3.2. Taxonomy
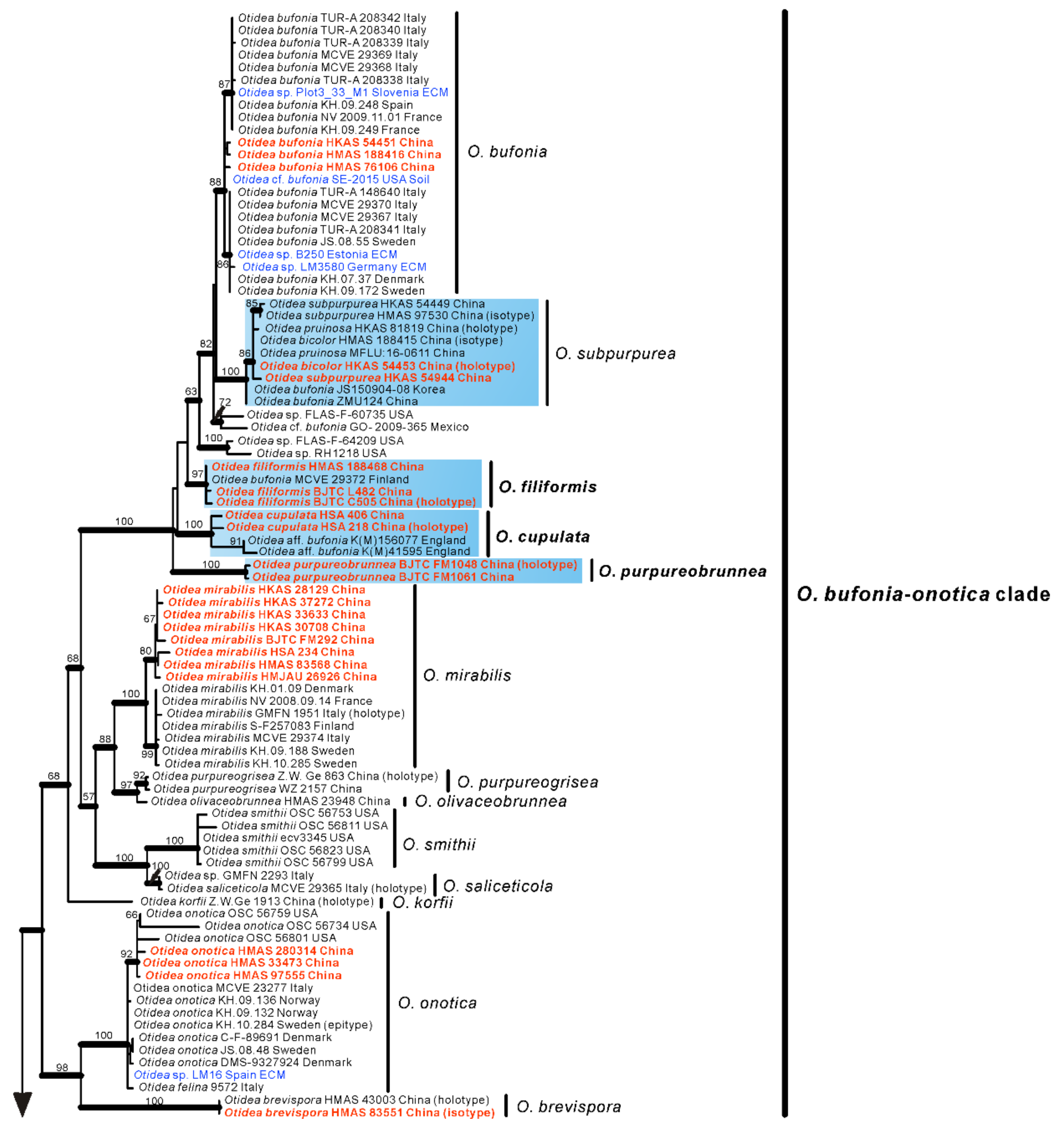
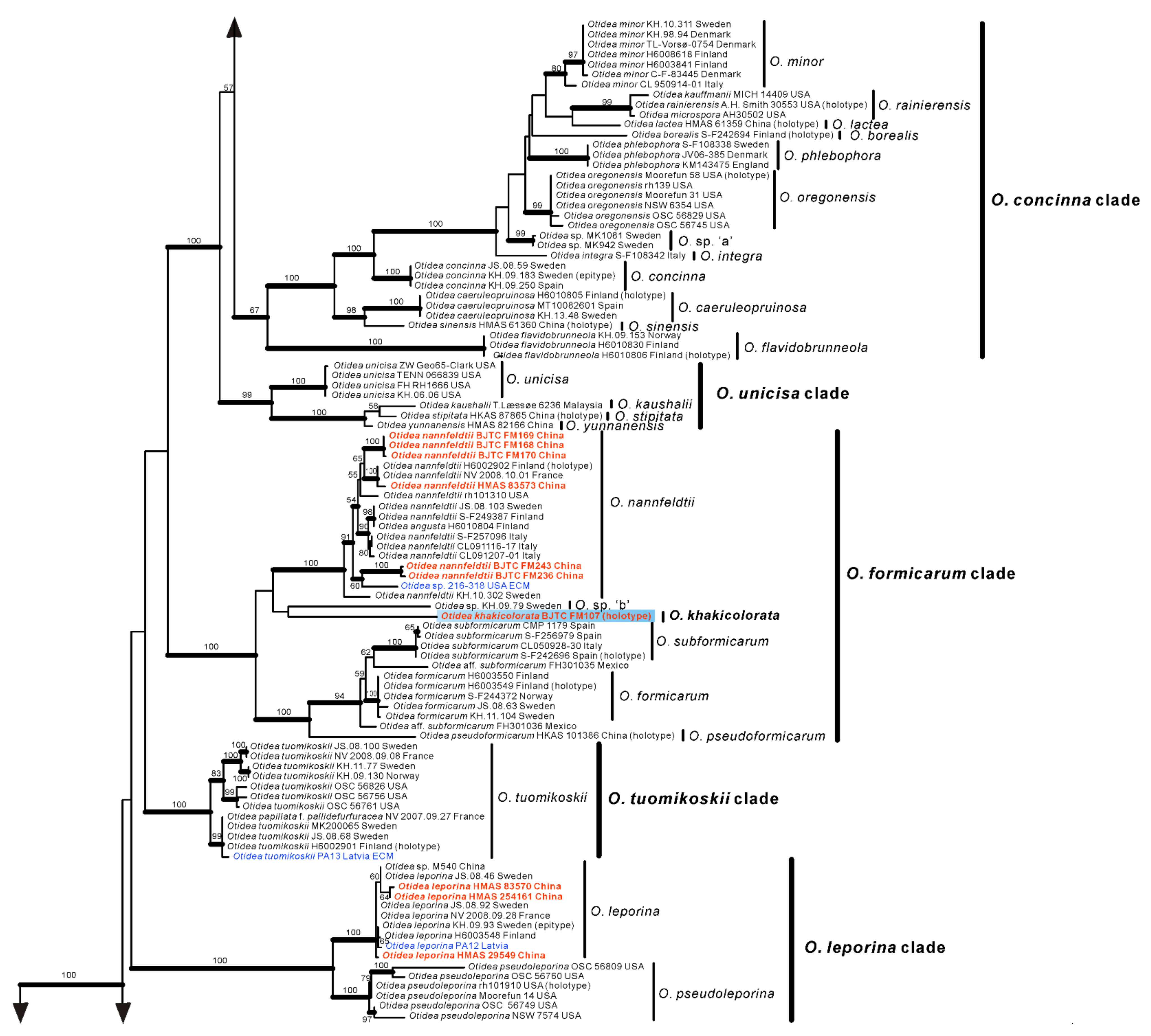
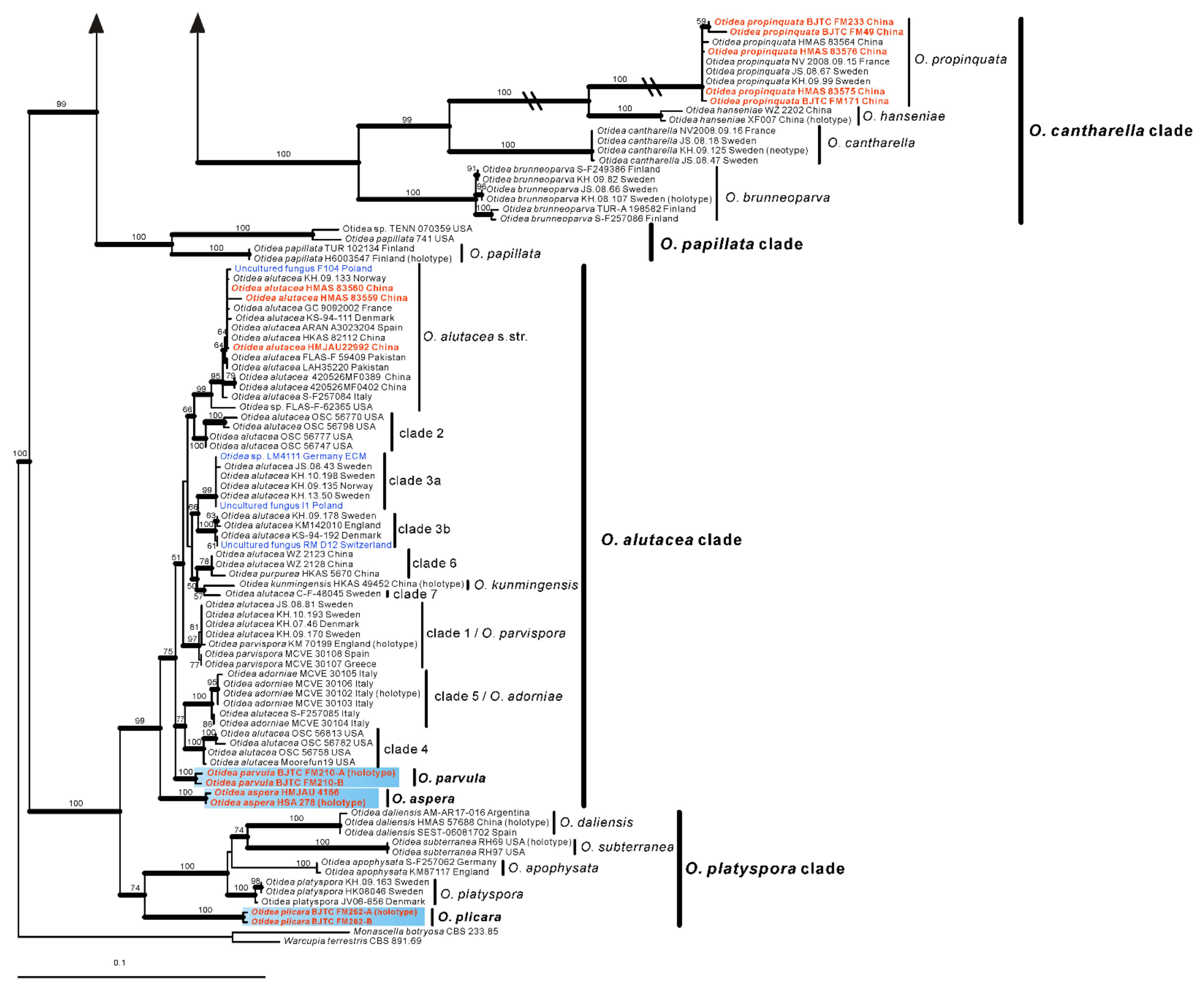
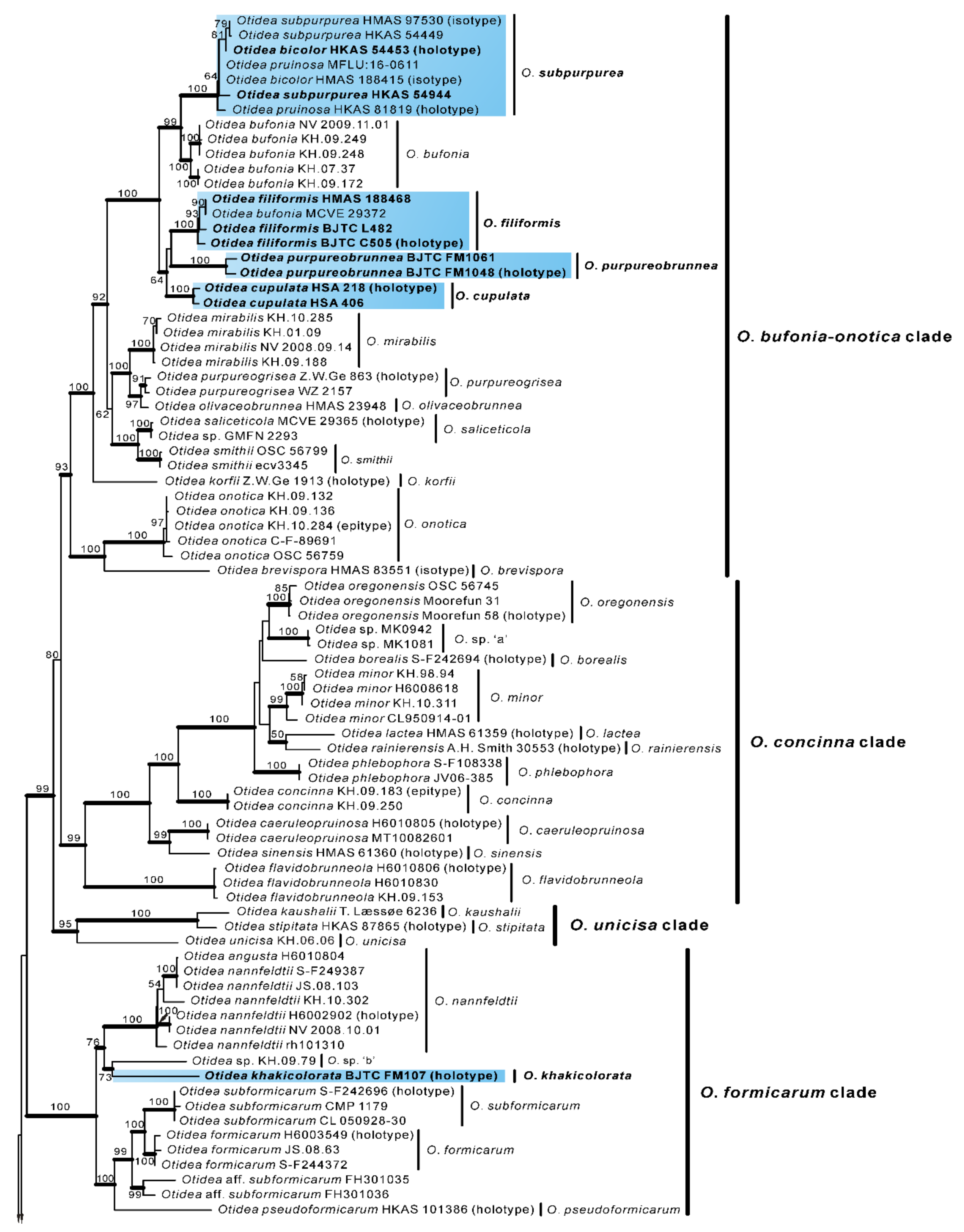
Based on our phylogenies and morphological data, a total of seven new species, a known species, and two new records of Otidea from China were described and illustrated here.
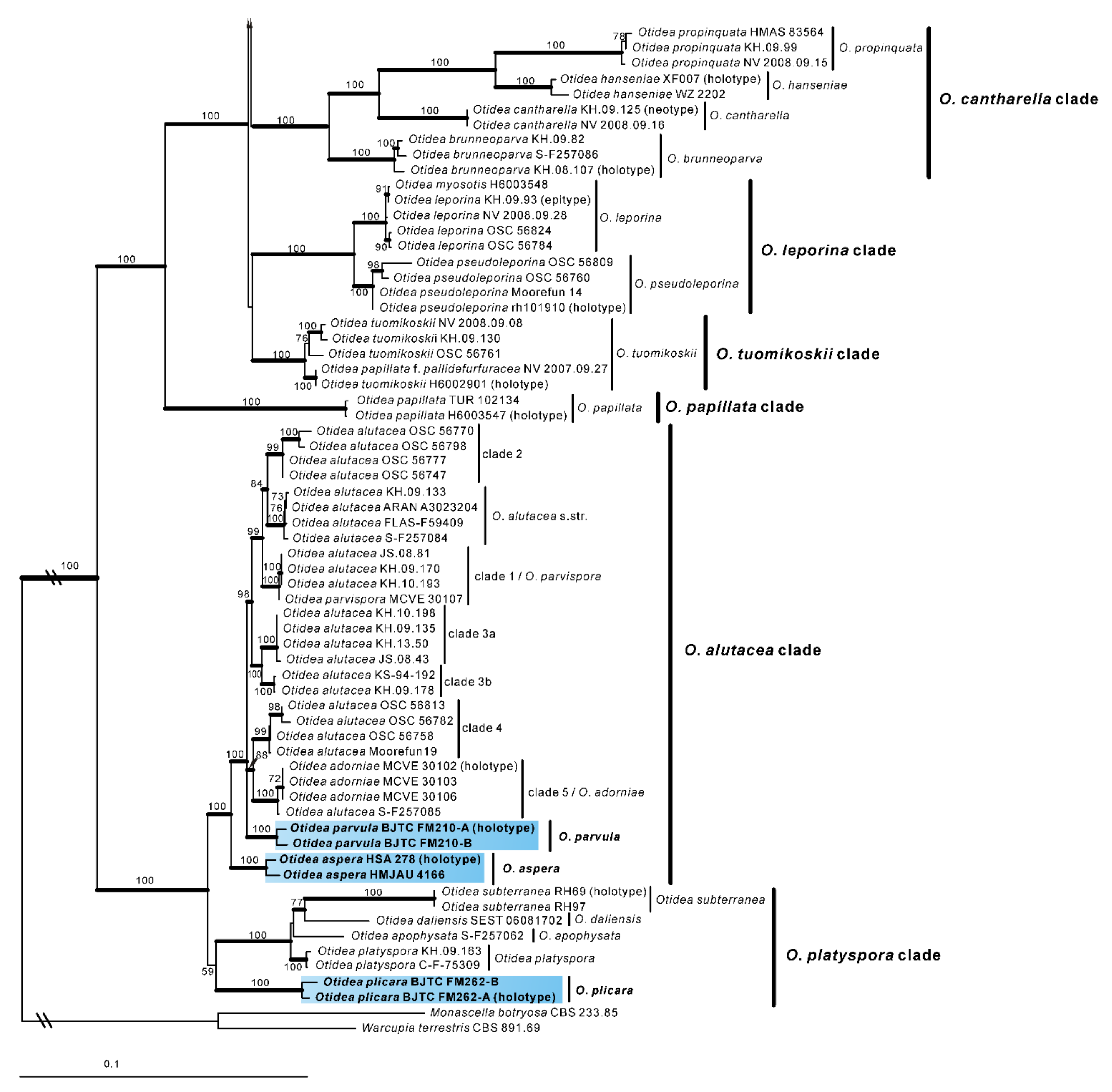
Otideaaspera L. Fan & Y.Y. Xu, sp. nov. (
Figure 3).
MycoBank: MB843176.
Etymology: aspera, referring to the rough receptacle surface.
Holotype: China. Shanxi Province, Jiaocheng County, Pangquangou National Nature Reserve, HaoJiagou, alt. 1800m, in the mixed forest dominated by Pinus tabuliformis Carrière and Quercus mongolica Fisch. ex Ledeb., 29 August 2018, J.Z. Cao, LH278 (HSA 278).
Saprobic on soil. Apothecia solitary or caespitose in nature, 30–45 mm high, 25–60 mm wide, initially ear-shaped, then expanding and sometimes becoming irregularly ear-shaped or shallowly to deeply cup-shaped, sometimes elongated on one side or obconical, split, stipitate. Hymenium surface greyish yellow (#fbe8c5) to light brown (#baab98) when fresh, ochre brown (#a87832) when dry, subsmooth. Receptacle surface pale yellow (#fafad2) to yellowish brown (#b5a27f) when fresh, slightly hygrophanous, pale whitish ochre (#c8b99f) when dry, finely warty. Stipe 5–12 × 4–8 mm. Basal tomentum and mycelium white. Apothecial section 600–900 µm thick. Ectal excipulum of textura angularis, 100–150 µm thick, cells thin walled, hyaline to pale brown, 11–28 × 8–20 µm. Medullary excipulum of textura intricata, 300–500 µm thick, hyphae 3–10 µm wide, sometimes slightly swollen, thin to thick walled, septate, hyaline to light brown. Subhymenium c. 50–120 µm thick, visible as a brown zone of densely arranged cylindrical to swollen cells. Paraphyses septate, straight to slightly curved, of uniform width or slightly enlarged at the apices to 3–4.7 µm wide, without or rarely with 1–2 low notches. Asci 150–200 × 9–13 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores ellipsoid to slightly subfusoid, inequilateral, with two large guttules, sometimes only with one big guttule, smooth, hyaline, (12–) 12.8–15 (–15.5) × (5.8–) 6.5–7.5 (–8) µm (Lm × Wm = 14 × 7 µm, Q = 1.8–2.2, Qm = 2, n = 50). Receptacle surface with warts, 50–80 µm high, formed by short, fasciculate, hyphoid hairs, of 5–7 subglobose to elongated cells, constricted at septa, 6–13 µm wide. Resinous exudates absent to scarce. Basal mycelium of 2–5 µm wide, septate, hyaline to pale brown hyphae, smooth, unchanged in KOH, smooth, turning yellow in MLZ.
Other materials examined: China. Inner Mongolia Autonomous Region, Daqinggou National Nature Reserve, on the broad-leaved woodland, 24 August 2005, Tolgor Bau (HMJAU 4166).
Notes:
Otidea aspera is diagnosed by the combination of the stipitate, broadly ear shaped to cup-shaped, greyish yellow to light brown hymenium, pale-yellow to yellowish-brown receptacle surface, ellipsoid to slightly subfusoid ascospores and straight to slightly curved paraphyses.
Otidea aspera and
O. parvispora have comparable apothecia color, however
O. parvispora differs from
O. aspera by the smaller ascospores ((11.0–) 11.5–13.0 × 5.0–6.5 µm) and shorter asci. DNA analysis showed that
O. aspera shared less than 93.39% ITS sequence similarity with other
Otidea species. Phylogenetic analyses revealed that the sequences of
O. aspera were grouped into an independent clade with a strong support value (
Figure 1 and
Figure 2). These supported the erection of the new species.
Otidea cupulata L. Fan & Y.Y. Xu, sp. nov. (
Figure 4).
MycoBank: MB843177.
Etymology: cupulata, referring to the apothecia shape of the fungus.
Holotype: China. Shanxi Province, Jiaocheng County, Pangquangou Township, Badaogou valley, alt. 2200m, on soil in mixed forest of Larix sp. and Betula sp., 28 August 2018, L.J. Guo, LH218 (HSA 218).
Saprobic on soil. Apothecia gregarious or caespitose in nature, 25–40 mm high, 15–50 mm wide, initially ear-shaped, soon expanding, becoming broadly ear-shaped or deeply cup-shaped, split, often broader above, margin sometimes lobate, stipitate. Hymenium surface dark orange brown (#734a12) to dark purple brown (#4d282d) when fresh, and usually with bluish-lilaceous shades, gray ochraceous brown (#37290e) when dry, subsmooth. Receptacle surface yellowish brown (#a1805f) to brown (#8a6660) when fresh, slightly hygrophanous, surface with shallow wrinkles, dark brown (#261600) when dry, furfuraceous. Stipe 5–10 × 5–7 mm. Basal tomentum and mycelium whitish to grayish yellow (#c6cbac). Apothecial section 900–1200 µm thick. Ectal excipulum of textura angularis, 80–110 µm thick, cells thin walled, brownish, 10–25 × 8–22 µm. Medullary excipulum of textura intricata, 500–700 µm thick, hyphae 3.5–10 µm wide, sometimes slightly swollen, thin to slightly thick walled, septate, hyaline to light brown. Subhymenium c. 60–100 µm thick, visible as a brown zone, of densely arranged cylindrical to swollen cells, with scattered brown resinous exudate at septa. Paraphyses septate, curved to hooked of uniform width or slightly enlarged at the apices to 2.6–4.2 µm wide, without or rarely with 1–2 low notches, sometimes forked near the apex. Asci 150–200 × 8–11 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores release from an eccentric split at the apical apex. Ascospores ellipsoid to slightly subfusoid, inequilateral, with two large guttules, sometimes with only one big guttule, smooth, hyaline, (12–) 12.5–15 (–15.5) × (6–) 6.5–7.4 (–7.9) µm (Lm × Wm= 13.9 × 7 µm, Q = 1.9–2.1, Qm = 2, n = 50). Receptacle surface with low warts, 25–45 µm high, formed by short, fasciculate, hyphoid hairs, of 3–4 subglobose to elongated cells, constricted at septa, 6–11 µm wide, sometimes with a gelatinous sheath. Resinous exudates abundant on the outer surface, dark brown, partly dissolving and converting into small particles in MLZ, entirely dissolving and turning bright yellow in KOH. Basal mycelium of 3–6.5 µm wide, septate, hyaline to pale brown hyphae, bright yellow in KOH, with abundant, very small, irregularly, brown, resinous exudates on the surface, dissolving and turning bright yellow in KOH, unchanged in MLZ.
Other materials examined: China. Shanxi Province, Jiaocheng County, Pangquangou Township, Badaogou Valley, alt. 1800m, on soil under Larix sp., 6 September 2018, L.J. Guo, LH 406 (HSA 406).
Notes: Otidea cupulata is recognized by the stipitate, broadly cup-shaped, dark-orange-brown to dark-purple-brown hymenium, yellowish-brown to brown receptacle surface, ellipsoid to subfusoid ascospores, forked or notched paraphyses, and furfuraceous receptacle surface. The forked paraphyses is rarely found in other species in the O. bufonia-onotica clade. Several species in the O. bufonia-onotica clade are similar to O. cupulata in apothecial shape and color, including O. bufonia, O. filiformis, O. mirabilis, and O. olivaceobrunnea Harmaja, but O. bufonia can be distinguished by its distinctly narrowly fusoid ascospores, the presence of hyphae with striate resinous exudates in the medullary excipulum, and resinous exudates of the ectal excipulum that does not turn bright yellow in KOH. Otidea filiformis differs in its apothecia with pinkish shades, distinctly narrowly fusoid ascospores, unenlarged and curved to hooked paraphyses, and higher warts (40–75 µm) on the receptacle surface. Otidea mirabilis differs by having purple to lilaceous-bluish shades on the receptacle surface and narrowly fusoid ascospores (Qm = 2.1–2.3). Otidea olivaceobrunnea can be separated by its olive-brown hymenium and wider ascospores (14–17 × 8–8.5 µm). Four species (O. purpureogrisea Pfister, F. Xu & Z.W. Ge, O. purpureobrunnea, O. simithii Kanouse and O. subpurpurea) have more or less bluish or lilaceous shades on apothecia that resembles that of O. cupulata. However, O. purpureogrisea is distinguished by its ear-shaped apothecia, dark-purple-brown to purple-gray receptacle surface, and the resinous exudate in the ectal excipulum turning amber and brown in KOH. Otidea purpureobrunnea is distinguished by its grayish-purple-brown to dark-purple-brown receptacle surface and mostly smooth basal mycelium. Otidea simithii differs in having typically narrower, ear-shaped apothecia, and resinous exudates of the ectal excipulum that does not turn bright yellow in KOH. Otidea subpurpurea in having smaller ascospores (9–12 × 4.5–6 µm) and lilac to purplish receptacle surface.
In the two-gene phylogenetic tree (
Figure 1), two specimens (K(M)41595 and K(M)156077) from England fall into the clade of
O.
cupulata, which are noted to belong to an unnamed taxon related to
O. bufonia and
O. subpurpurea by Parslow et al. [
10]. Regrettably, they did not further describe the morphology of these two specimens. According to the ITS sequence similarity analysis (ITS: 98%) and phylogenetic analyses (
Figure 1), they may be conspecific with
O. cupulata. Final confirmation requires morphological observation of these two specimens.
Otidea filiformis C.L. Hou, Y.Y. Xu & H. Zhou, sp. nov. (
Figure 5).
MycoBank: MB843178.
Etymology: filiformis, referring to the filiform paraphyses in the hymenium.
Holotype: China. Beijing City, Huairou District, Sunshanzi Village, alt. 770m, on soil in mixed forest of Populus sp. and Larix sp., 28 August 2020, G.Q. Chen C505 (BJTC C505).
Saprobic on soil. Apothecia gregarious or caespitose in nature, 15–55 mm high, 15–55 mm wide, initially ear-shaped, soon expanding, becoming shallowly or deeply cup-shaped, split, margin sometimes lobate, sessile or shortly stipitate, regular or sometimes undulate in the margin. Hymenium surface yellowish brown (#b19461) to ochre yellow (#cd9575) with pinkish shades, sometimes with brown spots or stains when fresh, margin dark brown when bruised, gray brown (#321f15) when dry, subsmooth. Receptacle surface yellowish brown (#b19461) to orange brown (#967059) when fresh, slightly hygrophanous, dark brown (#321f15) when dry, furfuraceous to finely warty. Stipe not well developed. Basal tomentum and mycelium whitish to pale brown (#dccdbf). Apothecial section 800–1000 µm thick. Ectal excipulum of textura angularis, 75–110 µm thick, cells thin walled, brown, 10–35 × 7–26 µm. Medullary excipulum of textura intricata, 400–500 µm thick, hyphae 4–10 µm wide, sometimes slightly swollen, thin to thick walled, septate, hyaline to light brown, without resinous exudates. Subhymenium ca. 75–100 µm thick, visible as a brown zone of densely arranged cylindrical to swollen cells, with scattered brown resinous exudate at septa. Paraphyses septate, curved to hooked of uniform width at the apices to 2–3 µm wide, without or with a low notch. Asci 140–175 × 10–14 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores narrowly fusoid, narrowed at both ends, inequilateral, with two large guttules, sometimes only with one big guttule, smooth, hyaline, (12.5–) 13–14.5 (–15) × (6–) 6.5–7 (–7.3) µm (Lm × Wm = 13.5 × 6.8 µm, Q = 1.9–2.1, Qm = 2, n = 50). Receptacle surface with broad conical warts, 40–75 µm high, formed by short, fasciculate, hyphoid hairs, of 6–7 subglobose to elongated cells, constricted at septa, 6–10 µm wide. Resinous exudates abundant on the outer surface, dark yellow brown, partly dissolving and converting into small particles in MLZ, partially dissolving and turning yellowish brown in KOH. Basal mycelium of interwoven, 3–6 µm wide, septate, hyaline to pale brown hyphae, turning yellow in KOH, with abundant small, regularly arranged, spheroid, pale brown, resinous exudates, partly dissolving in KOH, unchanged in MLZ.
Other materials examined: China. Hebei Province, Chicheng County, Dahaituo nature reserve, alt. 1640m, on soil under Betula platyphylla Suk., 22 August 2019, J.Q. Li L482 (BJTC L482); China. Inner Mongolia Autonomous Region, Balinyou Banner, Saihanwula Nature Reserve, on soil in mixed forest of Larix gmelinii (Rupr.) Kuzen. and Betula platyphylla Suk., 2 September 2008, T.Z. Liu, H.M. Zhou & C. Sun 3858 (HMAS 188468).
Notes: Otidea filiformis diagnosed by the combination of yellowish-brown to ochre-yellow apothecia, sometimes with brown spots or stains, narrowly fusoid ascospores, uniform width, narrow paraphyses (≤3 µm) and basal mycelium with abundant spheroid, pale brown, resinous exudates. Otidea bufonia and O. mirabilis are similar to O. filiformis in apothecia color and ascospore shape. Otidea bufonia differs from O. filiformis in having the longer ascospores (12–) 13–16.5 (–18) × 6–7.5 (–8) µm, and brown striate exudates on some hyphae of the medullary excipulum. O. mirabilis differs in dark-brown apothecia, purple to lilaceous-bluish shades on the receptacle surface and biflabellate crystal-like exudates in the medullary excipulum. The apothecia of Otidea korfii Pfister, F. Xu & Z.W. Ge, O. olivaceobrunnea and O. saliceticola Cartabia, M. Carbone & P. Alvarado also have brown tones. Otidea korfii is distinguished from O. filiformis by its ear-shaped apothecia, olivaceous brown receptacle surface, ellipsoid to broadly ellipsoid bigger ascospores (14.5–17 × 6.5–9 μm), resinous exudate of the ectal excipulum dissolving in MLZ and smooth basal mycelium. Otidea olivaceobrunnea differs from O. filiformis in olive-brown hymenium, ellipsoid ascospores. Otidea saliceticola differs in pale alutaceous-greyish hymenium surface, dark brown receptacle surface, wider ascospores (14–15 × 7.5–8 μm) with low Q value of 1.75–1.87.
Phylogenetic analyses revealed that the sequences of
O. filiformis were grouped into an independent clade with a strong support value (
Figure 1 and
Figure 2). DNA analysis showed that
O. filiformis shared less than 95.88% ITS sequence similarity with other
Otidea species. These supported the erection of the new species. One Finnish collection (MCVE 29372) identified as
O. bufonia by Carbone et al. [
22] is clustered into the
O. filiformis clade with strong support values in our phylogenetic trees (
Figure 1 and
Figure 2), and it shared more than 98% ITS similarity with our
O. filiformis and less than 94.6% with
O. bufonia. This evidence showed MCVE 29372 is more closely related to
O. filiformis, and whether it is conspecific with
O. filiformis need further morphological identification.
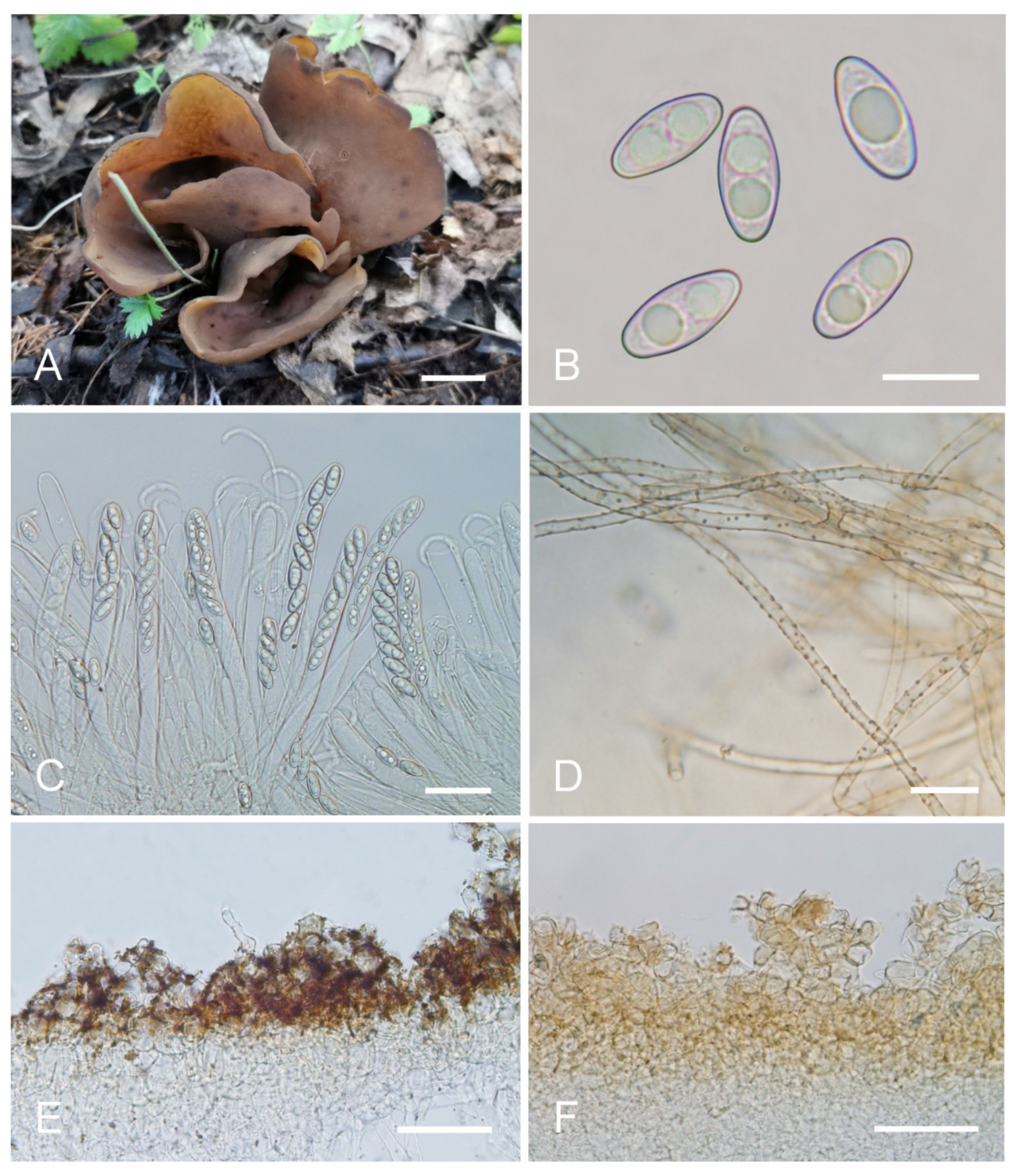
Otideakhakicolorata L. Fan & Y.Y. Xu, sp. nov. (
Figure 6).
MycoBank: MB843179.
Etymology: khakicolorata, referring to the khaki color of apothecia.
Holotype: China. Shanxi Province, Ningwu County, Guancen Mountain, Dashidong Forest Farm, alt. 2200m, among moss under coniferous forest dominated by Picea wilsonii Mast. and Larix principis-rupprechtii Mayr, 24 August 2017, X.Y. Yan YXY170824 (BJTC FM107).
Saprobic on soil. Apothecia solitary or gregarious in nature, 15–25 mm high, 5–8 mm wide, narrowly long ear-shaped, margin rounded, split, sessile or sub-stipitate. Hymenium surface khaki (#ca9a67) to pale ochre (#c08649) when fresh, yellowish ochre(#e3c57f) when dry, subsmooth. Receptacle surface concolorous with hymenium when fresh, slightly hygrophanous, dark reddish brown (#8c5738) when dry, furfuraceous. Warts absent or very low. Stipe, if present, very short. Basal tomentum and mycelium whitish to grayish whitish (#e7e2d9). Apothecial section 500–800 µm thick. Ectal excipulum of textura angularis, 70–120 µm thick, cells thin walled, brown, 10–25 × 6–20 µm. Medullary excipulum of textura intricata, 250–500 µm thick, hyphae 3.5–8 µm wide, sometimes slightly swollen, thin to slightly thick walled, septate, hyaline to light brown. Subhymenium ca. 60–90 µm thick, visible as a yellowish-brown zone, of densely arranged cylindrical to swollen cells. Paraphyses septate, curved to hooked, of uniform width at the apices, 2.5–3.7 µm wide, without notch. Asci 150–180 × 8.5–11 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores ellipsoid, sometimes slightly inequilateral, with two large guttules, smooth, hyaline, (8.5–) 9–10 (–10.5) × (4.5–) 5–6 (–6.5) µm (Lm × Wm= 9.5 × 5.5 µm, Q= 1.6–1.9, Qm= 1.75, n = 50). Receptacle surface with broadly conical warts, 30–40 µm high, formed by hyphoid hairs, of 2–5 subglobose to elongated cells, constricted at septa, 6–10 µm wide, sometimes with a gelatinous sheath. Resinous exudates abundant on the outer surface, yellow brown to dark brown, partly dissolving into amber drops in MLZ, turning reddish brown to dark reddish brown in KOH. Basal mycelium of 3–5.5 µm wide, septate, hyaline to pale-brown hyphae, unchanged in KOH, smooth or with little resinous exudates on the surface, partly dissolving in MLZ, and partly dissolving and more slowly in KOH.
Notes:
Otidea khakicolorata is characterized by khaki to pale-ochre, long, narrowly ear-shaped apothecia, small ascospores and resinous exudates on the ectal excipulum turning reddish brown in KOH.
Otidea nannfeldtii and
O. khakicolorata share similar apothecia shape and the reaction of the resinous exudate in the ectal excipulum and basal mycelium in MLZ and KOH, but
O. nannfeldtii can be distinguished by an ochre to orangish-ochre hymenium surface with pink tones, higher warts (45–85 µm) on the apothecial outer surface, medullary excipulum of textura intricata differentiated into two parts, and relatively bigger ascospores ((9–) 9.5–10.5 (–11.5) × 5.5–6.5 (–7) µm).
Otidea khakicolorata is phylogenetically close to
O. nannfeldtii; however, they are separated by a low support value (
Figure 1 and
Figure 2). DNA analyses showed that
O. khakicolorata shared less than 91% similarity in ITS sequence with
O. nannfeldtii.
Otidea parvula L. Fan & Y.Y. Xu, sp. nov. (
Figure 7).
MycoBank: MB843180.
Etymology: parvula, referring to the small apothecia of the fungus.
Holotype: China. Shanxi Province, Jiaocheng County, Guandi Mountain, Pangquangou National Nature Reserve, alt. 2000m, on soil in the mixed forest dominated by Picea wilsonii Mast., 7 September 2017, J.Z. Cao, Cao170803 (BJTC FM210-A).
Saprobic on soil. Apothecia gregarious to caespitose in nature, 8–15 mm high, 5–13 mm wide, initially narrowly to broadly ear-shaped, margin rounded, then expanding and sometimes becoming irregularly ear-shaped or cup-shaped, split, stipitate, or sessile. Hymenium surface pale whitish ochre (#ffffed) to ochre yellow (#b39a7f) when fresh, pale ochre brown (#c2a461) when dry, subsmooth. Receptacle surface pale ochre (#d7c498) to orangish ochre (#dba35e) when fresh, hygrophanous, light yellowish brown (#d7c498) when dry, furfuraceous. Stipe 3–7 × 3–5 mm. Basal tomentum and mycelium white. Apothecial section 500–700 µm thick. Ectal excipulum of textura angularis, 70–120 µm thick, cells thin walled, pale brown, 9–24 × 7–20 µm. Medullary excipulum of textura intricata, 80–160 µm thick, hyphae 3–7 µm wide, sometimes slightly swollen, thin to thick walled, septate, hyaline to light brown. Subhymenium c. 50–80 µm thick, visible as a brown zone, of densely arranged cylindrical to swollen cells, with scattered brown resinous exudate at septa. Paraphyses septate, bent to curved, sometimes straight, of uniform width or slightly enlarged at the apices, 2.5–4.5 µm wide, sometimes with 1–2 notches near the apex. Asci 150–200 × 10.5–13.5 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores ellipsoid, slightly inequilateral, with two large guttules, sometimes with only one big guttule, smooth, hyaline, (12.5–) 13–15.5 (–16) × (6.5–) 6.8–8 (–8.6) µm (Lm × Wm = 14.3 × 7.5 µm, Q = 1.7–2.1, Qm = 1.9, n = 50). Receptacle surface with low warts, 30–50 µm high, formed by short, fasciculate, hyphoid hairs, of 5–6 subglobose to elongated cells, constricted at septa, 5–10 µm wide. Resinous exudates absent. Basal mycelium of 2.5–5 µm wide, septate, hyaline to pale brown hyphae, smooth, unchanged in KOH, turning yellow in MLZ.
Other materials examined: China. Shanxi Province, Jiaocheng County, Guandi Mountain, Pangquangou National Nature Reserve, alt. 2000m, on soil in the mixed forest dominated by Picea wilsonii, 7 September 2017, J.Z. Cao, Cao170803 (BJTC FM210-B).
Notes:
Otidea parvula is easily recognized by the stipitate, irregularly ear-shaped or cup-shaped, small, pale-whitish-ochre, ochre-yellow to orangish-ochre apothecia, straight to bent paraphyses and furfuraceous receptacle surface.
Otidea parvula and
O. adorniae Agnello, M. Carbone & P. Alvarado are somewhat similar in the color of the apothecia, but
O. adorniae differs in its larger apothecia and smaller ascospores (11.8 × 6.4 µm).
Otidea parvula and
O. parvispora (Parslow & Spooner) M. Carbone, Agnello, Kautmanová, Z.W. Ge & P. Alvarado have the highest ITS sequence similarity of 96%, but upon examination of the phylogenetic tree (
Figure 1 and
Figure 2), they don’t seem to be closely related, and
O. parvispora is easily distinguished by its pale hymenium and smaller ascospores ((11.0–) 11.5–13.0 × 5.0–6.5 µm).
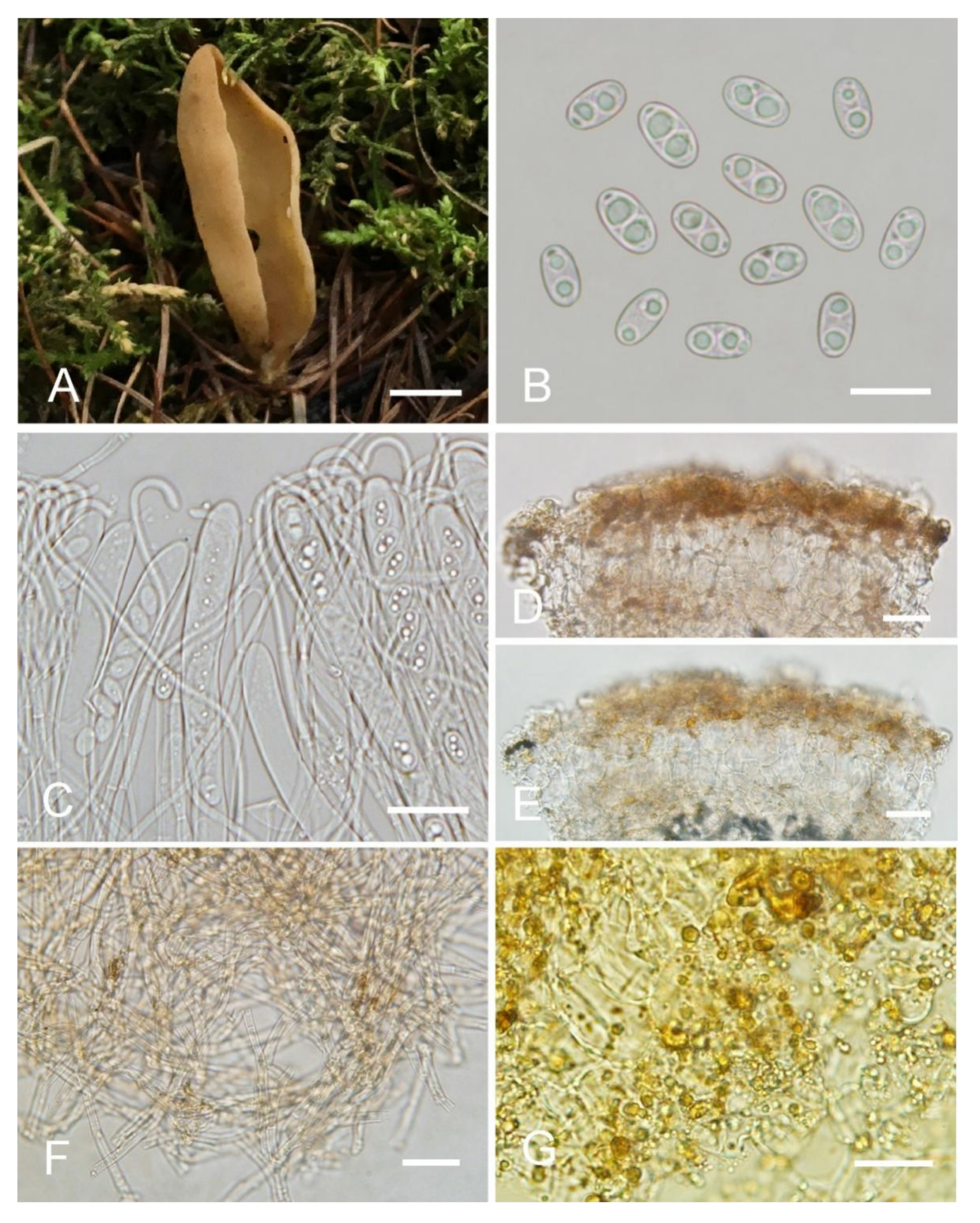
Otidea plicara L. Fan & Y.Y. Xu, sp. nov. (
Figure 8).
MycoBank: MB843181.
Etymology: plicara, referring to the small fold on the apothecia.
Holotype: China. Shanxi Province, Jiaocheng County, Guandi Mountain, Badaogou valley, alt. 2000m, on soil in the mixed forest dominated by Picea wilsonii, 7 September 2017, J.Z. Cao, Cao170855 (BJTC FM262-A).
Saprobic on soil. Apothecia solitary or gregarious in nature, 15–28 mm high, 12–42 mm wide, initially spoon-shaped or broadly ear-shaped, soon expending, in the end almost deeply cup-shaped, often broader above, margin entire, with a small fold on the apothecia, seemingly spilt yet not split, stipitate. Hymenium surface pale greyish brown (#6f6a61) to dark brown (#665856), margin yellow ochre (#837050) when fresh, when dry becoming slightly lighter but dull, light brown (#ab9876), subsmooth. Receptacle surface dark reddish brown (#7c6052) when fresh, slightly hygrophanous, pale ochre brown (#a48e6a) when dry, finely furfuraceous, wrinkle veined at the base. Stipe 8–15 × 5–8 mm. Basal tomentum and mycelium abundant, white to pale cream (#f1f9ed). Apothecial section 650–1000 µm thick. Ectal excipulum of textura angularis, 60–100 µm thick, cells thin walled, brown, 11–34 × 9–28 µm. Medullary excipulum of textura intricata, 300–500 µm thick, hyphae 3–7.5 µm wide, sometimes slightly swollen, thin walled, septate, hyaline to light brown. Subhymenium ca. 80–130 µm thick, visible as a yellowish-brown zone. Paraphyses septate, curved to hooked, usually enlarged at the apices, 3.5–5.5 µm wide at apex, 2–3 µm below. Asci 160–220 × 10–14 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores ellipsoid, sometimes slightly inequilateral, with one to two large guttules, smooth, hyaline, (12.5–) 13.5–16 (–17) × (6–) 6.5–8 (–8.5) µm (Lm × Wm = 14.7 × 7.4 µm, Q = 1.8–2.2, Qm = 2, n = 50). Receptacle surface with hyphoid hairs, 50–80 µm long, of 3–6 ovoid or subglobose to elongated cells, constricted at septa, 4–9 µm wide. Resinous exudates absent. Basal mycelium of interwoven, 2.5–6 µm wide, septate, hyaline to pale brown hyphae, unchanged in KOH, smooth, turning yellow in MLZ.
Other materials examined: China. Shanxi Province, Jiaocheng County, Guandi Mountain, Badaogou Scenic Area, alt. 2000m, on soil in the mixed forest dominated by Picea wilsonii Mast., 7 September 2017, J.Z. Cao, Cao170855 (BJTC FM262-B).
Notes:
Otidea plicara is characterized by greyish-brown to dark-brown, stipitate, rarely split, deeply cup-shaped apothecia, small ascospores, enlarged paraphyses and the lack of resinous exudates on the ectal excipulum and basal mycelium. Macroscopically,
Otidea apophysata (Cooke & W. Phillips) Sacc. and
O. platyspora Nannf. have similar apothecial shape and color to
O. plicara, but
O. apophysata can be distinguished by the larger ascospores (20–24.5 × 9–11 µm) and frequently branched paraphyses.
Otidea platyspora can be distinguished by split apothecia and larger ascospores (18–22 × (9.5–)10.5–12 µm). DNA analyses showed that
O.
plicara shared less than 92% similarity in ITS sequence with other species of
Otidea. Phylogenetic analyses revealed that the sequences of
O. plicara were grouped into an independent clade with a strong support value (
Figure 1 and
Figure 2). These supported the erection of the new species.
Otideapurpureobrunnea L. Fan & Y.Y. Xu, sp. nov. (
Figure 9).
MycoBank: MB843182.
Etymology: purpureobrunnea, referring to the purple-brown tone of apothecia.
Holotype: China. Shanxi Province, Qinshui County, Tuwo Township, Shangwoquan Village, alt. 1200m, on soil under Quercus sp., 25 August 2020, H. Liu 1065 (BJTC FM1048).
Saprobic on soil. Apothecia gregarious to caespitose in nature, 25–55 mm high, 50–80 mm wide, initially ear shaped, soon expanding, becoming broadly ear shaped or deeply cup shaped, often elongated on one side, split, margin sometimes lobate, stipitate or sessile. Hymenium surface ochraceous brown (#804618), grayish purple (#5e4f5f) to purple brown (#39242f) when fresh, gray brown to dark brown (#3e2c1c) when dry, subsmooth. Receptacle surface grayish purple brown (#816e71) to dark purple brown (#483131) when fresh, sometimes partly dark yellow brown, slightly hygrophanous, some apothecia with shallowly wrinkled, dark brown (#492615) when dry, furfuraceous to finely warty. Stipe 5–10 × 4–8 mm. Basal tomentum and mycelium whitish to pale brown (#dccdbf). Apothecial section 900–1300 µm thick. Ectal excipulum of textura angularis, 80–120 µm thick, cells thin walled, brownish, 13–33 × 7–26 µm. Medullary excipulum of textura intricata, 500–900 µm thick, formed of loosely woven cylindrical to slightly swollen thin-walled hyphae, 4.5–11 µm wide, septate, hyaline to light brown, with brown resinous exudates at septa. Subhymenium c. 100–150 µm thick, visible as a brown zone, of densely arranged cylindrical to swollen cells, with scattered brown resinous exudate at septa. Paraphyses septate, curved to hooked, a few curved, sometimes forming a coil or helix, of the same width or often enlarged at the apices, 3.5–5 µm wide, 2–3.3 µm below, sometimes with 1–2 notches, or with an obvious bulge near the apex. Asci 140–190 × 8.5–15 µm, 8-spored, unitunicate, cylindrical, hyaline, long pedicellate, arising from croziers, non-amyloid, ascospores released from an eccentric split at the apical apex. Ascospores ellipsoid to slightly subfusoid, inequilateral, with two large guttules, sometimes with only one big guttule, smooth, hyaline, (12.5–) 13–15 (–15.5) × (6–) 6.5–7 (–7.5) µm (Lm × Wm= 14 × 6.5 µm, Q= 1.9–2.3, Qm= 2.1, n = 50). Receptacle surface with broad conical warts, 35–60 µm high, formed by short, fasciculate, hyphoid hairs, of 2–5 subglobose to elongated cells, constricted at septa, 6–13 µm wide. Resinous exudates abundant on the outer surface, yellow brown to dark brown, partly dissolving into particles in MLZ, entirely dissolving and turning yellow in KOH. Basal mycelium of 3.5–6 µm wide, septate, hyaline to pale brown hyphae, turning yellow in KOH, mostly smooth, a few with very small, spheroid, pale-brown, resinous exudates, dissolving in KOH, partially dissolving in MLZ.
Other materials examined: China. Shanxi Province, Qinshui County, Tuwo Township, Shangwoquan Village, alt. 1200m, on soil under Quercus sp., 25 August 2020, H. Liu 1079 (BJTC FM1061).
Notes: Otidea purpureobrunnea is characterized by the stipitate, broadly ear-shaped to cup-shaped, grayish-purple to purple-brown apothecia, ellipsoid to slightly subfusoid ascospores, paraphyses enlarged at the apices, with 1–2 notches or an obvious bulge and smooth basal mycelium. Similar to O. purpureobrunnea, the apothecia of O. bufonia, O. cupulata, O. mirabilis, O. purpurea, O. purpureogrisea, O. smithii, and O. subpurpurea all have some purple tones, but Otidea bufonia differs in its fusoid ascospores, the presence of hyphae with striate resinous exudates in the medullary excipulum, resinous exudates of the ectal excipulum not turning bright yellow in KOH, and in having abundant resinous exudates on the basal mycelium. Otidea mirabilis differs by having fusoid ascospores, resinous exudates of the ectal excipulum that do not turn bright yellow in KOH and when present, biflabellate, crystal-like exudates in the medullary excipulum. Otidea purpurea and O. subpurpurea are easily distinguished by the obviously smaller spores (O. subpurpurea: 9–12 × 4.5–6 µm; O. purpurea: 8–10 × 4.5–6 µm). Otidea purpureogrisea is distinguished by the purple-gray tone of the receptacle surface near the base and resinous exudates of the ectal excipulum turning amber in MLZ and turning brown in KOH. Otidea smithii is distinguished by typically narrower, ear-shaped apothecia, relatively shorter ascospores (12–14 × 6–7.5 µm) with a lower Qm value (1.9–2), and resinous exudates of the ectal excipulum not turning bright yellow in KOH. For a comparison with O. cupulata see under that species below.
Phylogenetic analyses revealed that
O. purpureobrunnea and
O. filiformis are grouped together with a low support value (
Figure 2), but
O. filiformis is easy to distinguish from
O. purpureobrunnea by its apothecia without purple tones, fusoid ascospores, same width and narrow paraphyses (≤3 µm), as well as its basal mycelium with abundant spheroid, pale brown, resinous exudates. DNA analysis showed that
O. purpureobrunnea shared less than 94.53% similarity in its ITS sequence with
O. filiformis. These indicate that they are two different species.
Otidea subpurpurea W.Y. Zhuang, Mycologia Montenegrina 10: 238 (2007).
Holotype: China, Yunnan Province, Kunming City, Kunming Institute of Botany, alt. 1980m, 8 October 2005, Z.L. Yang 4602, (HKAS 49443); Isotype (HMAS 97530).
= Otidea bicolor W.Y. Zhuang & Zhu L. Yang, Mycotaxon 112: 35 (2010).
Holotype: China, Yunnan Province, Kunming City, Heilongtan Park, 16 August 2008, Z.L. Yang 5156, (HKAS 54453); Isotype (HMAS 188415).
= Otidea pruinosa Ekanayaka, Q. Zhao & K.D. Hyde, Fungal Diversity 87: 130 (2017).
Holotype: China, Yunnan Province, Kunming City, Xishan Scenic Area, 15 September 2012, T. Guo 617, (HKAS 81819).
Materials examined: China, Yunnan Province, Kunming City, Kunming Institute of Botany, alt. 1980m, 9 October 2005, Z.L. Yang 4602, (HKAS 49443); Isotype (HMAS 97530). China, Yunnan Province, Kunming City, Heilongtan Park, 16 August 2008, Z.L. Yang 5156, (HKAS 54453); Isotype (HMAS 188415). ibid., (HKAS 54449). China, Yunnan Province, Kunming City, Xishan Scenic Area, 15 September 2012, T. Guo 617, (HKAS 81819).
Notes:
Otidea bicolor,
O. pruinosa and
O. subpurpurea are highly similar species. In fact, previous scholars have also noticed the phenomenon that the type sequences of the three species are clustered together [
22,
25]; however, due to the unavailability of specimens, this issue has not been formally addressed. In this study, we examined the type specimens and obtained multiple loci sequences from them. DNA analyses revealed that
O. pruinosa,
O. bicolor, and
O. subpurpurea share high sequence similarity (ITS: >98.87%; nrLSU: >99.53%;
tef1-α: >99.72%;
rpb2: >99.45%). We performed morphological observation on these type specimens and found that there was no obvious difference in microscopic features. The reaction of the resinous exudate in the ectal excipulum and basal mycelium in MLZ and KOH are also the same. Although the receptacle surface of
O. bicolor and
O. subpurpurea is purplish in tint when fresh, the receptacle surface of
O. pruinosa is without a purplish tint [
23,
28,
29], but that may be influenced by its habitat.
Otidea pruinosa is proposed as a new species because of receptacle surface with pruinose, but we found a similar granulate on the surface of dry specimens of
O. bicolor and
O. subpurpurea. In addition, phylogenetic analyses based on the two-gene and four-gene datasets also confirmed that they represent the same species, so here we formally treat
O. bicolor and
O. pruinosa as synonyms of
O. subpurpurea. The sequence from ZMU124 (label as
O. bufonia) from Guizhou province of China grouped with
O. subpurpurea with a high support value (
Figure 1), indicating that
O. subpurpurea seems widely distributed in southwest China. Similarly, the sequence from JS150904-08 from Korea named
O. bufonia [
45] was also grouped into this clade (
Figure 1). We checked the original morphological description by Jin et al. [
45] and found that its ascospores size does not conform to
O. bufonia, but instead to
O. subpurpurea. This indicates that
O. subpurpurea also occurs in Korea.
Otidea mirabilis Bolognini & Jamoni in Jamoni, Funghi e Ambiente 85–86: 56 (2001). (
Figure 10).
Habitat: on soil under mixed forest of Larix principis-rupprechtii and Betula sp.
Distribution: Known from the northeast, northern, northwest and southwest regions of China.
Materials examined: China, Yunnan Province, Jingdong County, Ailao Mountain, Xujiaba Village, alt. 2500m, 24 August 1994, M. Zang, 12389 (HKAS 28129). China, Gansu Province, Wudu County, Liangshui Town, Gongba River Beach, alt. 2600m, 11 July 1996, M.S. Yuan, 2213 (HKAS 30708). China, Sichuan Province, Hongyuan County, Kangle Town, alt. 3400m, 19 August 1998, M.S. Yuan, 3433 (HKAS 33633). China, Jilin Province, Fusong County, Songjiang River, 19 August 2000, M.S. Yuan, 4725 (HKAS 37272). China, Xinjiang Autonomous Region, Jimusa’er, alt. 1700m, 1 August 2003, W.Y. Zhuang & Y. Nong, 4657 (HMAS 83568). China, Inner Mongolia Autonomous Region, Chifeng City, Baiyin Aobao National Nature Reserve, 2 August 2013, Tolgor Bau (HMJAU 26926). China, Shanxi Province, Jiaocheng County, Guandi Mountain, Shanshui Village, alt. 1800m, 8 September 2017, J.Z. Cao, CAO170863 (BJTC FM292). China, Shanxi Province, Jiaocheng County, Pangquangou Nature Reserve, alt. 2100m, 28 August 2018, H. Liu, LH234 (HSA 234).
Notes: The occurrence of
O. mirabilis is confirmed in China based on morphological and DNA evidence in this study. Olariaga et al. [
4] showed already that
O. mirabilis occur in China using nrLSU sequences from two Chinese collections in GenBank (identified as
O. leporina by Zhuang [
17] and Liu and Zhuang [
18], but by morphology it has not been previously confirmed, as Olariaga et al. did not study those two collections morphologically. It is interesting that two distinct clades were revealed, one comprising Chinese specimens, and another comprising specimens from Europe. The Chinese specimens shared 98.46–99.84% ITS sequence similarity and the European ones had 99.54–99.85% similarity, while the similarities between the two proveniences were 97–98.5%. However, we found no significant morphological differences between the Chinese and European specimens, which probably resulted from the geographic distance.
Otidea nannfeldtii Harmaja, Karstenia 15: 31 (1976). (
Figure 10).
Habitat: on soil under mixed forest of Larix principis-rupprechtii.
Distribution: Known in northern China and northwest China.
Materials examined: China, Shanxi Province, Ningwu County, Guancen Mountain, Qiuqiangou Village, alt. 2100m, on soil under L. principis-rupprechtii Mayr, 25 August 2017, X.Y. Yan, YXY170836 (BJTC FM168); ibid., X.Y. Yan, YXY170837 (BJTC FM169); ibid., X.Y. Yan, YXY170838 (BJTC FM170). China, Shanxi Province, Jiaocheng County, Guandi Mountain, Pangquangou Nature Reserve, alt. 2000m, 6 September 2017, J.Z. Cao, CAO170829 (BJTC FM236); ibid., J.Z. Cao, CAO170836 (BJTC FM243). China, Xinjiang Autonomous Region, Jimusa’er, alt. 1700m, 1 August 2003, W.Y. Zhuang & Y. Nong, 4655 (HMAS 83573).
Notes: The occurrence of
O. nannfeldtii in China is first confirmed based on molecular and morphological evidence.
Otidea nannfeldtii is originally described in Europe, and also reported from North America [
4]. Before this study, there are no DNA data that support the existence of this species in China.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












