
Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Materials, Experimental Design
2.2. Infection and Colonization of C. globosum ND35 on Cucumber Radicle in Vitro
2.3. Determination of Endogenous Metabolites and Enzymes in Cucumber Radicle
2.4. Transcriptome Sequencing and Functional Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. Detection of Secondary Metabolites in Fermentation Broth of Strain ND35
2.7. Statistical Analysis
3. Results
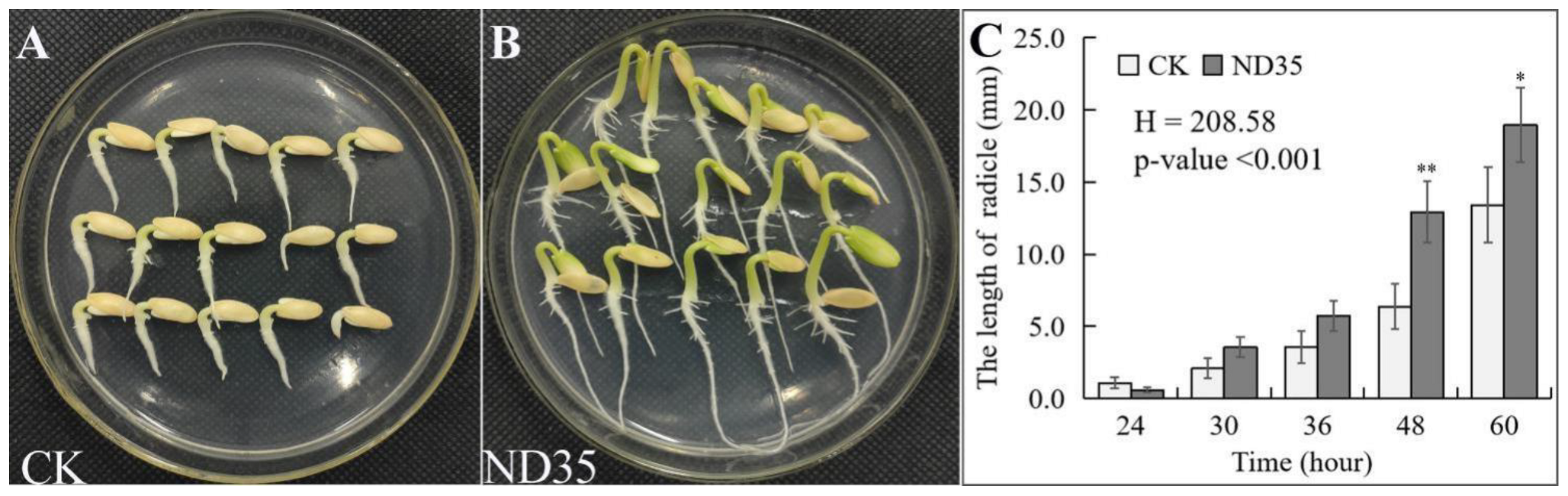
3.1. Effect of C. globosum Strain ND35 on the Cucumber Growth
3.2. Secondary Metabolites of C. globosum Strain ND35
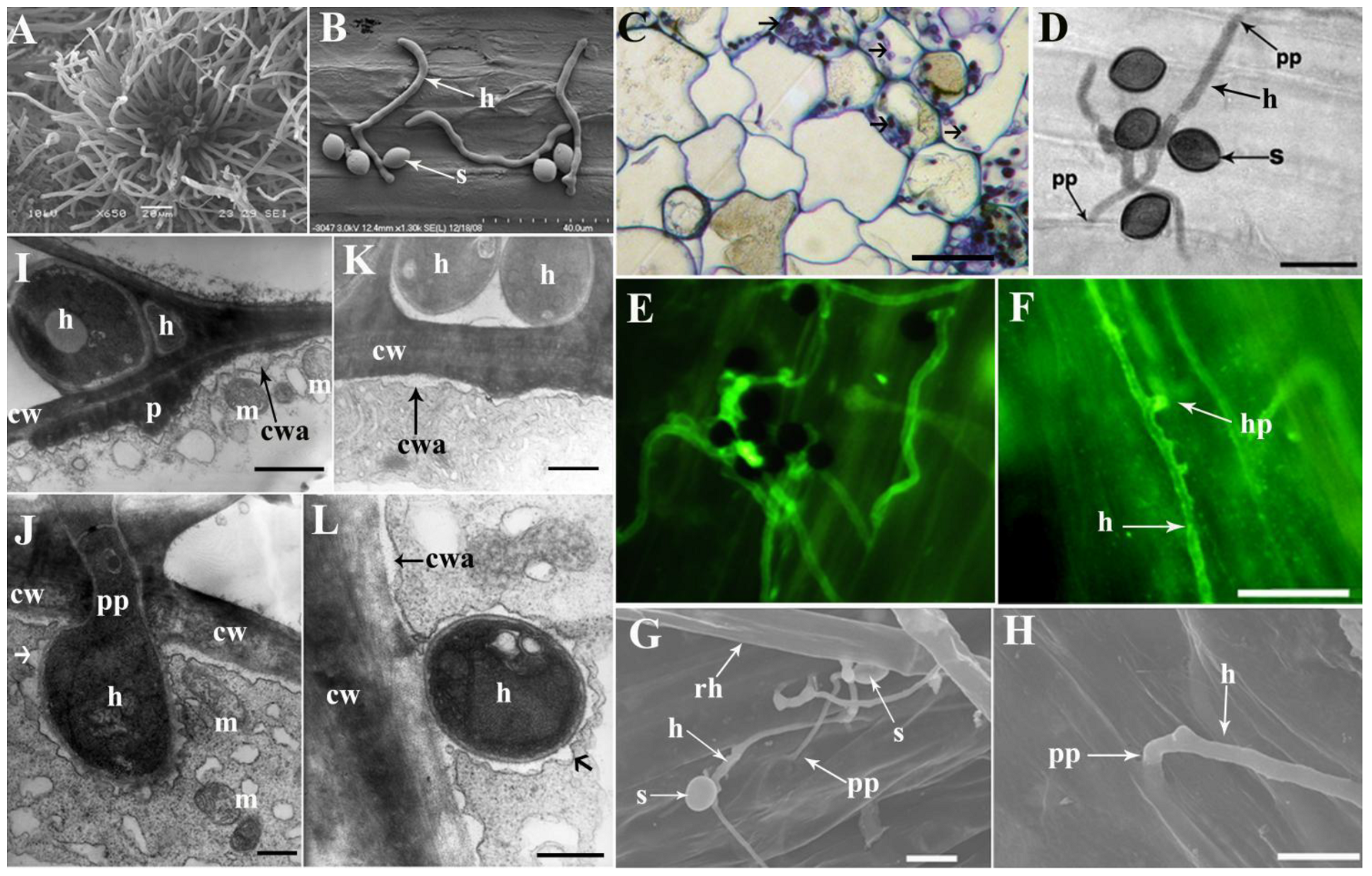
3.3. Infection and Colonization of C. globosum Strain ND35 Spores on the Radicle of Cucumber Seeds
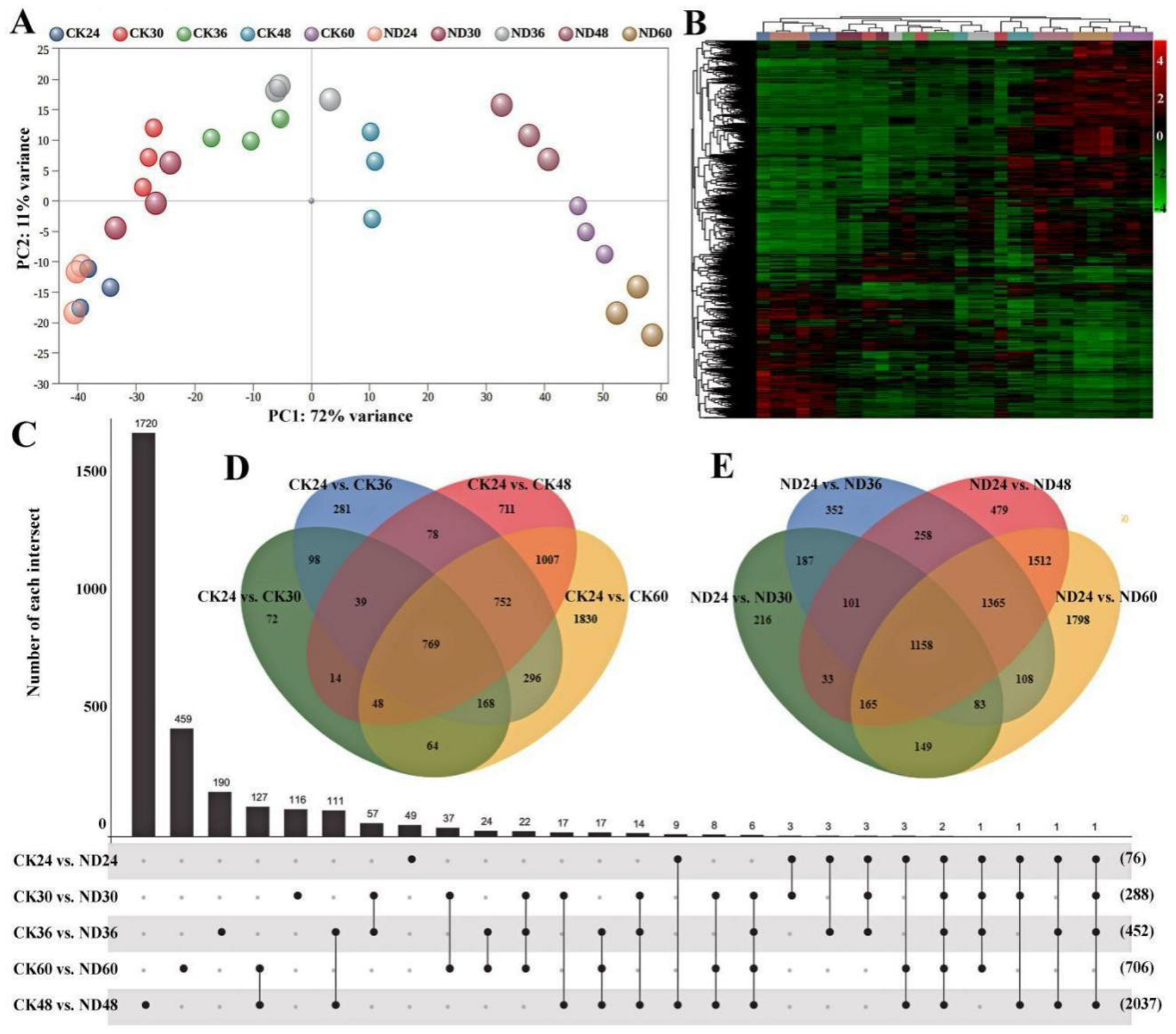
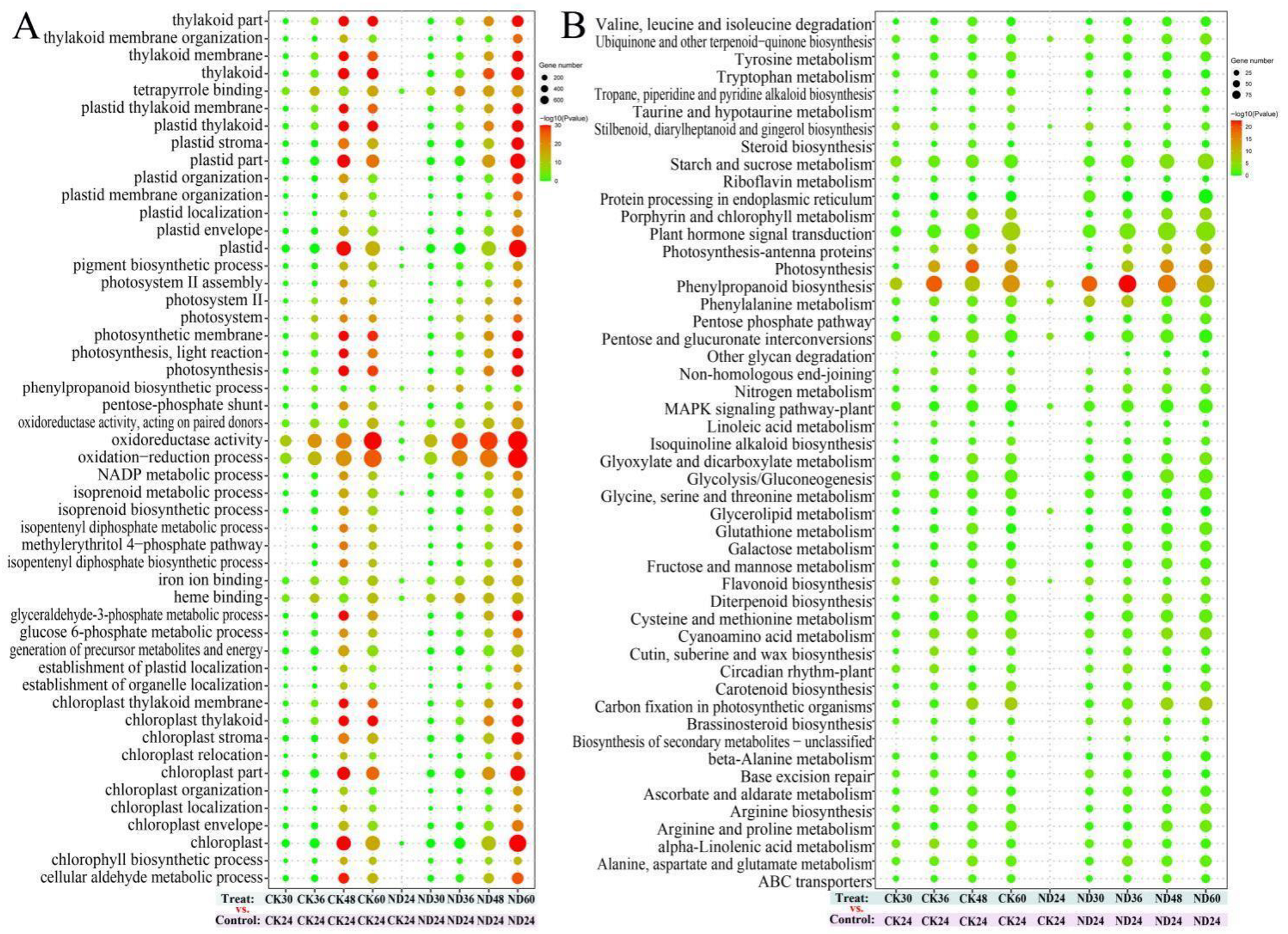
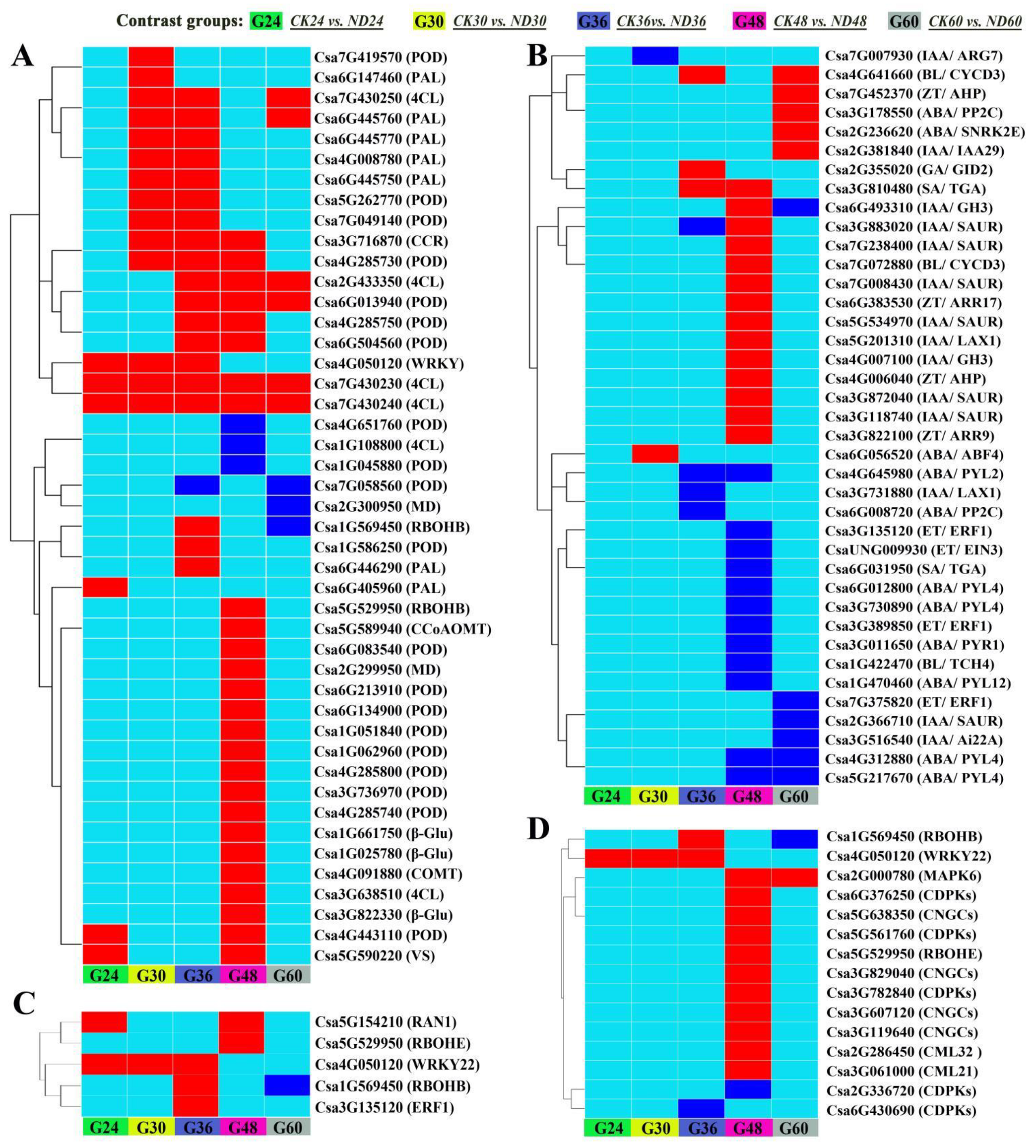
3.4. Transcriptome Sequencing and Functional Analysis of Differentially Expressed Genes (DEGs)
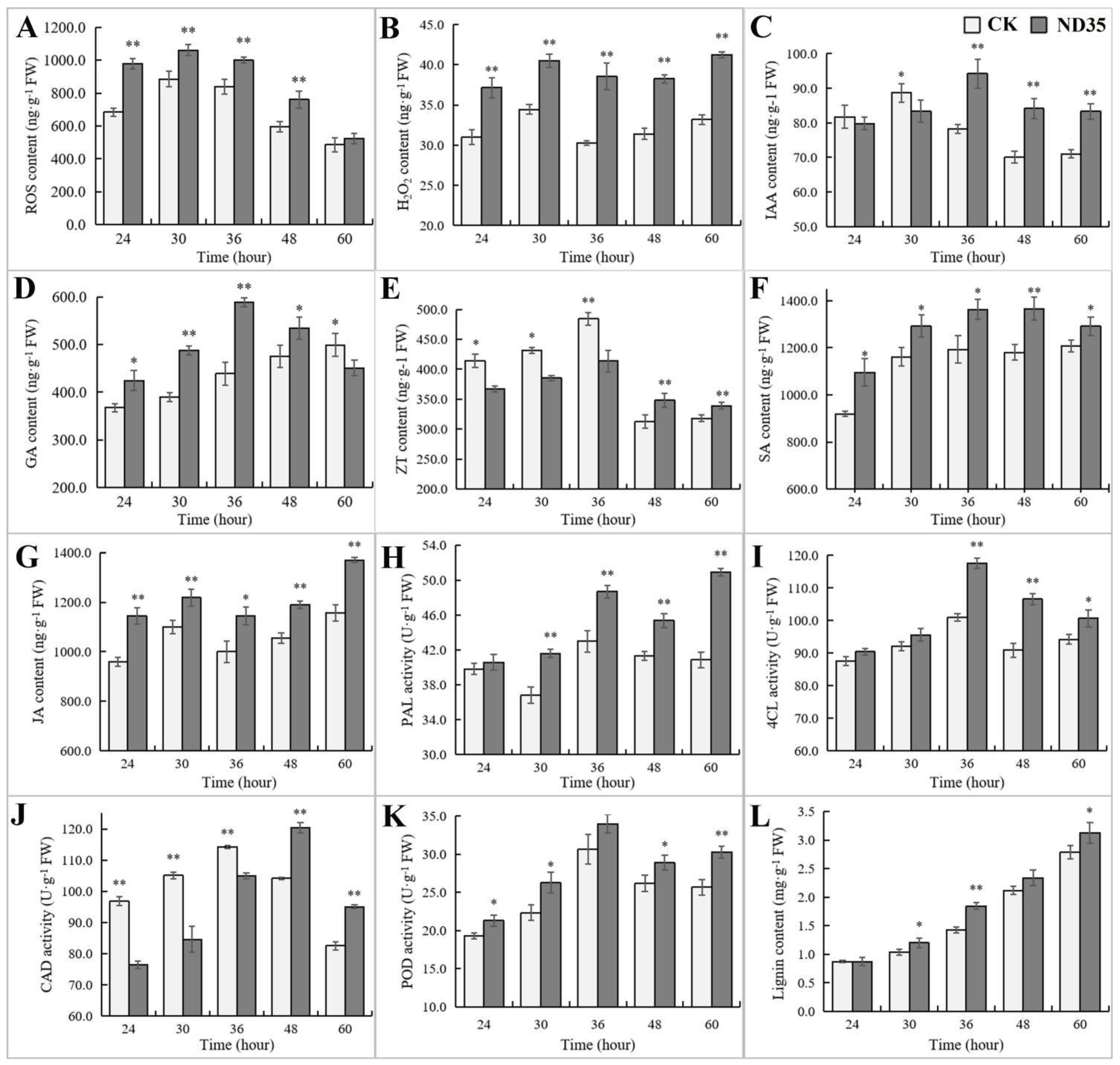
3.5. Influence of C. globosum Strain ND35 on the Endogenous Hormonal and Other Secondary Metabolites Levels of Cucumber
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.E.; Torres-Mendoza, D.; Cubilla-Rios, L. Patents on endophytic fungi for agriculture and bio- and phytoremediation applications. Microorganismas 2020, 8, 1237. [Google Scholar] [CrossRef]
- Ernst, M.; Mendgen, K.W.; Wirsel, S.G.R. Endophytic fungal mutualists: Seed-borne Stagonospora spp. Enhance reed biomass production in axenic microcosms. Mol. Plant Microbe Interact. 2003, 16, 580–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, K.X.; Mendgen, K. Seed-transmitted beneticial endophytic Stagonospora sp. can penetrate the walls of the root epidermis, but does not proliferate in the cortex, of Phragmites australis. Can. J. Bot. 2006, 84, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Mejia, L.; Rojas, E.I.; Maynard, Z.; Van Bael, S.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Lee, I.-J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef]
- Bianco, C.; Andreozzi, A.; Romano, S.; Fagorzi, C.; Cangioli, L.; Prieto, P.; Cisse, F.; Niangado, O.; Sidibé, A.; Pianezze, S.; et al. Endophytes from african rice (Oryza glaberrima L.) efficiently colonize asian rice (Oryza sativa L.) stimulating the activity of its antioxidant enzymes and increasing the content of nitrogen, carbon, and chlorophyll. Microoranisms 2021, 9, 1714. [Google Scholar] [CrossRef]
- Gao, F.K.; Dai, C.C.; Liu, X.Z. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res. 2010, 4, 1346–1351. [Google Scholar]
- Tseng, Y.-H.; Rouina, H.; Groten, K.; Rajani, P.; Furch, A.C.U.; Reichelt, M.; Baldwin, I.T.; Nataraja, K.N.; Shaanker, R.U.; Oelmüller, R. An endophytic trichoderma strain promotes growth of its hosts and defends against pathogen attack. Front. Plant Sci. 2020, 11, 573670. [Google Scholar] [CrossRef]
- Serfling, A.; Wirsel, S.G.R.; Lind, V.; Deising, H.B. Performance of the biocontrol fungus Piriformospora indica on wheat under greenhouse and field conditions. Phytopathology 2007, 97, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Franken, P. The plant strengthening root endophyte Piriformospora indica: Potential application and the biology behind. Appl. Microbiol. Biotechnol. 2012, 96, 1455–1464. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.; Saiyam, D.; Kumar, A.; Hashem, A.; Allah, E.F.A.; Khan, M.L. Bacterial root endophytes: Characterization of their competence and plant growth promotion in soybean (Glycine max (L.) Merr.) under drought stress. Int. J. Environ. Res. Public Health 2021, 18, 931. [Google Scholar] [CrossRef]
- Fontana, D.; de Paula, S.; Torres, A.; de Souza, V.; Pascholati, S.; Schmidt, D.; Neto, D.D. Endophytic fungi: Biological control and induced resistance to phytopathogens and abiotic stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial seed endophyte shapes disease resistance in rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldan, A.; Albacete, A.; Pascual, J. The interaction with arbuscular mycorrhizal fungi or Trichoderma harzianum alters the shoot hormonal profile in melon plants. Phytochemistry 2011, 72, 223–229. [Google Scholar] [CrossRef]
- Bonini, P.; Rouphael, Y.; Miras-Moreno, B.; Lee, B.; Cardarelli, M.; Erice, G.; Cirino, V.; Lucini, L.; Colla, G. A microbial-based biostimulant enhances sweet pepper performance by metabolic reprogramming of phytohormone profile and secondary metabolism. Front. Plant Sci. 2020, 11, 567388. [Google Scholar] [CrossRef]
- Ismail, M.; Amin, M.; Eid, A.; Hassan, S.; Mahgoub, H.; Lashin, I.; Abdelwahab, A.; Azab, E.; Gobouri, A.; Elkelish, A.; et al. Comparative study between exogenously applied plant growth hormones versus metabolites of microbial endophytes as plant growth-promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Chand, K.; Shah, S.; Sharma, J.; Paudel, M.R.; Pant, B. Isolation, characterization, and plant growth-promoting activities of endophytic fungi from a wild orchid Vanda cristata. Plant Signal. Behav. 2020, 15, 1744294. [Google Scholar] [CrossRef]
- Venneman, J.; Vandermeersch, L.; Walgraeve, C.; Audenaert, K.; Ameye, M.; Verwaeren, J.; Steppe, K.; Van Langenhove, H.; Haesaert, G.; Vereecke, D. Respiratory CO2 combined with a blend of volatiles emitted by endophytic Serendipita strains strongly stimulate growth of arabidopsis implicating auxin and cytokinin signaling. Front. Plant Sci. 2020, 11, 544435. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, Y.; Qin, J.; Qu, X.; Liu, J.; Li, X.; Pan, H. Antifungal metabolites produced by Chaetomium globosum No.04, an endophytic fungus isolated from ginkgo biloba. Indian J. Microbiol. 2013, 53, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Liu, X.; Kang, Z.; Mendgen, K. Mycoparasitism of Rhizoctonia solani by endophytic Chaetomium spirale ND35: Ultrastructure and cytochemistry of the interaction. J. Phytopathol. 2005, 153, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Liu, X.G.; Cao, R.H.; Gao, K.X.; Song, Y. Production of antibiotics by Chaetomium spirale ND35 and the role in biocontrol of plant diseases. Acta Phytophylacica Sin. 2007, 34, 51–56. [Google Scholar]
- Istifadah, N.; McGee, P.A. Endophytic Chaetomium globosum reduces development of tan spot in wheat caused by Pyrenophora tritici-repentis. Australas. Plant Pathol. 2006, 35, 411–418. [Google Scholar] [CrossRef]
- Zhao, S.-S.; Zhang, Y.-Y.; Yan, W.; Cao, L.-L.; Xiao, Y.; Ye, Y.-H. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites. FEMS Microbiol. Lett. 2016, 364, fnw287. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Kundu, A.; Dutta, A.; Saha, S.; Das, A.; Bhowmik, A. Chemo-profiling of bioactive metabolites from Chaetomium globosum for biocontrol of Sclerotinia rot and plant growth promotion. Fungal Biol. 2021, 125, 167–176. [Google Scholar] [CrossRef]
- Alhamed, M.F.A.; Shebany, Y.M. Endophytic Chaetomium globosum enhances maize seedling copper stress tolerance. Plant Biol. 2012, 14, 859–863. [Google Scholar] [CrossRef]
- Zhai, X.; Luo, D.; Li, X.; Han, T.; Jia, M.; Kong, Z.; Ji, J.; Rahman, K.; Qin, L.; Zheng, C. Endophyte Chaetomium globosum D38 promotes bioactive constituents accumulation and root production in Salvia miltiorrhiza. Front. Microbiol. 2018, 8, 2694. [Google Scholar] [CrossRef]
- Liu, X.G.; Gao, K.X.; Gu, J.C.; Du, J.L.; Tang, X.G. Testing on the antagonism of the dominant of endophytic fungi from Populus tomentosa, Chaetomium ND35 in the laboratory. Sci. Silvae Sin. 1999, 35, 57–61. [Google Scholar]
- Meng, Q.G.; Liu, X.G.; Mi, S.W.; Gao, K.X.; Kang, Z.S.; Wang, H.Q. Chaetomium globosum ND35 colonization and influence on enzymes activities in poplar. Acta Phytophylacica Sin. 2009, 36, 91–92. [Google Scholar]
- Mi, S.W.; Dai, Y.; Liu, X.G.; Meng, Q.G.; Gao, K.X.; Kurt, M. Infection and colonization of Chaetomium globosum ND35 on host plant. Acta Phytophylacica Sin. 2011, 38, 493–498. [Google Scholar]
- Cong, G.Q.; Yin, C.L.; He, B.L.; Li, L.; Gao, K.X. Effect of the endophytic fungus Chaetomium globosum ND35 on the growth and resistance to drought of winter wheat at the seedling stage under water stress. Acta Ecologica Sin. 2015, 35, 6120–6128. [Google Scholar]
- Song, F.H.; Wang, S.; Zhang, X.F.; Gao, K.X.; Yin, C.L.; Chen, X.S.; Mao, Z.Q. Effects of Chaetomium globosum ND35 fungal fertilizer on continuous cropping soil microorganism and Malus hupehensis seedling biomass. Acta Hortic. Sin. 2015, 42, 205–213. [Google Scholar]
- Li, J.H.; Gao, K.X.; Wan, L.; Cao, G.Y.; Jiao, F.Y.; Wang, Y.P.; Tian, Y.H.; Zhao, B.J.; Li, C.R. Effects of microbial agent on the growth of Catalpa bungei seedlings and the diversity of bacterial community in rhizosphere soil. Acta Ecol. Sin. 2020, 40, 7588–7601. [Google Scholar]
- Kang, Z.S.; Shang, H.S.; Li, Z.Q. Fluorescence staining technique of wheat rust tissue. Plant Prot. 1993, 17, 27. [Google Scholar]
- Tang, L.; Yu, X.; Zhang, L.; Zhang, L.; Chen, L.; Zou, S.; Liang, Y.; Yu, J.; Dong, H. Mitochondrial FgEch1 is responsible for conidiation and full virulence in Fusarium graminearum. Curr. Genet. 2020, 66, 361–371. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Genet. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Huang, W.; Xu, Y.; Zhou, Q.; Wang, S.; Ruan, J.; Huang, S.; Zhang, Z. A chromosome-scale genome assembly of cucumber (Cucumis sativus L.). GigaScience 2019, 8, 072. [Google Scholar] [CrossRef] [Green Version]
- Sarsaiya, S.; Shi, J.; Chen, J. A comprehensive review on fungal endophytes and its dynamics on Orchidaceae plants: Current research, challenges, and future possibilities. Bioengineered 2019, 10, 316–334. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Sahib, M.R.; Amna, A.; Opiyo, S.O.; Zhao, Z.; Gao, Y.G. Culturable endophytic fungal communities associated with plants in organic and conventional farming systems and their effects on plant growth. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Bowman, E.A.; Arnold, A.E. Drivers and implications of distance decay differ for ectomycorrhizal and foliar endophytic fungi across an anciently fragmented landscape. ISME J. 2021, 15, 3437–3454. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Jung, H.-Y.; Lee, J.-H.; Lee, I.-J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Malmierca, M.G.; McCormick, S.P.; Cardoza, R.E.; Monte, E.; Alexander, N.J.; Gutiérrez, S. Trichodiene production in a Trichoderma harzianum erg1-silenced strain provides evidence of the importance of the sterol biosynthetic pathway in inducing plant defense-related gene expression. Mol. Plant Microbe Interact. 2015, 28, 1181–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindo, L.; Cardoza, R.E.; Lorenzana, A.; Casquero, P.A.; Gutiérrez, S. Identification of plant genes putatively involved in the perception of fungal ergosterol-squalene. J. Integr. Plant Biol. 2020, 62, 927–947. [Google Scholar] [CrossRef] [Green Version]
- Parniske, M. Molecular genetics of the arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2004, 7, 414–421. [Google Scholar] [CrossRef]
- Harrison, M.J. Signaling in the arbuscular mycorrhizal symbiosis. Annu. Rev. Microbiol. 2005, 59, 19–42. [Google Scholar] [CrossRef]
- Russo, G.; Carotenuto, G.; Fiorilli, V.; Volpe, V.; Faccio, A.; Bonfante, P.; Chabaud, M.; Chiapello, M.; Van Damme, D.; Genre, A. TPLATE recruitment reveals endocytic dynamics at sites of symbiotic interface assembly in arbuscular mycorrhizal interactions. Front. Plant Sci. 2019, 10, 1628. [Google Scholar] [CrossRef]
- Roeder, A.H.K.; Otegui, M.S.; Dixit, R.; Anderson, C.T.; Faulkner, C.; Zhang, Y.; Harrison, M.J.; Kirchhelle, C.; Goshima, G.; Coate, J.E.; et al. Fifteen compelling open questions in plant cell biology. Plant Cell 2021, 34, 72–102. [Google Scholar] [CrossRef]
- MacLean, A.M.; Bravo, A.; Harrison, M.J. Plant signaling and metabolic pathways enabling arbuscular mycorrhizal symbiosis. Plant Cell 2017, 29, 2319–2335. [Google Scholar] [CrossRef]
- Choi, J.; Summers, W.; Paszkowski, U. Mechanisms underlying establishment of arbuscular mycorrhizal symbioses. Annu. Rev. Phytopathol. 2018, 56, 135–160. [Google Scholar] [CrossRef]
- De Palma, M.; Salzano, M.; Villano, C.; Aversano, R.; Lorito, M.; Ruocco, M.; Docimo, T.; Piccinelli, A.L.; D’Agostino, N.; Tucci, M. Transcriptome reprogramming, epigenetic modifications and alternative splicing orchestrate the tomato root response to the beneficial fungus Trichoderma harzianum. Hortic. Res. 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, T.; Liu, W.-C.; Zhang, D.-P.; Dong, D.; Wu, H.-L.; Zhang, T.-T.; Liu, D.-W. Transcriptomic insights into growth promotion effect of Trichoderma afroharzianum TM2-4 microbial agent on tomato plants. J. Integr. Agric. 2021, 20, 1266–1276. [Google Scholar] [CrossRef]
- Reddy, A.S.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [Green Version]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.-P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signaling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Hu, C.-H.; Wang, P.-Q.; Zhang, P.-P.; Nie, X.-M.; Li, B.-B.; Tai, L.; Liu, W.-T.; Li, W.-Q.; Chen, K.-M. NADPH Oxidases: The vital performers and center hubs during plant growth and signaling. Cells 2020, 9, 437. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Du, L.; Poovaiah, B. Calcium signaling and biotic defense responses in plants. Plant Signal. Behav. 2014, 9, e973818. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Pan, Y.; Tian, W.; Dong, M.; Zhu, H.; Luan, S.; Li, L. Arabidopsis CNGC14 mediates calcium influx required for tip growth in root hairs. Mol. Plant 2017, 10, 1004–1006. [Google Scholar] [CrossRef] [Green Version]
- Zeb, Q.; Wang, X.; Hou, C.; Zhang, X.; Dong, M.; Zhang, S.; Zhang, Q.; Ren, Z.; Tian, W.; Zhu, H.; et al. The interaction of CaM7 and CNGC14 regulates root hair growth in arabidopsis. J. Integr. Plant Biol. 2020, 62, 887–896. [Google Scholar] [CrossRef]
- Li, X.; Sun, Y.; Yang, Y.; Yang, X.; Xue, W.; Wu, M.; Chen, P.; Weng, Y.; Chen, S. Transcriptomic and histological analysis of the response of susceptible and resistant cucumber to Meloidogyne incognita infection revealing complex resistance via multiple signaling pathways. Front. Plant Sci. 2021, 12, 675429. [Google Scholar] [CrossRef]
- Rafiqi, M.; Ellis, J.G.; Ludowici, V.A.; Hardham, A.R.; Dodds, P.N. Challenges and progress towards understanding the role of effectors in plant–fungal interactions. Curr. Opin. Plant Biol. 2012, 15, 477–482. [Google Scholar] [CrossRef]
- Schulze-Lefert, P. Knocking on the heaven’s wall: Pathogenesis of and resistance to biotrophic fungi at the cell wall. Curr. Opin. Plant Biol. 2004, 7, 377–383. [Google Scholar] [CrossRef]
- von Röpenack, E.; Parr, A.; Schulze-Lefert, P. Structural analyses and dynamics of soluble and cell wall-bound phenolics in a broad spectrum resistance to the powdery mildew fungus in barley. J. Biol. Chem. 1998, 273, 9013–9022. [Google Scholar] [CrossRef] [Green Version]
- Olivares-García, C.A.; Mata-Rosas, M.; Peña-Montes, C.; Quiroz-Figueroa, F.; Segura-Cabrera, A.; Shannon, L.M.; Loyola-Vargas, V.M.; Monribot-Villanueva, J.L.; Elizalde-Contreras, J.M.; Ibarra-Laclette, E.; et al. Phenylpropanoids are connected to cell wall fortification and stress tolerance in avocado somatic embryogenesis. Int. J. Mol. Sci. 2020, 21, 5679. [Google Scholar] [CrossRef]
- Hoech, K.; Koopmann, B.; von Tiedemann, A. Lignin composition and timing of cell wall lignification are involved in brassica napus resistance to stem rot caused by Sclerotinia sclerotiorum. Phytopathology 2021, 111, 1438–1448. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Xu, W.; Zeng, J.; Li, S.; Li, L.; Gao, Z. Bacillus pumilus LZP02 promotes rice root growth by improving carbohydrate metabolism and phenylpropanoid biosynthesis. Mol. Plant Microbe Interact. 2020, 33, 1222–1231. [Google Scholar] [CrossRef]
- Carotenuto, G.; Sciascia, I.; Oddi, L.; Volpe, V.; Genre, A. Size matters: Three methods for estimating nuclear size in mycorrhizal roots of Medicago truncatula by image analysis. BMC Plant Biol. 2019, 19, 1–14. [Google Scholar] [CrossRef]
- Kou, M.-Z.; Bastías, D.; Christensen, M.; Zhong, R.; Nan, Z.-B.; Zhang, X.-X. The plant salicylic acid signalling pathway regulates the infection of a biotrophic pathogen in grasses associated with an Epichloë endophyte. J. Fungi 2021, 7, 633. [Google Scholar] [CrossRef]
- Zhao, Z.; Kou, M.; Zhong, R.; Xia, C.; Christensen, M.J.; Zhang, X. Transcriptome analysis revealed plant hormone biosynthesis and response pathway modification by Epichloë gansuensis in Achnatherum inebrians under different soil moisture availability. J. Fungi 2021, 7, 640. [Google Scholar] [CrossRef]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of arabidopsis auxin repsonse factor17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Hassan, M.J.; Li, Z.; Peng, Y. Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Lin, H. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Spore Concentrations of Strain ND35 (Spores/mL) | Seed Germination Stage | Seedling Stage | |||||
|---|---|---|---|---|---|---|---|---|
| Radicle Length (mm) | Fresh Weight of Radicle (g) | Dry Weight of Radicle (g) | Plant Height(cm) | Root Length (cm) | Shoot dry Weight (g) | Root Dry Weight (g) | ||
| CK | 0 | 18.31 ± 4.68 c | 0.108 ± 0.025 c | 0.076 ± 0.012 d | 16.46 ± 2.47 cd | 8.49 ± 1.28 a | 0.42 ± 0.045 bc | 0.047 ± 0.014 b |
| T1 | 1 × 104 | 19.92 ± 6.84 c | 0.113 ± 0.009 c | 0.094 ± 0.004 c | 17.31 ± 1.46 cd | 7.15 ± 1.34 c | 0.41 ± 0.043 bc | 0.057 ± 0.006 b |
| T2 | 1 × 105 | 25.41 ± 5.49 b | 0.402 ± 0.009 b | 0.154 ± 0.004 b | 19.36 ± 1.96 a | 6.64 ± 0.63 c | 0.47 ± 0.036 ab | 0.060 ± 0.018 ab |
| T3 | 1 × 106 | 30.56 ± 3.57 a | 0.593 ± 0.039 a | 0.258 ± 0.007 a | 18.78 ± 1.38 ab | 8.77 ± 1.40 a | 0.49 ± 0.036 a | 0.079 ± 0.008 a |
| T4 | 1 × 107 | 16.51 ± 3.23 c | 0.089 ± 0.009 c | 0.057 ± 0.008 e | 15.94 ± 1.63 d | 8.68 ± 1.11 a | 0.48 ± 0.033 a | 0.066 ± 0.004 ab |
| T5 | 1 × 108 | 17.79 ± 7.76 c | 0.076 ± 0.017 c | 0.051 ± 0.007 e | 17.69 ± 1.46 bc | 8.19 ± 1.52 ab | 0.36 ± 0.030 cd | 0.060 ± 0.010 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Fu, X.; Zhang, G.; Zhang, R.; Kang, Z.; Gao, K.; Mendgen, K. Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35. J. Fungi 2022, 8, 180. https://doi.org/10.3390/jof8020180
Tian Y, Fu X, Zhang G, Zhang R, Kang Z, Gao K, Mendgen K. Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35. Journal of Fungi. 2022; 8(2):180. https://doi.org/10.3390/jof8020180
Chicago/Turabian StyleTian, Yehan, Xuesong Fu, Gongchen Zhang, Rui Zhang, Zhensheng Kang, Kexiang Gao, and Kurt Mendgen. 2022. "Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35" Journal of Fungi 8, no. 2: 180. https://doi.org/10.3390/jof8020180
APA StyleTian, Y., Fu, X., Zhang, G., Zhang, R., Kang, Z., Gao, K., & Mendgen, K. (2022). Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35. Journal of Fungi, 8(2), 180. https://doi.org/10.3390/jof8020180