154310 | MH869437 | KM232296 | KM231861 |
| P. macrosporus | CBS 42388 T | MH862133 | KM231607 | KM232294 | KM231859 |
| P. martinii | BRIP 59225 T | NR168155 | NG068753 | - | KJ869241 |
| P. pulcher | CBS 56067 T | NR154311 | NG058093 | KM232297 | KM231862 |
| P. sinicus | CGMCC 3.24130 T | OP223439 a | OP223435 | OP272863 | OP272864 |
| Stachybotrys chartarum | CBS 12913 | MH854622 | MH866145 | KM232434 | KM231994 |
| Species | Herbarium/Strain Number | GenBank Accession Numbers | ||
|---|---|---|---|---|
| ITS | LSU | tub2 | ||
| Corallomycetella repens | AR 4547 | JF832594 | JF832679 | JF832838 |
| Microcera larvarum | AR 4580 | KC291751 | KC291759 | KC291935 |
| P. beijingensis | CGMCC 3.24131 T | OP223438 a | OP223434 | OP272862 |
| P. curvispora | CGMCC 3.20176 T | MT592897 | MT592879 | MT606156 |
| P. eutypae | CH 1101 T | KC291735 | KC291766 | KC291925 |
| IMI 73016 | KC291736 | KC291786 | - | |
| AR 4527 | KC291720 | KC291756 | KC291909 | |
| P. eutypellae | AR 4562 T | KC291721 | KC291757 | KC291912 |
| GJS 10248 | KC291722 | KC291772 | KC291911 | |
| P. henanensis | HMAS 183528 T | GU075856 | GU075863 | HM054103 |
| P. hypoxylicola | cLL 19020 T | MN886606 | MN886608 | - |
| P. joca | AR 4779 T | KC291746 | KC291762 | KC291924 |
| P. metajoca | AR 4576 T | KC291745 | KC291758 | KC291923 |
| P. rogersonii | GJS 9056 T | KC291729 | KC291780 | KC291915 |
| GJS 10296 | KC291727 | KC291774 | KC291917 | |
| GJS 091384 | KC291726 | KC291770 | KC291914 | |
| P. shennongjiana | CGMCC 3.20177 T | MT592898 | MT592880 | MT606157 |
| P. vilior | AR 4810 T | KC291737 | KC291763 | KC291928 |
| AR 4771 | KC291734 | KC291761 | KC291926 | |
| PC 1246 | KC291738 | KC291791 | KC291927 | |
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| act | ITS | LSU | rpb1 | tub2 | ||
| Cosmospora coccinea | CBS 114050 | GQ505967 | FJ474072 | GQ505990 | GQ506020 | DQ522501 |
| Nectria cinnabarina | AR 4477/AR 4302 | HM484627 | HM484548 | HM484562 | M484577 | HM484820 |
| T. acrotyla | GJS 90171 | JQ365047 | JQ403329 | JQ403368 | JQ403407 | JQ394720 |
| T. amamiensis | MAFF 239819 | JQ365054 | JQ403337 | JQ403375 | KJ022408 | JQ394727 |
| T. chlamydospora | ST 121 T | LC519560 | LC509450 | LC509452 | - | - |
| T. globulosa | CGMCC 3.24132 T | OP272865 a | OP223436 | OP223432 | OP272867 | OP586762 |
| T. gongylodes | GJS 04171 T | JQ365038 | JQ4033 | JQ403358 | JQ403395 | JQ394710 |
| T. nodosa | GJS 04155 T | JQ365037 | JQ403317 | JQ403357 | JQ403394 | - |
| T. olida | CBS 21567 T | HM352884 | KJ021982 | KJ022058 | HM364334 | KM232024 |
| T. rubrococca | IMI 324475 T | KJ022275 | KJ022008 | KJ022061 | KJ022439 | KJ022329 |
| T. spinulospora | CGMCC 3.24133 T | OP272866 | OP223437 | OP223433 | OP272868 | OP586764 |
| T. torulosa | AR 4768A | JQ365031 | JQ403310 | JQ403350 | JQ403386 | JQ394702 |
| T. trachosa | GJS 9245 T | KF569832 | KF569842 | KF569851 | KF569879 | KF569869 |
| T. truncata | GJS 04357 T | JQ365039 | JQ403319 | JQ403359 | JQ403396 | KJ022324 |
| T. veuillotiana | AR 1751 | KJ022273 | JQ403305 | JQ403345 | JQ403382 | JQ394698 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Z.-Q.; Zhuang, W.-Y. New Species of Nectriaceae (Hypocreales) from China. J. Fungi 2022, 8, 1075. https://doi.org/10.3390/jof8101075
Zeng Z-Q, Zhuang W-Y. New Species of Nectriaceae (Hypocreales) from China. Journal of Fungi. 2022; 8(10):1075. https://doi.org/10.3390/jof8101075
Chicago/Turabian StyleZeng, Zhao-Qing, and Wen-Ying Zhuang. 2022. "New Species of Nectriaceae (Hypocreales) from China" Journal of Fungi 8, no. 10: 1075. https://doi.org/10.3390/jof8101075
APA StyleZeng, Z. -Q., & Zhuang, W. -Y. (2022). New Species of Nectriaceae (Hypocreales) from China. Journal of Fungi, 8(10), 1075. https://doi.org/10.3390/jof8101075