

Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast, Pathogen, and Jujube Fruit
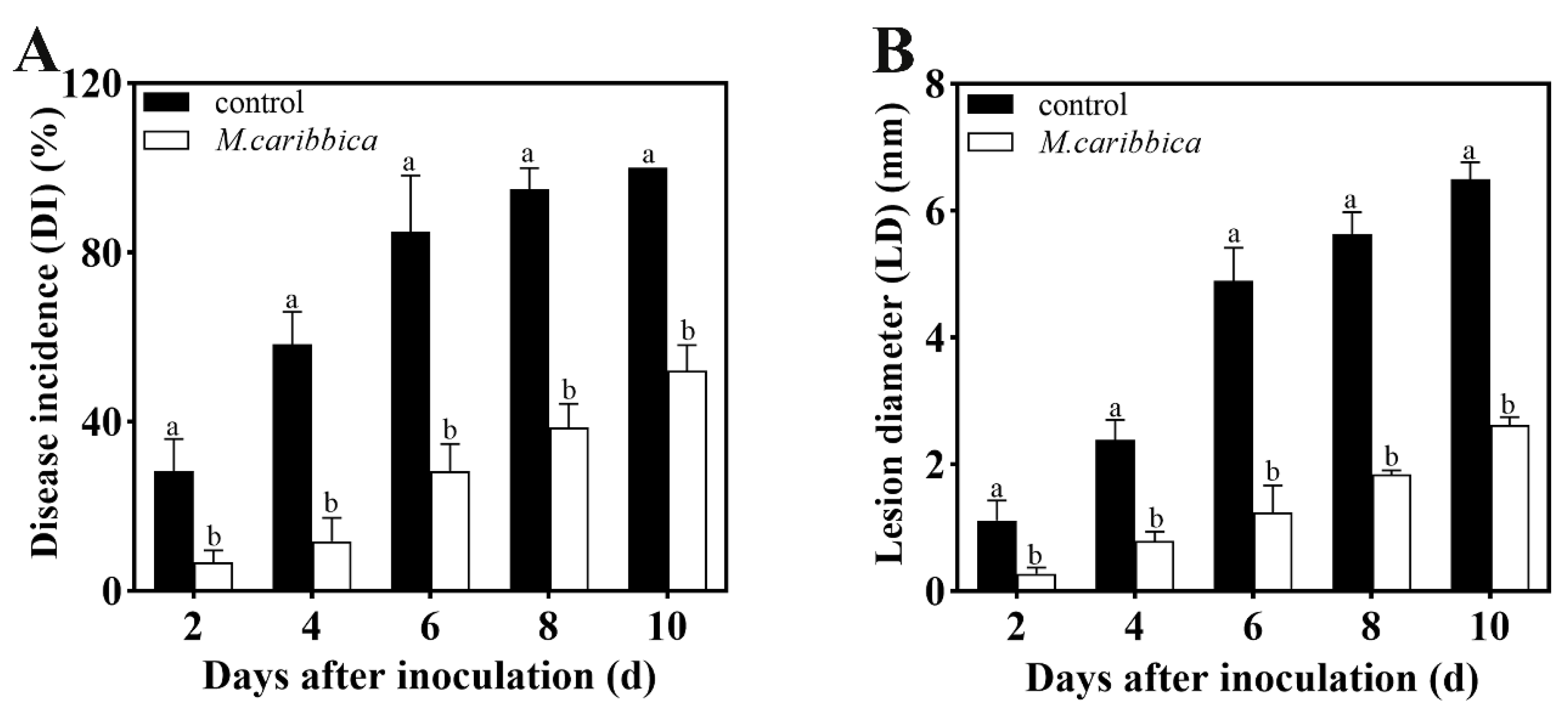
2.2. Effect of M. caribbica on the Biological Control of A. alternata on Jujube Fruit
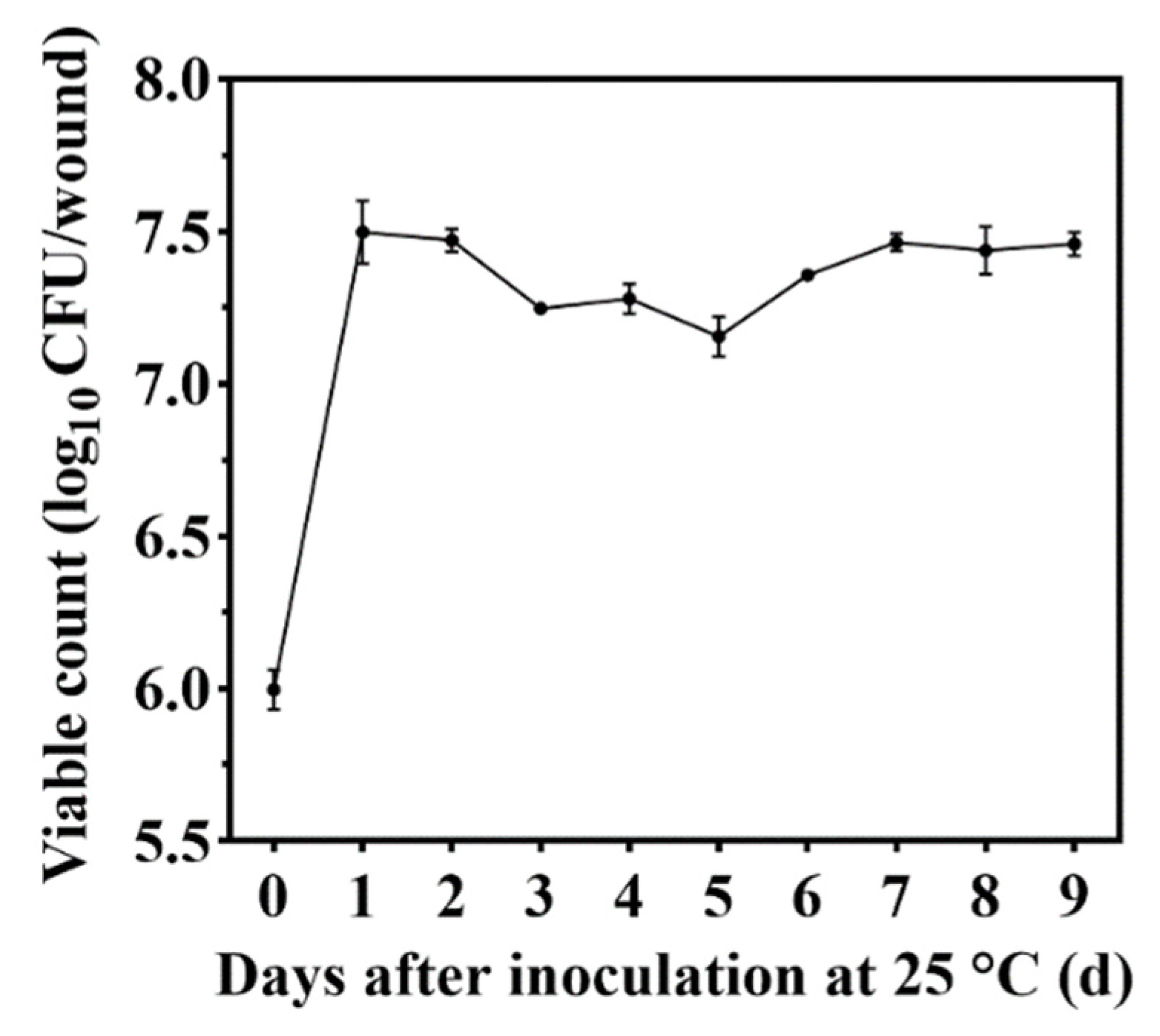
2.3. Dynamics of M. caribbica Colonization on Jujube Wounds
2.4. The Adhesion of M. caribbica to A. alternata
2.4.1. Microscopy Observation of M. caribbica and A. alternata
2.4.2. SEM Observation of M. caribbica and A. alternata in the Jujube Wound
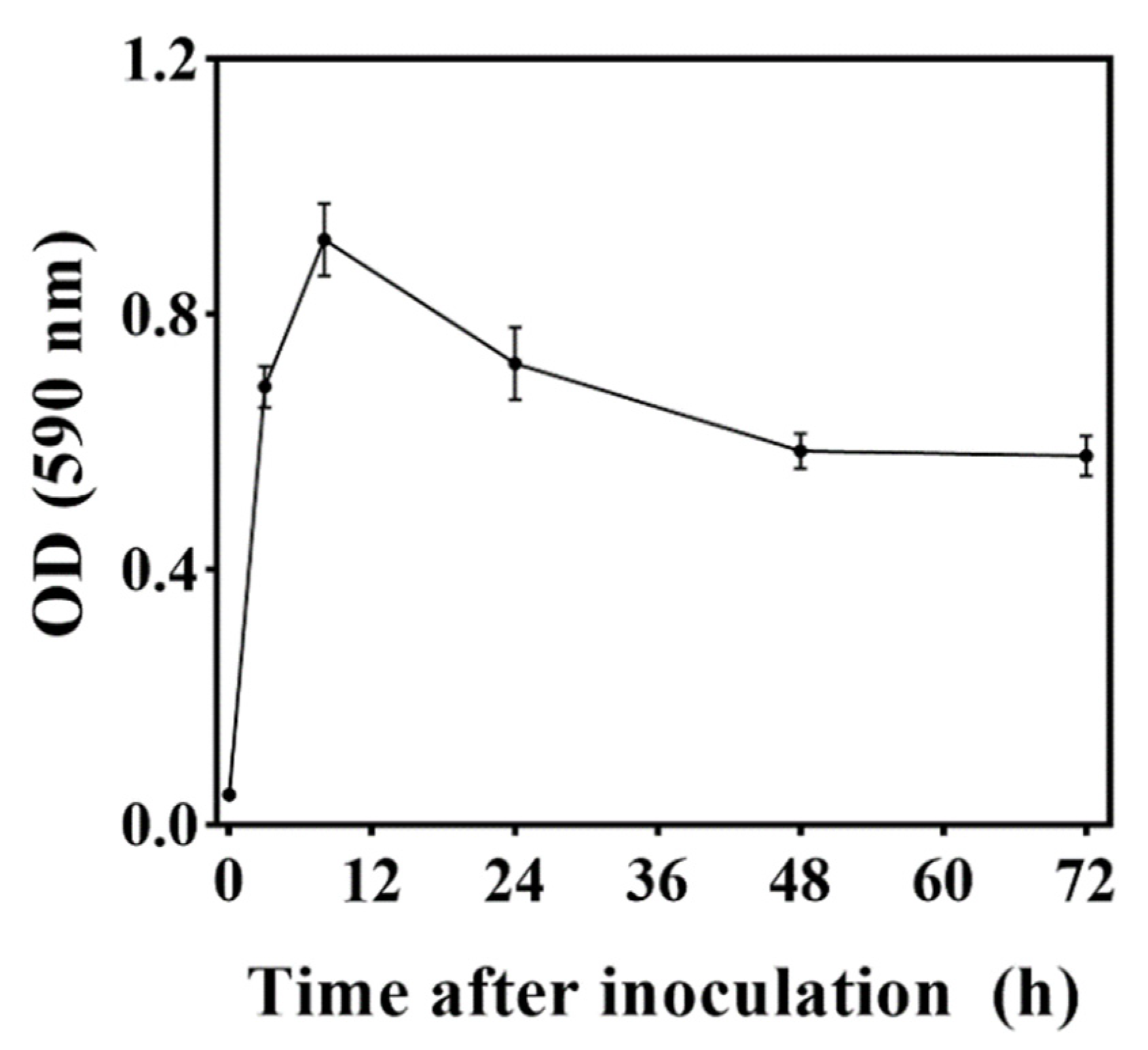
2.5. Detection of Biofilms Formed by M. caribbica
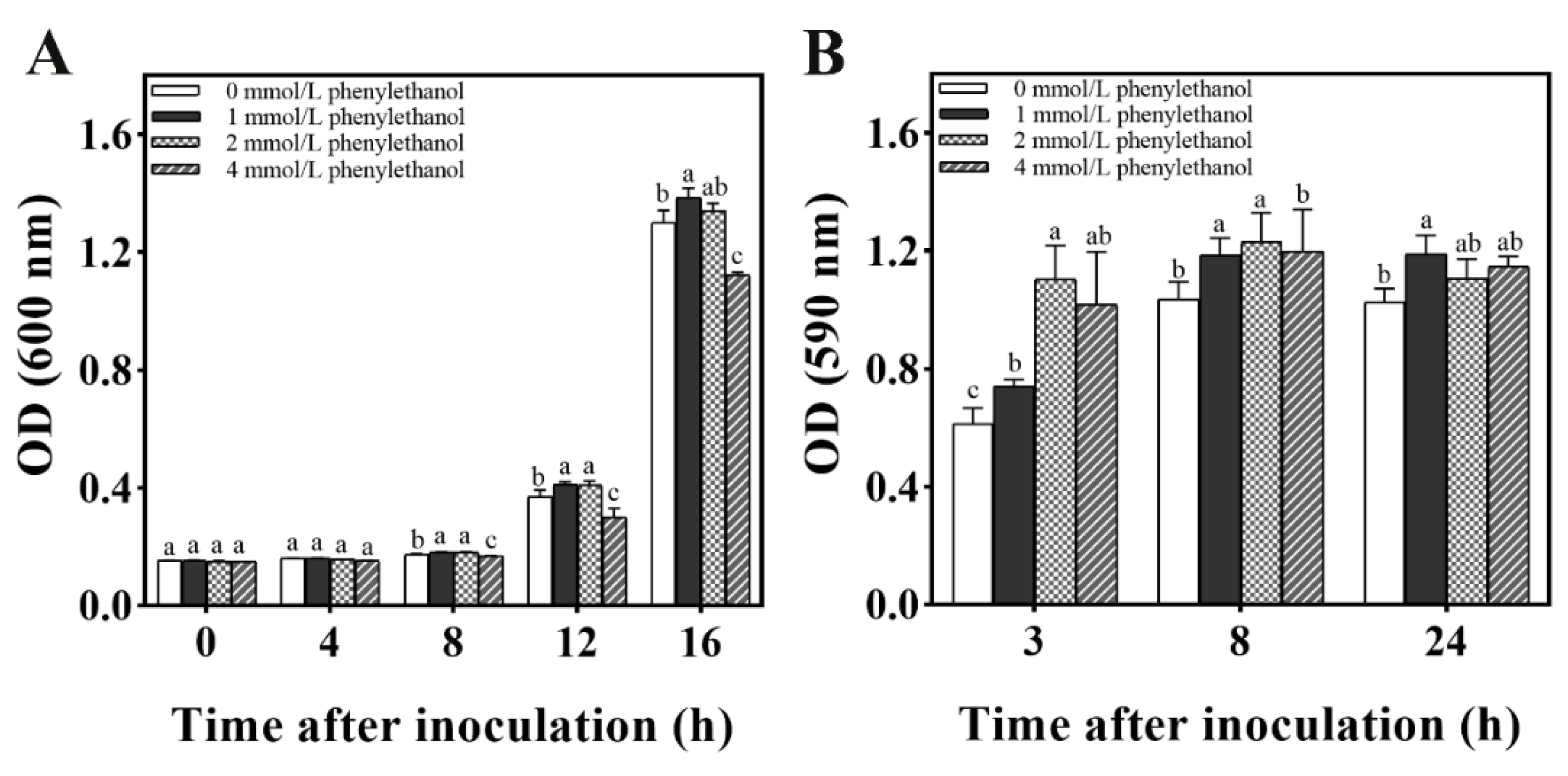
2.6. Effect of Phenylethanol on Biofilm Formation of M. caribbica
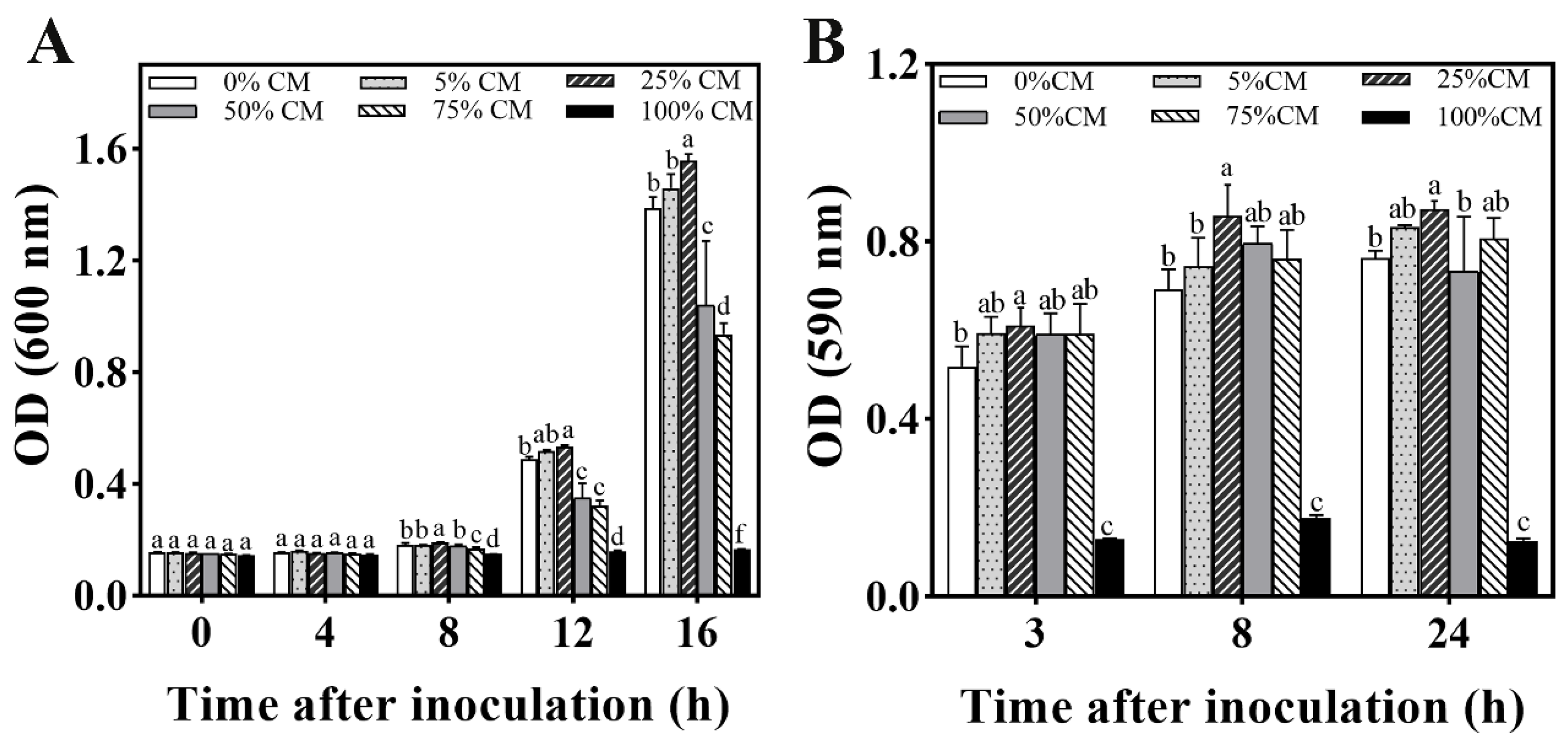
2.6.1. Biofilm Formation of M. caribbica in CM Medium
2.6.2. Evaluation of Phenylethanol Production from M. caribbica by HPLC
2.6.3. Effect of Phenylethanol on Biofilm Formation by M. caribbica
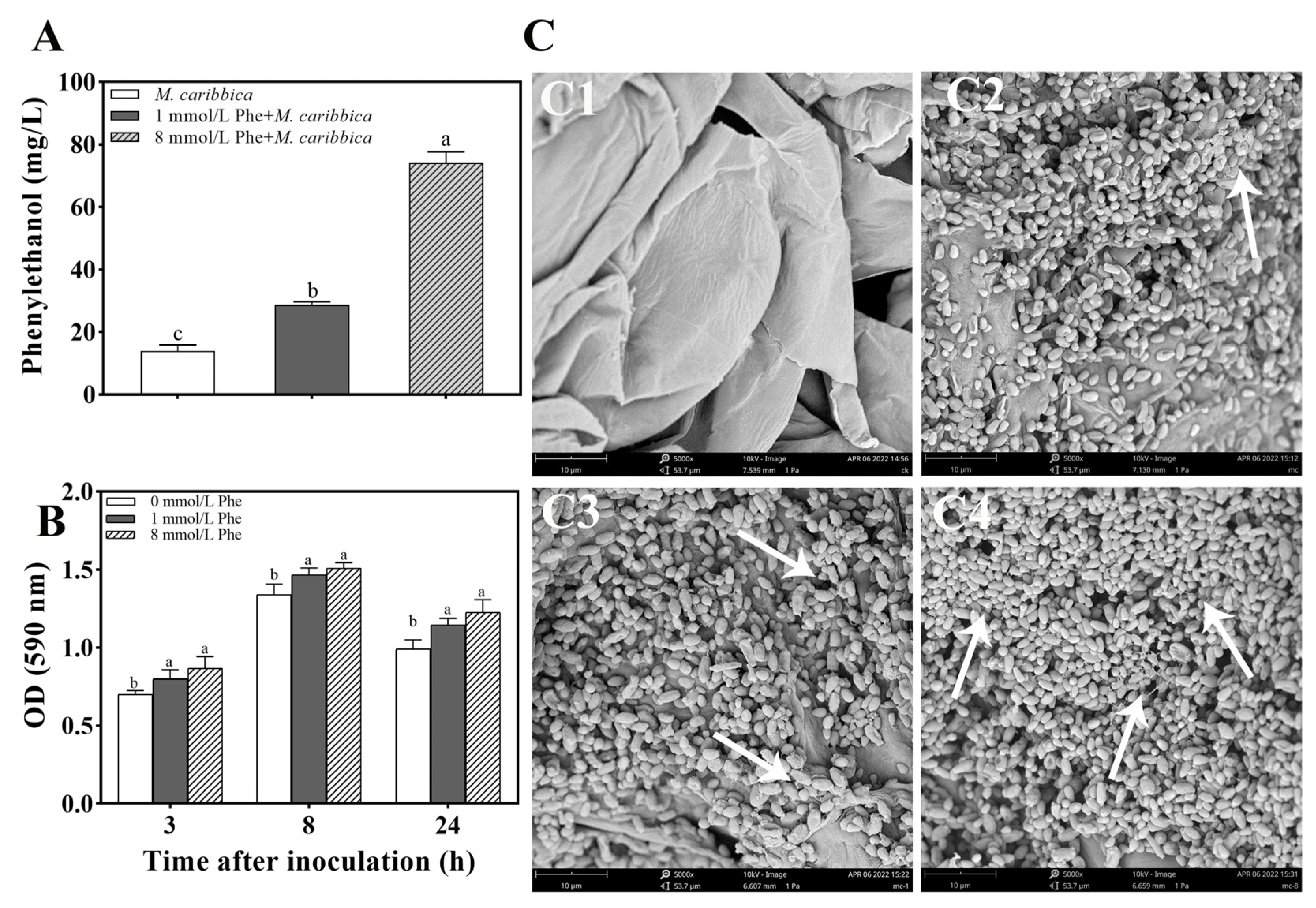
2.7. Influence of Phe on the Biocontrol Efficiency of M. caribbica
2.7.1. Effect of Phe on Phenylethanol Production of M. caribbica
2.7.2. Effect of Phe on Biofilm Formation of M. caribbica
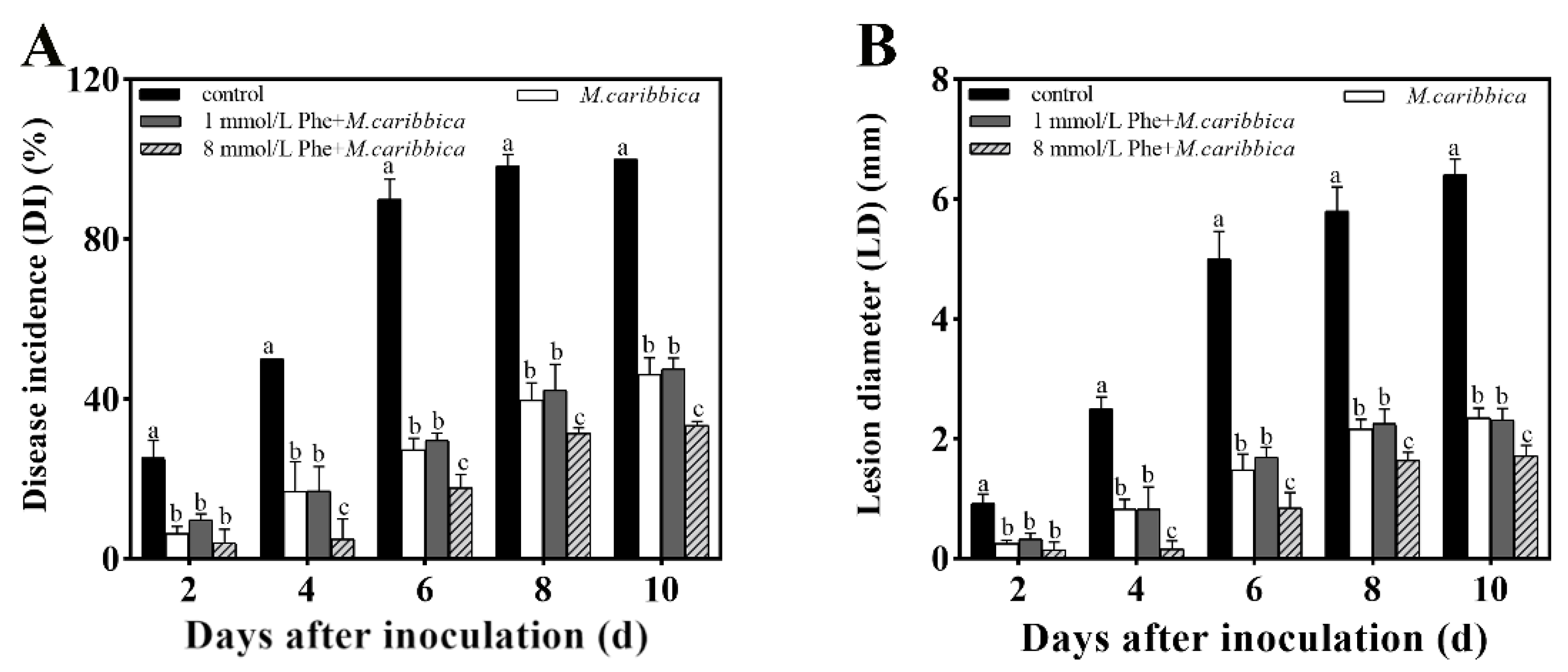
2.7.3. Effect of Phe on the Biocontrol Assay of M. caribbica
2.8. Statistical Analysis
3. Results
3.1. Efficiency of M. caribbica against Jujube Black Spot Rot
3.2. Colonization of Jujube Wounds by M. caribbica
3.3. Adherence of M. caribbica to A. alternata
3.3.1. In Vitro Interaction Study between M. caribbica and A. alternata
3.3.2. SEM Observation of M. caribbica and A. alternata on Jujube Wounds
3.4. Biofilm-Forming Ability of M. caribbica
3.5. The Mechanism of Biofilm Formation in M. caribbica
3.5.1. Development of M. caribbica Biofilm by CM Medium
3.5.2. Phenylethanol Determination in CM Medium by HPLC
3.5.3. Biofilm Formation in M. caribbica Induced by Phenylethanol
3.6. Influence of Phe on the Biocontrol Efficiency of M. caribbica
3.6.1. Efficacy of Phe on Phenylethanol Production in M. caribbica
3.6.2. Induction of Biofilm Formation in M. caribbica by Phe
3.6.3. Enhancement of Biocontrol Performance in M. caribbica by Phe
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, W.; Yuan, S.; Li, Q.; Sang, W.; Cao, J.; Jiang, W. Methyl p-coumarate inhibits black spot rot on jujube fruit through membrane damage and oxidative stress against Alternaria alternata. Postharvest Biol. Technol. 2018, 145, 230–238. [Google Scholar] [CrossRef]
- Yuan, S.; Yan, J.; Wang, M.; Ding, X.; Zhang, Y.; Li, W.; Cao, J.; Jiang, W. Transcriptomic and metabolic profiling reveals ‘green ring’ and ‘red ring’ on jujube fruit upon postharvest Alternaria alternata infection. Plant Cell Physiol. 2019, 60, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, F.; Xia, J.; Yu, T.; Wang, J.; Azhati, R.; Zheng, X.D. A combination of marine yeast and food additive enhances preventive effects on postharvest decay of jujubes (Zizyphus jujuba). Food Chem. 2011, 125, 835–840. [Google Scholar] [CrossRef]
- Guo, D.; Zhu, L.; Hou, X. Combination of UV-C treatment and Metschnikowia pulcherrimas for controlling Alternaria rot in postharvest winter jujube fruit. J. Food Sci. 2015, 80, M137–M141. [Google Scholar] [CrossRef]
- Qin, G.Z.; Tian, S.P. biocontrol of postharvest diseases of jujube fruit by Cryptococcus laurentii combined with a low dosage of fungicides under different storage conditions. Plant Dis. 2004, 88, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Sui, Y.; Li, J.; Tian, X.; Wang, Q. Biological control of postharvest fungal decays in citrus: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 861–870. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yao, S.; Deng, L.; Ming, J.; Zeng, K. Different mechanisms of action of isolated epiphytic yeasts against Penicillium digitatum and Penicillium italicum on citrus fruit. Postharvest Biol. Technol. 2019, 152, 100–110. [Google Scholar] [CrossRef]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Álvarez, N.A.; Vázquez-Juárez, R.; Ragazzo-Sánchez, J.A. Biocontrol action mechanisms of Cryptococcus laurentii on Colletotrichum gloeosporioides of mango. Crop. Prot. 2014, 65, 194–201. [Google Scholar] [CrossRef]
- Chi, M.; Li, G.; Liu, Y.; Liu, G.; Li, M.; Zhang, X.; Sun, Z.; Sui, Y.; Liu, J. Increase in antioxidant enzyme activity, stress tolerance and biocontrol efficacy of Pichia kudriavzevii with the transition from a yeast-like to biofilm morphology. Biol. Control 2015, 90, 113–119. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Černáková, L. Farnesol and Tyrosol: Secondary metabolites with a crucial quorum-sensing role in Candida biofilm development. Genes 2020, 11, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal quorum-sensing molecules and inhibitors with potential antifungal activity: A review. Molecules 2019, 24, 950. [Google Scholar] [CrossRef] [Green Version]
- Barriuso, J.; Hogan, D.A.; Keshavarz, T.; Martínez, M.J. Role of quorum sensing and chemical communication in fungal biotechnology and pathogenesis. FEMS Microbiol. Rev. 2018, 42, 627–638. [Google Scholar] [CrossRef]
- Wongsuk, T.; Pumeesat, P.; Luplertlop, N. Fungal quorum sensing molecules: Role in fungal morphogenesis and pathogenicity. J. Basic Microbiol. 2016, 56, 440–447. [Google Scholar] [CrossRef]
- Pu, L.; Jingfan, F.; Kai, C.; Chao-an, L.; Yunjiang, C. Phenylethanol promotes adhesion and biofilm formation of the antagonistic yeast Kloeckera apiculata for the control of blue mold on citrus. FEMS Yeast Res. 2014, 14, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Deng, B.; Ruan, C.; Deng, L.; Zeng, K. Phenylethanol as a quorum sensing molecule to promote biofilm formation of the antagonistic yeast Debaryomyces nepalensis for the control of black spot rot on jujube. Postharvest Biol. Technol. 2022, 185, 111788. [Google Scholar] [CrossRef]
- Yang, L.; Zheng, C.; Chen, Y.; Ying, H. FLO genes family and transcription factor mig1 regulate Saccharomyces cerevisiae biofilm formation during immobilized fermentation. Front. Microbiol. 2018, 9, 1860. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, F.; Yu, Y.; Ding, S.; Chen, T.; Sun, W.; Liang, C.; Yu, B.; Ying, H.; Liu, D.; et al. Effect of quorum-sensing molecule 2-phenylethanol and ARO genes on Saccharomyces cerevisiae biofilm. Appl. Microbiol. Biotechnol. 2021, 105, 3635–3648. [Google Scholar] [CrossRef]
- Sekar, B.S.; Lukito, B.R.; Li, Z. Production of natural 2-phenylethanol from glucose or glycerol with coupled Escherichia coli strains expressing L -phenylalanine biosynthesis pathway and artificial biocascades. ACS Sustain. Chem. Eng. 2019, 104, 177. [Google Scholar] [CrossRef]
- Mo, Q.; Chen, H.; Fan, C.; Zhang, D.; Liu, L.; Fu, B.; Yuan, J. Utilization of a styrene-derived pathway for 2-phenylethanol production in budding yeast. Appl. Microbiol. Biotechnol. 2021, 105, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H. Metabolic regulatory mechanisms and physiological roles of functional amino acids and their applications in yeast. Biosci. Biotech. Bioch. 2019, 83, 1449–1462. [Google Scholar] [CrossRef]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yi, L.; Ruan, C.; Yao, S.; Deng, L.; Zeng, K. Proline increases pigment production to improve oxidative stress tolerance and biocontrol ability of Metschnikowia citriensis. Front. Microbiol. 2019, 10, 1273. [Google Scholar] [CrossRef] [PubMed]
- Simões, L.A.; Cristina de Souza, A.; Ferreira, I.; Melo, D.S.; Lopes, L.A.A.; Magnani, M.; Schwan, R.F.; Dias, D.R. Probiotic properties of yeasts isolated from Brazilian fermented table olives. J. Appl. Microbiol. 2021, 131, 1983–1997. [Google Scholar] [CrossRef]
- Amorim, J.C.; Piccoli, R.H.; Duarte, W.F. Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages. Food Res. Int. 2018, 107, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.J.; Bressani, A.P.P.; Simão, J.B.P.; Pylro, V.S.; Dias, D.R.; Schwan, R.F. Dominant microbial communities and biochemical profile of pulped natural fermented coffees growing in different altitudes. Food Res. Int. 2022, 159. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Herrera, Y.Y.; Ortíz-Moreno, M.L. Yeast strains with antagonist activity against Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. and their phenotypic characterization. Egypt. J. Biol. Pest Control 2020, 30, 120. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.-e.; Zhao, L.; Jiang, S.; Abiso Godana, E.; Zhang, X.; Zhang, H. Efficacy of Meyerozyma caribbica in the biocontrol of blue mold in kiwifruit and mechanisms involved. Biol. Control 2022, 173, 105000. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.; Zhou, Y.; Yao, S.; Deng, L.; Zeng, K. Isolation, identification and in vitro screening of Chongqing orangery yeasts for the biocontrol of Penicillium digitatum on citrus fruit. Biol. Control 2017, 110, 18–24. [Google Scholar] [CrossRef]
- Chen, O.; Yi, L.; Deng, L.; Ruan, C.; Zeng, K. Screening antagonistic yeasts against citrus green mold and the possible biocontrol mechanisms of Pichia galeiformis (BAF03). J. Sci. Food Agric. 2020, 100, 3812–3821. [Google Scholar] [CrossRef] [PubMed]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.; Nicola, A.M.; Nieves, E.; Paes, H.C.; Williamson, P.R.; Silva-Pereira, I.; Casadevall, A. Quorum sensing-mediated, cell density-dependent regulation of growth and virulence in Cryptococcus neoformans. mBio 2013, 5-e00986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic yeasts: A promising alternative to chemical fungicides for controlling postharvest decay of fruit. J. Fungi 2020, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.A. Biological control of postharvest diseases of fruit. Eur. J. Plant Pathol. 2012, 133, 181–196. [Google Scholar] [CrossRef]
- Aguirre-Güitrón, L.; Calderón-Santoyo, M.; Lagarón, J.M.; Prieto, C.; Ragazzo-Sánchez, J.A. Formulation of the biological control yeast Meyerozyma caribbica by electrospraying process: Effect on postharvest control of anthracnose in mango (Mangifera indica L.) and papaya (Carica papaya L.). J. Sci. Food Agric. 2022, 102, 696–706. [Google Scholar] [CrossRef]
- Aguirre-Güitrón, L.; Calderón-Santoyo, M.; Bautista-Rosales, P.U.; Ragazzo-Sánchez, J.A. Application of powder formulation of Meyerozyma caribbica for postharvest control of Colletotrichum gloeosporioides in mango (Mangifera indica L.). Lebensm.-Wiss. Technol. 2019, 113, 108271. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Li, P.; Zhang, H.; Zhang, X.; Zheng, X.; Yang, Q.; Apaliya, M.T.; Boateng, N.A.S.; Sun, Y. The biocontrol effect of Sporidiobolus pararoseus Y16 against postharvest diseases in table grapes caused by Aspergillus niger and the possible mechanisms involved. Biol. Control 2017, 113, 18–25. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, C.; Yi, L.; Deng, L.; Yao, S.; Zeng, K. Biocontrol ability and action mechanism of Metschnikowia citriensis against Geotrichum citri-aurantii causing sour rot of postharvest citrus fruit. Food Microbiol. 2020, 87, 103375. [Google Scholar] [CrossRef]
- Bandara, H.M.H.N.; Lam, O.L.T.; Jin, L.J.; Samaranayake, L. Microbial chemical signaling: A current perspective. Crit. Rev. Microbiol. 2012, 38, 217–249. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi—A review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iñiguez-Moreno, M.; Ragazzo-Sánchez, J.A.; Barros-Castillo, J.C.; Sandoval-Contreras, T.; Calderón-Santoyo, M. Sodium alginate coatings added with Meyerozyma caribbica: Postharvest biocontrol of Colletotrichum gloeosporioides in avocado (Persea americana Mill. cv. Hass). Postharvest Biol. Technol. 2020, 163, 111123. [Google Scholar] [CrossRef]
- Choińska, R.; Piasecka-Jóźwiak, K.; Chabłowska, B.; Dumka, J.; Łukaszewicz, A. Biocontrol ability and volatile organic compounds production as a putative mode of action of yeast strains isolated from organic grapes and rye grains. Anton. Leeuw. Int. J. G. 2020, 113, 1135–1146. [Google Scholar] [CrossRef]
- Gori, K.; Knudsen, P.B.; Nielsen, K.F.; Arneborg, N.; Jespersen, L. Alcohol-based quorum sensing plays a role in adhesion and sliding motility of the yeast Debaryomyces hansenii. FEMS Yeast Res. 2011, 11, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Avbelj, M.; Zupan, J.; Raspor, P. Quorum-sensing in yeast and its potential in wine making. Appl. Microbiol. Biotechnol. 2016, 100, 7841–7852. [Google Scholar] [CrossRef]
- Mitri, S.; Koubaa, M.; Maroun, R.G.; Rossignol, T.; Nicaud, J.-M.; Louka, N. Bioproduction of 2-phenylethanol through yeast fermentation on synthetic media and on agro-industrial waste and by-products: A review. Foods 2022, 1, 109. [Google Scholar] [CrossRef]
- Dai, J.; Xia, H.; Yang, C.; Chen, X. Sensing, uptake and catabolism of L-phenylalanine during 2-phenylethanol biosynthesis via the Ehrlich pathway in Saccharomyces cerevisiae. Front. Microbiol. 2021, 12, 601963. [Google Scholar] [CrossRef]
- Berrocal, A.; Navarrete, J.; Oviedo, C.; Nickerson, K.W. Quorum sensing activity in Ophiostoma ulmi: Effects of fusel oils and branched chain amino acids on yeast-mycelial dimorphism. J. Appl. Microbiol. 2012, 113, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Sanna, M.L.; Zara, S.; Zara, G.; Migheli, Q.; Budroni, M.; Mannazzu, I. Pichia fermentans dimorphic changes depend on the nitrogen source. Fungal Biol. 2012, 116, 769–777. [Google Scholar] [CrossRef]
- Klein, M.N.; Kupper, K.C. Biofilm production by Aureobasidium pullulans improves biocontrol against sour rot in citrus. Food Microbiol. 2018, 69, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, H.; Qi, T.; Deng, L.; Yi, L.; Zeng, K. Influence of arginine on the biocontrol efficiency of Metschnikowia citriensis against Geotrichum citri-aurantii causing sour rot of postharvest citrus fruit. Food Microbiol. 2022, 101, 103888. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Q.; Lei, X.; Zhang, H.; Deng, L.; Yi, L.; Zeng, K. Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot. J. Fungi 2022, 8, 1313. https://doi.org/10.3390/jof8121313
Deng Q, Lei X, Zhang H, Deng L, Yi L, Zeng K. Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot. Journal of Fungi. 2022; 8(12):1313. https://doi.org/10.3390/jof8121313
Chicago/Turabian StyleDeng, Qian, Xingmeng Lei, Hongyan Zhang, Lili Deng, Lanhua Yi, and Kaifang Zeng. 2022. "Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot" Journal of Fungi 8, no. 12: 1313. https://doi.org/10.3390/jof8121313
APA StyleDeng, Q., Lei, X., Zhang, H., Deng, L., Yi, L., & Zeng, K. (2022). Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot. Journal of Fungi, 8(12), 1313. https://doi.org/10.3390/jof8121313