
Identification and Characterization of Auxin/IAA Biosynthesis Pathway in the Rice Blast Fungus Magnaporthe oryzae

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Growth Conditions, and Genetic Transformation
2.2. Blast Infection Assay
2.3. Chemical Reagents Used in This Study
2.4. Plasmid Constructs and Fungal Transformants
2.5. Nucleus Acid Manipulation
2.6. Tryptophan Metabolomics Analysis
2.7. Extraction and Detection of IAA by LC-MS
2.8. Data Analysis
3. Results
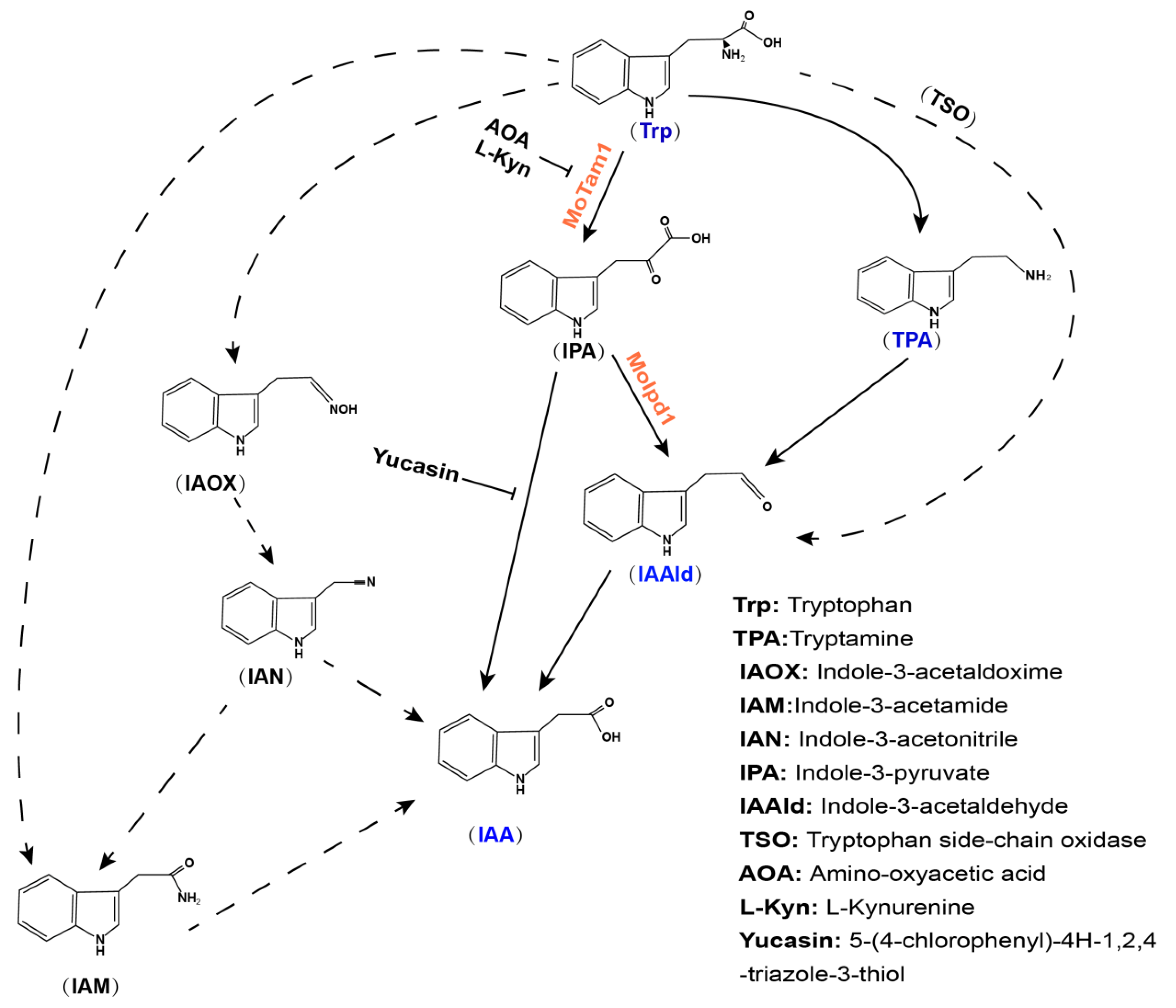
3.1. Identification of Tryptophan-Dependent IAA Synthesis Pathway in M. oryzae

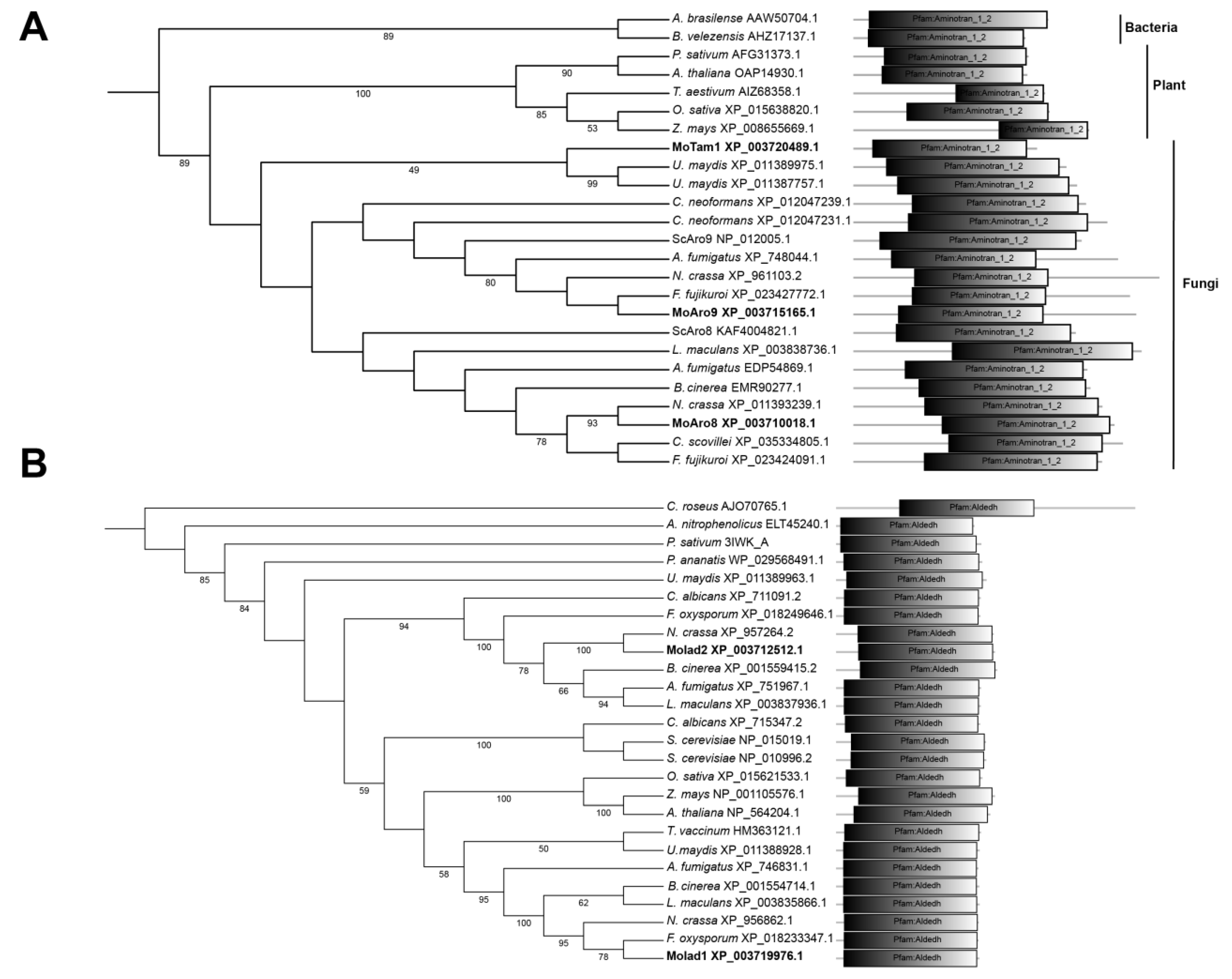{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme [Reference] | M. oryzae ID | Bait | Identity (%) | E Value |
|---|---|---|---|---|
| TPA pathway | ||||
| TDC [21] | MGG_03869 | CAA47898.1 | 25.63 | 8 × 10−24 |
| AOX [5] | MGG_10751 | P49250.1 | 30.63 | 7 × 10-78 |
| IPA pathway | ||||
| TAM [8] | MGG_14221 | XP_011387757.1 | 39.81 | 4 × 10−98 |
| MGG_08189 | 24.91 | 4 × 10−31 | ||
| MGG_09919 | 27.20 | 1 × 10−47 | ||
| IPDC [10] | MGG_01892 | XP_003844157.1 | 30.00 | 4 × 10−70 |
| IAD [9] | MGG_03900 | XP_011388928.1 | 34.18 | 1 × 10−90 |
| MGG_05008 | XP_011389876.1 | 34.02 | 2 × 10−88 | |
| YUCs [22] | MGG_07629 | NP_194980.1 (YUC1) | 27.00 | 4 × 10−32 |
| NP_193062.1 (YUC2) | 25.07 | 5 × 10−24 | ||
| NP_171955.1 (YUC3) | 29.53 | 2 × 10−31 | ||
| NP_196693.1 (YUC4) | 28.15 | 7 × 10−30 | ||
| NP_199202.1 (YUC5) | 26.10 | 5 × 10−28 | ||
| NP_197944.2 (YUC6) | 25.15 | 2 × 10−22 | ||
| NP_180881.1 (YUC7) | 27.41 | 1 × 10−27 | ||
| NP_194601.1 (YUC8) | 27.78 | 4 × 10−31 | ||
| NP_171914.1 (YUC9) | 28.82 | 9 × 10−32 | ||
| MGG_04751 | NP_175321.1 (YUC10) | 25.49 | 6 × 10−14 | |
| MGG_07629 | NP_173564.1 (YUC11) | 28.82 | 1 × 10−30 | |
| IAM pathway | ||||
| TR2M [23] | MGG_16373 | AAD30489.1 | 39.70 | 1 × 10−6 |
| IAM hydrolase [24] | MGG_15383 | AEX60887.1 | 32.07 | 7 × 10−63 |
| IAN pathway | ||||
| TPH [25] | MGG_03352 | OAP11515.1 | 25.05 | 6 × 10−30 |
| IAOx dehydratase [26] | MGG_14859 | OAP11700 | 24.36 | 2 × 10−27 |
| Nhase [27] | No hit | AFY70185.1 | ||
| NIT [28] | MGG_03280 | KIS68359.1 | 33.75 | 7 × 10−53 |
| TSO pathway | ||||
| TSO [29] | MGG_16230 | SCZ74856.1 | 38.68 | 4 × 10−97 |

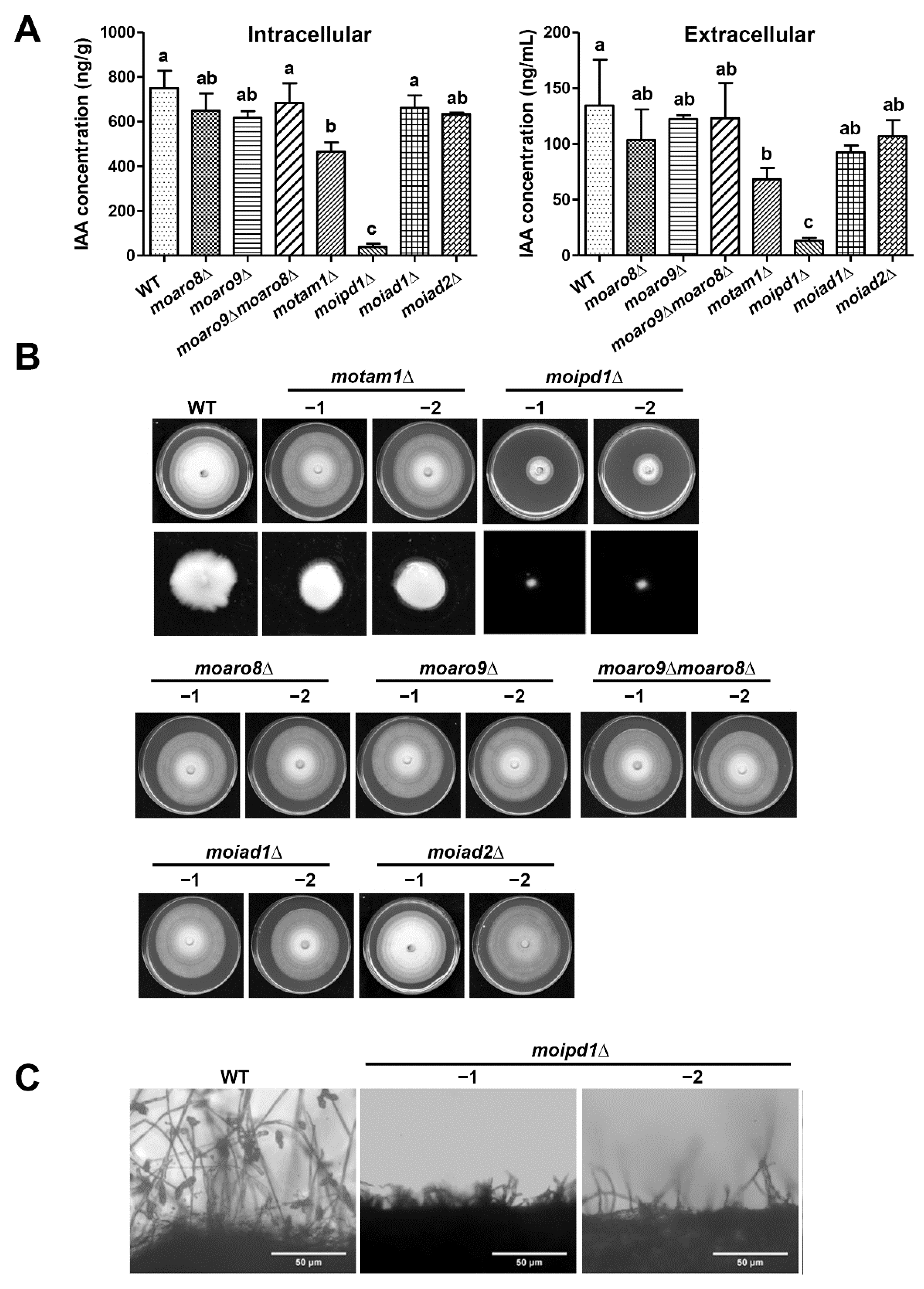
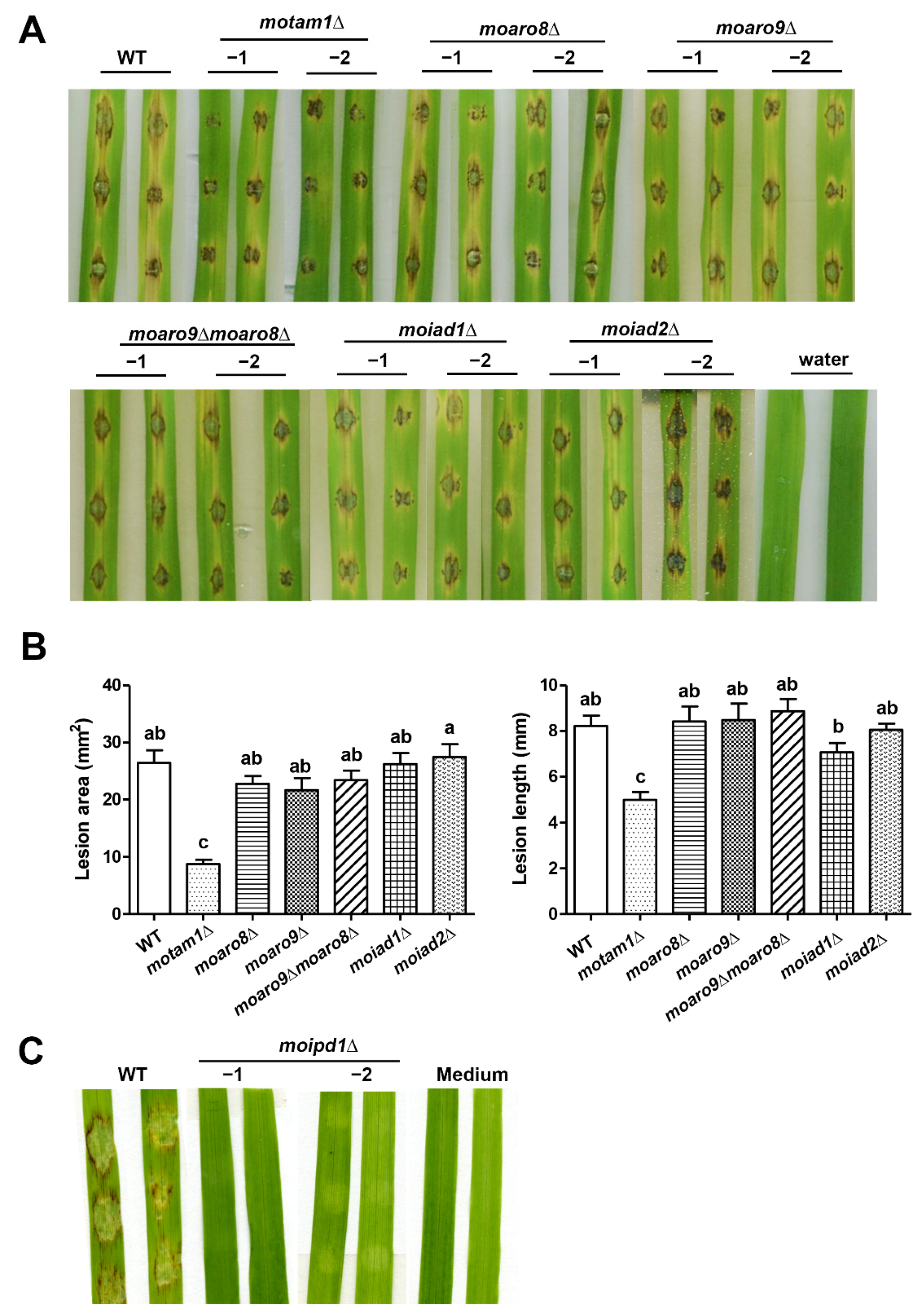
3.2. MoTam1 and MoIpd1 Regulate Mycelial Growth, Conidiation, and Pathogenicity of M. oryzae
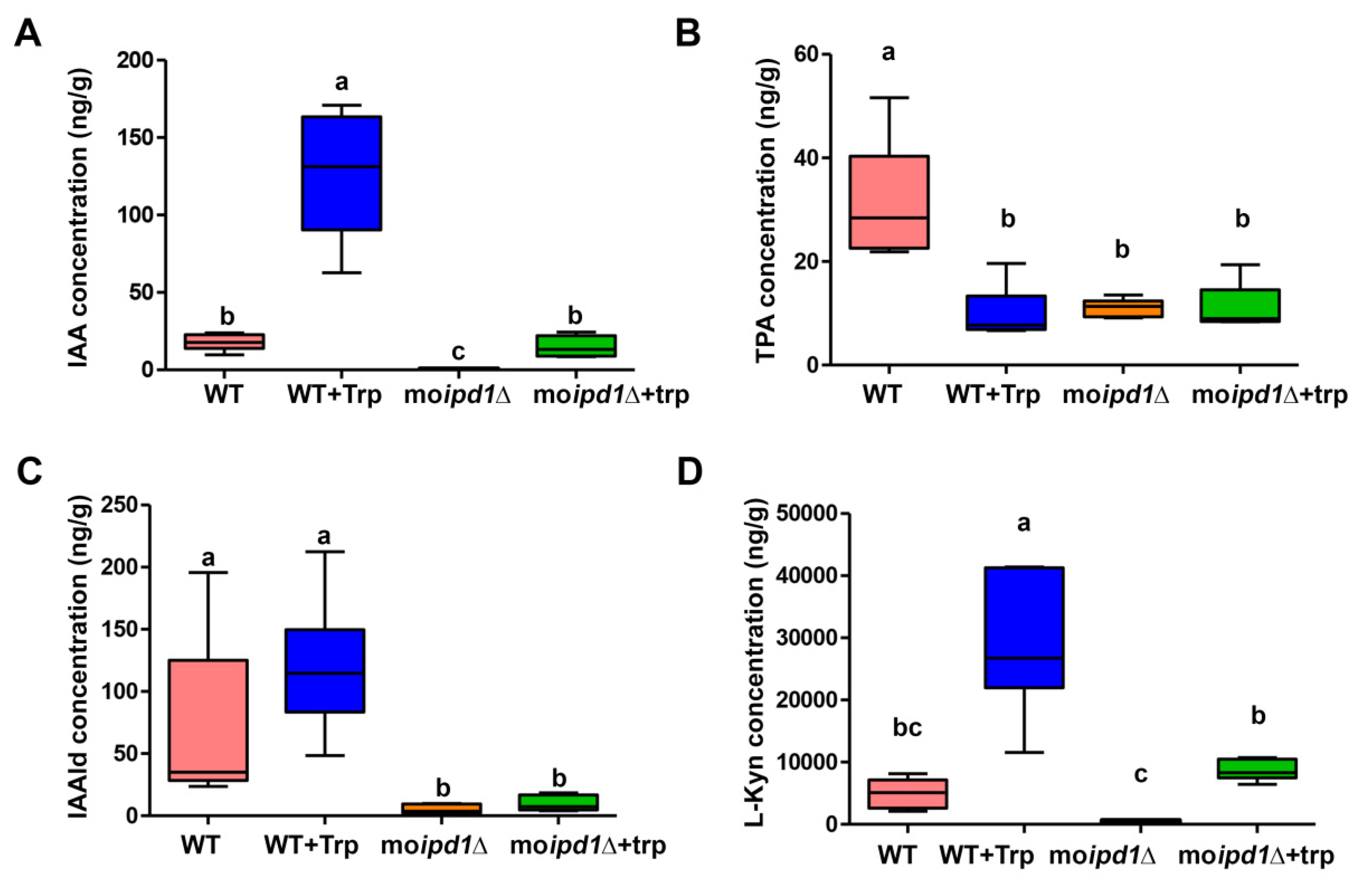
3.3. The IPA Pathway Is A Major Contributor of IAA Biosynthesis in M. oryzae
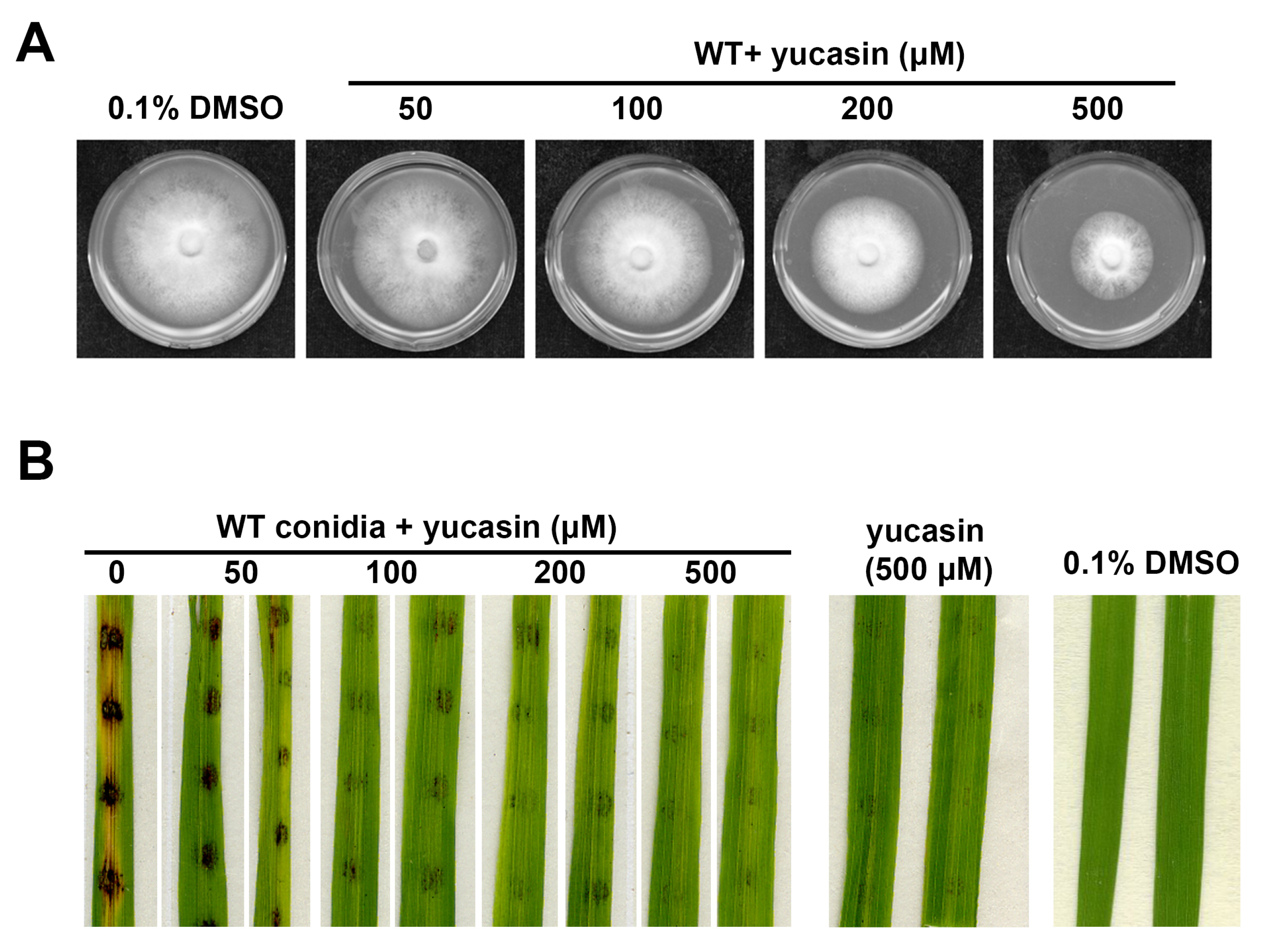
3.4. YUCCA Pathway May Also Contribute to M. oryzae Growth, Conidiation, and Pathogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [Green Version]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, G.B.; Sanjeevkumar, S.; Kirankumar, B.; Santoshkumar, M.; Karegoudar, T.B. Indole-3-acetic acid biosynthesis in Fusarium delphinoides strain GPK, a causal agent of wilt in chickpea. Appl. Biochem. Biotechnol. 2013, 169, 1292–1305. [Google Scholar] [CrossRef] [PubMed]
- Kumla, J.; Suwannarach, N.; Matsui, K.; Lumyong, S. Biosynthetic pathway of indole-3-acetic acid in ectomycorrhizal fungi collected from northern Thailand. PLoS ONE 2020, 15, e0227478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuther, K.; Mayser, P.; Hettwer, U.; Wu, W.; Spiteller, P.; Kindler, B.L.J.; Karlovsky, P.; Basse, C.W.; Schirawski, J. The tryptophan aminotransferase Tam1 catalyses the single biosynthetic step for tryptophan-dependent pigment synthesis in Ustilago maydis. Mol. Microbiol. 2008, 68, 152–172. [Google Scholar] [CrossRef]
- Basse, C.W.; Lottspeich, F.; Steglich, W.; Kahmann, R. Two potential indole-3-acetaldehyde dehydrogenases in the phytopathogenic fungus Ustilago maydis. Eur. J. Biochem. 1996, 242, 648–656. [Google Scholar] [CrossRef]
- Leontovyčová, H.; Trdá, L.; Dobrev, P.I.; Šašek, V.; Gay, E.; Balesdent, M.H.; Burketová, L. Auxin biosynthesis in the phytopathogenic fungus Leptosphaeria maculans is associated with enhanced transcription of indole-3-pyruvate decarboxylase LmIPDC2 and tryptophan aminotransferase LmTAM1. Res. Microbiol. 2020, 171, 174–184. [Google Scholar] [CrossRef]
- Talbot, N.J. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea. Annu. Rev. Microbiol. 2003, 57, 177–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamer, J.E.; Howard, R.J.; Chumley, F.G.; Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. Science 1988, 239, 288–290. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Liu, H.; Li, Y.; Yu, H.; Li, X.; Xiao, J.; Wang, S. Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice. Plant Physiol. 2011, 155, 589–602. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.J.; Shimono, M.; Sugano, S.; Kojima, M.; Liu, X.; Inoue, H.; Sakakibara, H.; Takatsuji, H. Cytokinins act synergistically with salicylic acid to activate defense gene expression in rice. Mol. Plant-Microbe Interact. 2013, 26, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rho, H.S.; Kang, S.; Lee, Y.H. Agrobacterium tumefaciens-mediated transformation of the plant pathogenic fungus, Magnaporthe grisea. Mol. Cells 2001, 12, 407–411. [Google Scholar]
- Dong, L.H.; Shen, Q.; Chen, C.-Y.; Shen, L.Z.; Yang, F.; Naweed, I.N.; Deng, Y.Z. Fungal auxin is a quorum-based modulator of blast disease severity. bioRxiv 2021, 10, 29–33. [Google Scholar] [CrossRef]
- Feng, J.; Hwang, R.; Chang, K.F.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; Zhou, Q. An inexpensive method for extraction of genomic DNA from fungal mycelia. Can. J. Plant Pathol. 2010, 32, 396–401. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Hazelwood, L.A.; Daran, J.M.; Van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, J.; Adachi, T.; Hidaka, H. Purification and characterization of indolepyruvate decarboxylase: A novel enzyme for indole-3-acetic acid biosynthesis in Enterobacter cloacae. J. Biol. Chem. 1992, 267, 15823–15828. [Google Scholar] [CrossRef]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The roles of auxin biosynthesis YUCCA gene family in plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [Green Version]
- Van Onckelen, H.; Prinsen, E.; Inzé, D.; Rüdeisheim, P.; Van Lijsebettens, M.; Follin, A.; Schell, J.; Van Montagu, M.; De Greef, J. Agrobacterium T-DNA gene 1 codes for tryptophan 2-monooxygenase activity in tobacco crown gall cells. FEBS Lett. 1986, 198, 357–360. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Oeser, B.; Oren-Young, L.; Israeli, M.; Sasson, Y.; Tudzynski, B.; Sharon, A. Identification and functional characterization of indole-3-acetamide-mediated IAA biosynthesis in plant-associated Fusarium species. Fungal Genet. Biol. 2012, 49, 48–57. [Google Scholar] [CrossRef]
- Hull, A.K.; Vij, R.; Celenza, J.L. Arabidopsis cytochrome P450s that catalyze the first step of tryptophan-dependent indole-3-acetic acid biosynthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2379–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nafisi, M.; Goregaoker, S.; Botanga, C.J.; Glawischnig, E.; Olsen, C.E.; Halkier, B.A.; Glazebrook, J. Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-3-acetaldoxime in camalexin synthesis. Plant Cell 2007, 19, 2039–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duca, D.; Rose, D.R.; Glick, B.R. Characterization of a nitrilase and a nitrile hydratase from Pseudomonas sp. strain UW4 that converts indole-3-acetonitrile to indole-3-acetic acid. Appl. Environ. Microbiol. 2014, 80, 4640–4649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reineke, G.; Heinze, B.; Schirawski, J.; Buettner, H.; Kahmann, R.; Basse, C.W. Indole-3-acetic acid (IAA) biosynthesis in the smut fungus Ustilago maydis and its relevance for increased IAA levels in infected tissue and host tumour formation. Mol. Plant Pathol. 2008, 9, 339–355. [Google Scholar] [CrossRef]
- Ai, Y.; Wang, B.; Xiao, S.; Luo, S.; Wang, Y. Tryptophan side-chain oxidase enzyme suppresses hepatocellular carcinoma growth through degradation of tryptophan. Int. J. Mol. Sci. 2021, 22, 12428. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the Bootstrap. Soc. Study Evol. Stable 1985, 39, 783–791. [Google Scholar]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. Evol. Genes Proteins 1965, 97–166. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Perley, J.E.; Stowe, B.B. On the ability of Taphrina deformans to produce indoleacetic acid from tryptophan by way of tryptamine. Plant Physiol. 1966, 41, 234–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberhansli, T.; Defago, G.; Haas, D. Indole-3-acetic acid (IAA) synthesis in the biocontrol strain CHA0 of Pseudomonas fluorescens Role of tryptophan side chain oxidase. J. Gen. Microbiol. 1991, 137, 2273–2279. [Google Scholar] [CrossRef] [Green Version]
- Zelante, T.; Choera, T.; Beauvais, A.; Fallarino, F.; Paolicelli, G.; Pieraccini, G.; Pieroni, M.; Galosi, C.; Beato, C.; De Luca, A.; et al. Aspergillus fumigatus tryptophan metabolic route differently affects host immunity. Cell Rep. 2021, 34, 108673. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A small-molecule screen identifies L-Kynurenine as a competitive inhibitor of TAA1/TAR activity in ethylene-directed auxin biosynthesis and root growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, T.; Hayashi, K.I.; Suzuki, H.; Gyohda, A.; Takaoka, C.; Sakaguchi, Y.; Matsumoto, S.; Kasahara, H.; Sakai, T.; Kato, J.I.; et al. Yucasin is a potent inhibitor of YUCCA, a key enzyme in auxin biosynthesis. Plant J. 2014, 77, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Frankenberger, W.T. Microbial production of plant hormones. Plant Soil 1991, 133, 1–8. [Google Scholar] [CrossRef]
- Ishii, T.; Soeno, K.; Asami, T.; Fujioka, S.; Shimada, Y. Arabidopsis seedlings over-accumulated indole-3-acetic acid in response to aminooxyacetic acid. Biosci. Biotechnol. Biochem. 2010, 74, 2345–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, L.; Zheng, L.; Sheng, C.; Zhao, H.; Jin, H.; Niu, D. Rice siR109944 suppresses plant immunity to sheath blight and impacts multiple agronomic traits by affecting auxin homeostasis. Plant J. 2020, 102, 948–964. [Google Scholar] [CrossRef]
- Qin, Q.; Li, G.; Jin, L.; Huang, Y.; Wang, Y.; Wei, C.; Xu, Z.; Yang, Z.; Wang, H.; Li, Y. Auxin response factors (ARFs) differentially regulate rice antiviral immune response against rice dwarf virus. PLoS Pathog. 2020, 16, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Yang, Y.; He, Z. Roles of plant hormones and their interplay in rice immunity. Mol. Plant 2013, 6, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djami-Tchatchou, A.T.; Harrison, G.A.; Harper, C.P.; Wang, R.; Prigge, M.J.; Estelle, M.; Kunkel, B.N. Dual role of auxin in regulating plant defense and bacterial virulence gene expression during Pseudomonas syringae PtoDC3000 pathogenesis. Mol. Plant-Microbe Interact. 2020, 33, 1059–1071. [Google Scholar] [CrossRef]
- Fan, S.; Chang, Y.; Liu, G.; Shang, S.; Tian, L.; Shi, H. Molecular functional analysis of auxin/indole-3-acetic acid proteins (Aux/IAAs) in plant disease resistance in cassava. Physiol. Plant. 2020, 168, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Andrés, F.; Tharreau, D.; Iglesias, D.J.; Talón, M. Constitutive expression of OsGH3.1 reduces auxin content and enhances defense response and resistance to a fungal pathogen in rice. Mol. Plant-Microbe Interact. 2009, 22, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, X.; Long, J.; Zhao, K.; Peng, A.; Chen, M.; Long, Q.; He, Y.; Chen, S. Overexpressing GH3.1 and GH3.1L reduces susceptibility to Xanthomonas citri subsp. Citri by repressing auxin signaling in citrus (Citrus sinensis Osbeck). PLoS ONE 2019, 14, e0220017. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.X.; Feng, Q.; Cao, X.L.; Zhu, Y.; Wang, H.; Chandran, V.; Fan, J.; Zhao, J.Q.; Pu, M.; Li, Y.; et al. Osa-miR167d facilitates infection of Magnaporthe oryzae in rice. J. Integr. Plant Biol. 2020, 62, 702–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, R.N.; Chattopadhyay, K.K.; Basu, P.S. Auxin activity of 3-hydroxymethyl oxindole and 3-methylene oxindole in oat. Biol. Plant 1986, 28, 219–226. [Google Scholar] [CrossRef]
- Fukuyama, T.T.; Moyed, H.S. Inhibition of cell growth by photooxidation products of indole-3-acetic. J. Biol. Chem. 1964, 239, 2392–2397. [Google Scholar] [CrossRef]
- Goldacre, P.L. The photochemical inactivation of indoleacetic acid sensitized by non-protein components of plant tissues. Aust. J. Biol. Sci. 1954, 7, 225–250. [Google Scholar] [CrossRef] [Green Version]





| Strain | Radius of Colony (Average ± S.E.) | Conidia Count (×105/mL; Average ± S.E.) |
|---|---|---|
| Wild type (WT) | 3.52 ± 0.02 A | 58.76 ± 5.08 a |
| motam1Δ | 3.53 ± 0.07 A | 60.58 ± 4.10 a |
| moaro8Δ | 3.58 ± 0.02 A | 65.05 ± 1.89 a |
| moaro9Δ | 3.50 ± 0.02 A | 63.25 ± 7.76 a |
| moaro9Δ moaro8Δ | 3.60 ± 0.04 A | 59.20 ± 6.71 a |
| moipd1Δ | 1.42 ± 0.03 D | 0 c |
| moiad1Δ | 3.57 ± 0.03 A | 60.32 ± 6.35 a |
| moiad2Δ | 3.52 ± 0.05 A | 61 ± 5.10 a |
| WT + Yucasin (50 μM) | 3.42 ± 0.03 A | 17.42 ± 4.49 b |
| (100 μM) | 3.05 ± 0.02 B | 13.55 ± 2.37 b |
| (200 μM) | 2.63 ± 0.03 C | 8.71 ± 2.37 bc |
| (500 μM) | 1.72 ± 0.07 D | 6.78 ± 1.19 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Ma, Y.; Chen, C.-Y.; Shen, L.; Sun, W.; Cui, G.; Naqvi, N.I.; Deng, Y.Z. Identification and Characterization of Auxin/IAA Biosynthesis Pathway in the Rice Blast Fungus Magnaporthe oryzae. J. Fungi 2022, 8, 208. https://doi.org/10.3390/jof8020208
Dong L, Ma Y, Chen C-Y, Shen L, Sun W, Cui G, Naqvi NI, Deng YZ. Identification and Characterization of Auxin/IAA Biosynthesis Pathway in the Rice Blast Fungus Magnaporthe oryzae. Journal of Fungi. 2022; 8(2):208. https://doi.org/10.3390/jof8020208
Chicago/Turabian StyleDong, Lihong, Yuming Ma, Cheng-Yen Chen, Lizheng Shen, Wenda Sun, Guobing Cui, Naweed I. Naqvi, and Yi Zhen Deng. 2022. "Identification and Characterization of Auxin/IAA Biosynthesis Pathway in the Rice Blast Fungus Magnaporthe oryzae" Journal of Fungi 8, no. 2: 208. https://doi.org/10.3390/jof8020208
APA StyleDong, L., Ma, Y., Chen, C. -Y., Shen, L., Sun, W., Cui, G., Naqvi, N. I., & Deng, Y. Z. (2022). Identification and Characterization of Auxin/IAA Biosynthesis Pathway in the Rice Blast Fungus Magnaporthe oryzae. Journal of Fungi, 8(2), 208. https://doi.org/10.3390/jof8020208