
Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Isolates and Inoculation
2.3. Evaluation of Symptoms and Risks
2.4. Preparing Samples for Toxin Analyses
2.5. Toxin Analysis
2.6. Statistical Methods
3. Results
3.1. Experiment 1, 2017/2018
3.1.1. Ear Rot Severity
3.1.2. Resistance to Toxins
3.2. Experiment 2, 2019–2020
3.2.1. Ear Rot Data
3.2.2. Resistance to Toxin Contamination
4. Discussion
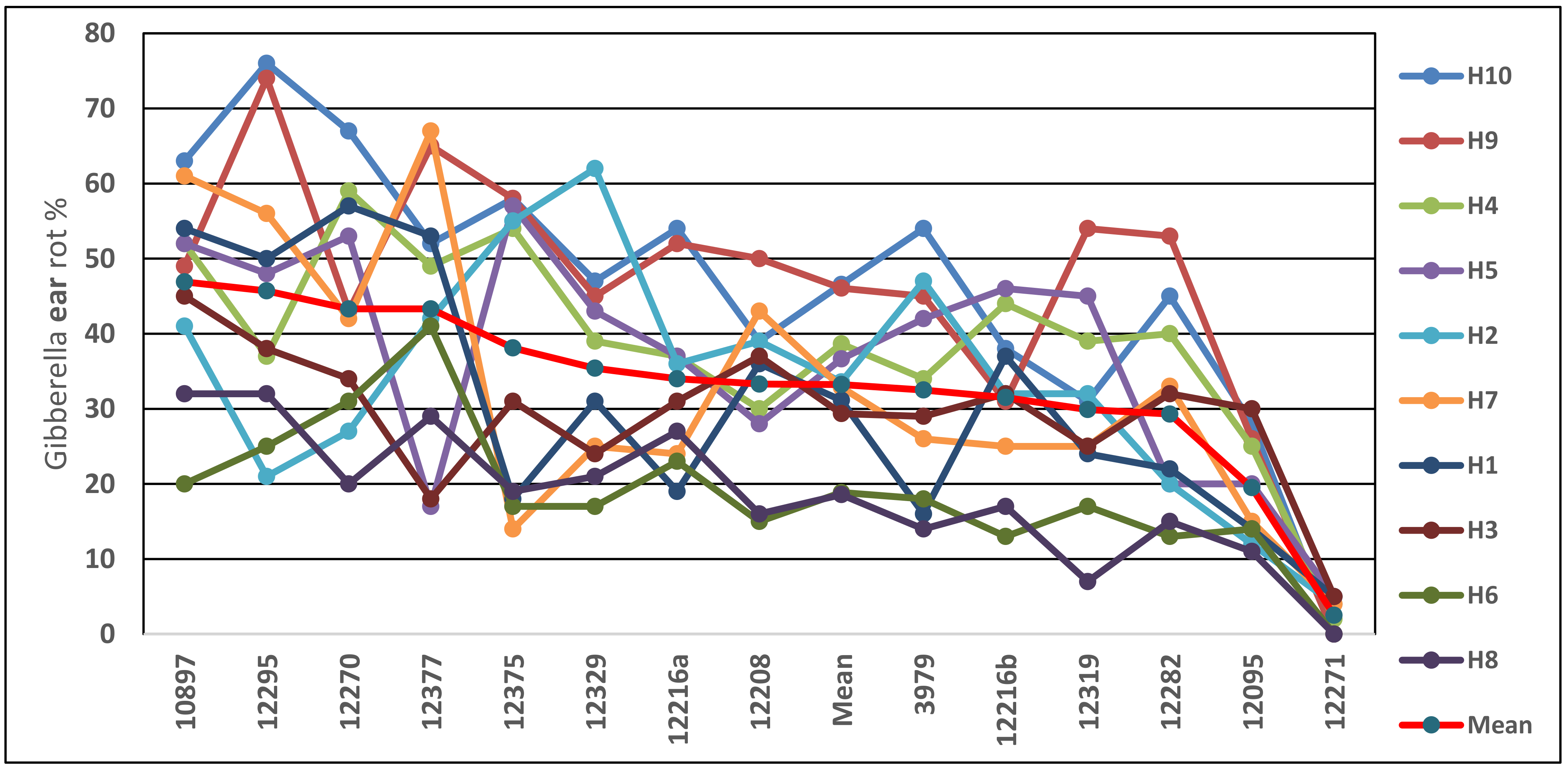
4.1. Evaluating Hybrids under Artificial and Natural Infection Regimes
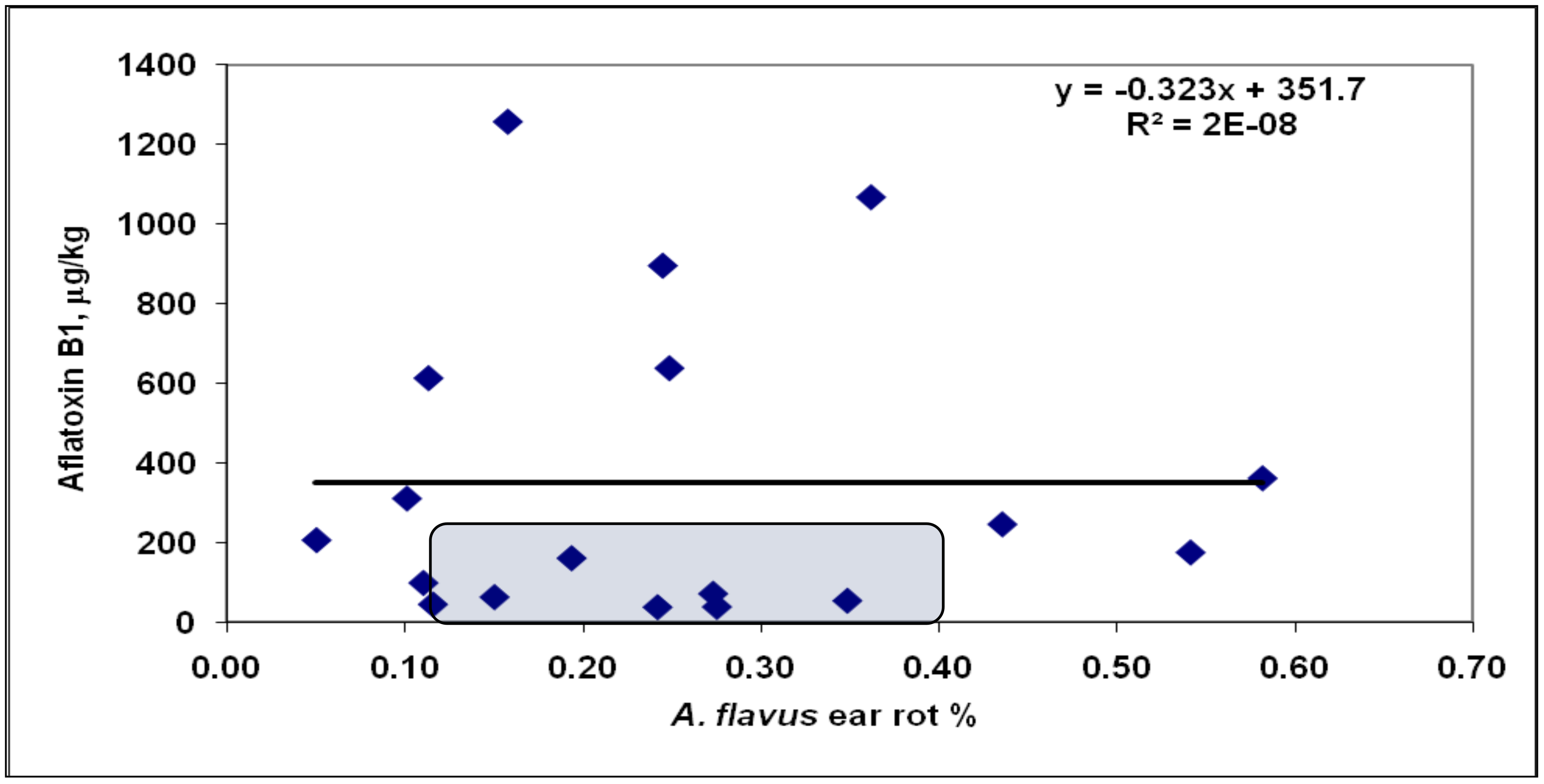
4.2. Visual Ear Rot, Rachilla-Mediated Infection, and Toxin Contamination
4.3. Advantages of the Suggested Testing Methodology
4.4. Food or Feed Safety Risk and the Toxin Production for One Percent of the Ear Infections Analyzed
- Separate artificial inoculation ear rot data for the three pathogens.
- Separate artificial toxin data for the three pathogens.
- Natural infection data for Fusarium spp. and Aspergillus spp.
- Natural toxin data for the three most important toxins.
4.5. Resistance to Ear Rots, Toxin Contamination, and the Changing Climate
4.6. Breeding Aspects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AER | Aspergillus ear rot |
| Af | Aspergillus flavus |
| AFB1 | aflatoxin B1 |
| ANOVA | analysis of variance |
| Bt | hybrids containing an insecticide gene from B. thuringiensis |
| DKC | DeKalb Company: belonging to Bayer Inc., Germany |
| DON | deoxynivalenol |
| FAO | Food Agricultural Organization of the United Nations |
| FER | Fusarium ear rot (mostly F. verticillioides) |
| Fg | F. graminearum |
| FB1+B2 | fumonisin B1+B2 |
| Fv | F. verticillioides |
| GER | Gibberella ear rot (mostly F. graminearum) |
| KR | Kernel resistance |
| LSD | 5% limit of significant difference |
| MMT | million metric tons |
| MS/MS | mass spectrometry |
| PCR | polymerase chain reaction |
| RAGT | French Seed Company |
| SR | silk channel resistance |
| UPLC | ultra-high-pressure liquid chromatography |
References
- International Grains Council. International Grains Council Grain Market Report Five-Year Baseline Projections of Supply and Demand for Wheat, Maize (Corn), Rice and Soyabeans to 2023/24 March 2019; International Grains Council: London, UK, 2019; pp. 1–4. Available online: http://www.igc.int/en/downloads/gmrsummary/gmrsumme.pdf (accessed on 15 January 2020).
- Lanubile, A.; Machietto, V.; Marocco, A. Breeding maize for resistance to mycotoxins. In Mycotoxin Reduction in Grain Chains; Leslie, J.F., Logrieco, A.F., Eds.; Wiley Blackwell: Ames, IA, USA; Chichester, UK, 2014; pp. 37–58. ISBN 978-0-8138-2083-5. [Google Scholar]
- Miedaner, T.; Loeffler, M.; Bolduan, C.; Kessel, B.; Ouzunova, M.; Mirdita, V.; Melchinger, A.E. Genetic variation for resistance and mycotoxin content of European maize inoculated with Fusarium graminearum and F. verticillioides. Cereal Res. Commun. 2008, 36 (Suppl. B), 45–48. [Google Scholar] [CrossRef]
- Mesterházy, A.; Lemmens, M.; Reid, L.M. Breeding for resistance to ear rots caused by Fusarium spp. in maize—A review. Plant Breed. 2012, 131, 1–19. [Google Scholar] [CrossRef]
- Szabo, B.; Toth, B.; Toth Toldine, E.; Varga, M.; Kovacs, N.; Varga, J.; Kocsube, S.; Palagyi, A.; Bagi, F.; Budakov, D.; et al. A new concept to secure food safety standards against Fusarium species and Aspergillus flavus and their toxins in maize. Toxins 2018, 10, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corneillie, S. DEKALB 2021 Seed Guide Easter Canada, Bayer. Available online: https://www.cropscience.bayer.ca/grower-tools/resources-and-guides/dekalb-seed-guide (accessed on 28 January 2022).
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the US corn industry from AFB1 toxin contamination. Food Addit. Contam. Part A 2016, 33, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Oláh, J.; Popp, J. Losses in the Grain Supply Chain: Causes and Solutions. Sustainability 2020, 12, 2342. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Munkvold, G.P.; White, D.G. (Eds.) Compendium of Corn Diseases; The American Phytopathological Society (APS) Press: St. Paul, MN, USA, 2016; 165p. [Google Scholar]
- Mesterházy, Á.; Vojtovics, M. A kukorica Fusarium spp. okozta fertőzöttségének vizsgálata 1972–1975-ben. (Infection of maize grain by Fusarium spp. 1972–1977. Növénytermelés 1977, 26, 367–378. [Google Scholar]
- Mesterházy, Á.; Vojtovics, M. Kukorica magminták gombaflórája Magyarországon 1974–1975-ben. (Fungal flora of seed samples of corn in Hungary). Növényvédelem (Plant Prot.) 1977, 13, 441–446. [Google Scholar]
- Rose, L.J.; Okoth, S.; Beukes, I.; Ouko, A.; Mouton, M.; Flett, B.C.; Makumbi, D.; Viljoen, A. Determining resistance to Fusarium verticillioides and fumonisin accumulation in African maize inbred lines resistant to Aspergillus flavus and aflatoxins. Euphytica 2017, 213, 93. [Google Scholar] [CrossRef]
- Palumbo, R.; Gonçalves, A.; Gkrillas, A.; Logrieco, A.; Dorne, J.L.; Dall’Asta, C.; Battilani, P. Mycotoxins in maize: Mitigation actions, with a chain management approach. Phytopath. Mediterr. 2020, 59, 5–28. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Tóth, B.; Szieberth, D. Toxin Termelő Gombák Okozta Növénybetegségek Búzában és Kukoricában. (Toxic Diseases of Maize and Wheat), 2nd ed.; Magy Kukorica Klub/Hung Maize Club: Kőszárhegy, Hungary, 2019; p. 72. [Google Scholar]
- Parkanyi, G. Raktározási kihívások (Challanges in grain storage). Agronaplo 2020, 24, 44. [Google Scholar]
- Buric, D.; Doderovic, M. Projected temperature changes in Kolašin (Montenegro) up to 2100 according to EBU-POM and ALADIN regional climate models. Q. J. Hung. Meteorol. Serv. 2020, 124, 427–445. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; van der Fels-Klerx, H.J.; Moretti, A.; Leggieri, M.C.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Maggiore, A.; Afonso, A.; Barrucci, F.; De Sanctis, G.; European Food Safety Authority (EFSA). Climate Change as a Driver of Emerging Risks for Food and Feed Safety, Plant, Animal Health and Nutritional Quality; EFSA Supporting Publication: Brussels, Belgium, 2020; 146p. [CrossRef]
- Battilani, P.; Leggieri, M.C.; Rossi, V.; Giorni, P. AFLA-maize, a mechanistic model for Aspergillus flavus infection and aflatoxin B1 contamination in maize. Comput. Electron. Agric. 2013, 94, 38–46. [Google Scholar] [CrossRef]
- Okoth, S.; Nyongesa, B.; Ayugi, V.; Kangethe, E.; Korhonen, H.; Joutsjoki, V. Toxigenic potential of Aspergillus species occurring on maize kernels from two agro-ecological Zones in Kenya. Toxins 2012, 4, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.K.; Sinha, K.K. Competition between a toxigenic Aspergillus flavus strain and other fungi on stored maize kernels. J. Stored Prod. Res. 1993, 29, 75–80. [Google Scholar] [CrossRef]
- Degola, F.; Berni, E.; Spotti, E.; Ferrero, I.; Restivo, F.M. Facing the problem of “false positives”: Re-assessment and improvement of a multiplex RT-PCR procedure for the diagnosis of A. flavus mycotoxin producers. Int. J. Food Microbiol. 2009, 129, 300–305. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Toldine Toth, E.; Szel, S.; Varga, M.; Toth, B. Resistance of maize hybrids to Fusarium graminearum, F. culmorum, and F. verticillioides ear rots with toothpick and silk channel inoculation, as well as their toxin production. Agronomy 2020, 10, 1283. [Google Scholar] [CrossRef]
- Foley, D.C. Systemic infection of corn by Fusarium moniliforme. Phytopathology 1962, 52, 870–872. [Google Scholar]
- Pechanova, O.; Pechan, T.; Williams, W.P.; Luthe, D.S. Proteomic analysis of maize rachis: Potential roles constitutive and induced proteins in resistance to Aspergillus flavus infection and aflatoxin accumulation. Protheomics 2011, 11, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Smart, M.G.; Wicklow, D.T.; Caldwell, R.W. Pathogenesis of Aspergillus ear rot of maize: Light microscopy of fungal spread from wounds. Phytopathology 1990, 80, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Mu, C.; Gao, J.; Zhou, Z.; Wang, Z.; Sun, X.; Zhang, X.; Wu, J. Genetic analysis of cob resistance to F. verticillioides: Another step towards the protection of maize from ear rot. Theor. Appl. Genet. 2019, 132, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, E.; Ellner, F. Distribution of disease symptoms and mycotoxins in maize ears infected by Fusarium culmorum and Fusarium graminearum. Mycotoxin Res. 2015, 31, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Kovács, G., Jr.; Kovács, K. Breeding resistance for Fusarium ear rot (FER) in corn. 18th Int. Conference on Maize and Sorghum Genetics and Breeding, Eucarpia, Beograd. Acta Biol. Yugosl. Ser. F Genet. 2000, 32, 495–505. [Google Scholar]
- Reid, L.M.; Hamilton, R.E.; Mather, D.E. Screening Maize for Resistance to Gibberella Ear Rot; Publication 1996–5E; Agriculture and Agri-Food Canada, Technical Bulletin: Ottawa, ON, Canada, 1996; 62p. [Google Scholar]
- Chungu, C.; Mather, D.E.; Reid, L.M.; Hamilton, R.I. Comparison of techniques for inoculating maize silk, kernel, and cob tissues with Fusarium graminearum. Plant Dis. 1996, 80, 81–84. [Google Scholar] [CrossRef]
- Reid, L.M.; McDiarmid, G.; Parker, A.J.; Woldemariam, T. CO441 corn inbred line. Can. J. Plant Sci. 2003, 83, 79–80. [Google Scholar] [CrossRef]
- Loeffer, M.; Miedaner, T.; Kessel, B.; Ouzunova, M. Mycotoxin accumulation and corresponding ear rot rating in three maturity groups of European maize inoculated by two Fusarium species. Euphytica 2010, 174, 153–164. [Google Scholar]
- Munkvold, G.P.; Desjardins, A.E. Fumonisins in Maize. Can we reduce their occurrence? Plant Dis. 1997, 81, 556–565. [Google Scholar]
- Miller, S.S.; Reid, L.M.; Harris, L.J. Colonization of maize silks by Fusarium graminearum, the causative organism of Gibberella ear rot. Can. J. Bot. 2007, 85, 369–376. [Google Scholar] [CrossRef]
- Presello, D.A.; Reid, L.M.; Mather, D.E. Resistance of Argentine maize germplasm to Gibberella and Fusarium ear rots. Maydica 2004, 49, 73–81. [Google Scholar]
- Presello, D.A.; Iglesias, J.; Botta, G.; Reid, L.M.; Lori, G.A.; Eyherabide, G.H. Stability of maize resistance to the ear rots caused by Fusarium graminearum and F. verticillioides in Argentinean and Canadian environments. Euphytica 2006, 147, 403–407. [Google Scholar] [CrossRef]
- Loeffler, M.; Kessel, B.; Ouzunova, M.; Miedaner, T. Population parameters for resistance to Fusarium graminearum and Fusarium verticillioides ear rot among large sets of early, mid-late and late maturing European maize (Zea mays L.) inbred lines. Theor. Appl. Genet. 2010, 120, 1053–1062. [Google Scholar] [CrossRef]
- Henry, W.B.; Williams, W.P.; Windham, G.L.; Hawkins, L.K. Evaluation of maize inbred lines for resistance to Aspergillus and Fusarium ear rot and mycotoxin accumulation. Agron. J. 2009, 101, 1219–1226. [Google Scholar] [CrossRef] [Green Version]
- Reid, L.M.; Zhu, A.X.; Parker, A.A.; Yan, A.W. Increased resistance to Ustilago zeae and Fusarium verticillioides in maize inbred lines bred for Fusarium graminearum resistance. Euphytica 2009, 165, 567–578. [Google Scholar] [CrossRef]
- Robertson-Hoyt, L.A.; Betran, J.; Payne, G.A.; Isakeit, T.; White, D.G.; Maragos, C.M.; Molnar, T.L.; Holland, J.B. Relationships among resistances to Fusarium and Aspergillus ear rots and contamination by fumonisin and aflatoxin in maize. Phytopathology 2007, 97, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, W.P. Breeding for resistance to aflatoxin accumulation in maize. Mycotoxin Res. 2006, 22, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Williams, W.P.; Windham, G.L. Diallel analysis of fumonisin accumulation in maize. Field Crops Res. 2009, 114, 324–326. [Google Scholar] [CrossRef]
- Reid, L.M.; Mather, D.E.; Hamilton, R.I. Distribution of deoxynivalenol in Fusarium graminearum infected maize ears. Phytopathology 1996, 86, 110–114. [Google Scholar] [CrossRef]
- Pascale, M.; Visconti, A.; Pronczuk, M.; Wisniewska, H.; Chelkowski, J. Accumulation of fumonisins in maize hybrids inoculated under field conditions with Fusarium moniliforme Sheldon. J. Sci. Food Agric. 1997, 74, 1–6. [Google Scholar] [CrossRef]
- Perkowski, J.; Pronczuk, M.; Chelkowski, J. Deoxynivalenol and acetyldeoxynivalenol accumulation in field maize inoculated by F. graminearum. J. Phytopathol. 1997, 145, 113–116. [Google Scholar] [CrossRef]
- Reid, L.M.; Sinha, R.C. Maize maturity, and the development of Gibberella ear rot symptoms and deoxynivalenol after inoculation. Eur. J. Plant Pathol. 1998, 104, 147–154. [Google Scholar] [CrossRef]
- Bolduan, C.; Miedaner, T.; Schipprack, W.; Dhillon, B.S.; Melchinger, A.E. Genetic Variation for Resistance to Ear Rots and Mycotoxins Contamination in Early European Maize Inbred Lines. Crop Sci. 2009, 49, 2019–2028. [Google Scholar] [CrossRef]
- Focker, M.; van der Fels-Klerx, H.J.; Magan, N.; Edwards, S.G.; Grahovac, M.; Bagi, F.; Budakov, D.; Suman, M.; Schatzmayr, G.; Krska, R.; et al. The impact of management practices to prevent and control mycotoxins in the European food supply chain: MyToolBox project results. World Mycotoxin J. 2020, 14, 139–154. [Google Scholar] [CrossRef]
- Mesterhazy, A. Resistance of corn to Fusarium ear rot and its relation to seedling resistance. Phytopath. Z. (J. Phytopathol.) 1982, 103, 218–231. [Google Scholar] [CrossRef]
- Mesterhazy, A. Relationship between resistance to stalk rot and ear rot of corn influenced by rind resistance, premature death, and the rate of drying of the ear. Maydica 1983, 28, 425–437. [Google Scholar]
- Budakov, D.; Barošević, T.; Savić, Z.; Stojšin, V.; Grahovac, M.; Dudaš, T.; Bagi, F. Relationship between susceptibility to aflatoxin contamination and yield in maize hybrids. In Proceedings of the WMF Meets IUPAC, Belfast, UK, 14–16 October 2019. [Google Scholar]
- Reid, L.M.; Woldemariam, T.; Zhu, X.; Stewart, D.W.; Schaafsma, A.W. Effect of inoculation time and point of entry on disease severity in Fusarium graminearum, Fusarium verticillioides, or Fusarium subglutinans inoculated maize ears. Can. J. Plant Pathol. 2002, 24, 162–167. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Kovács, K. Breeding corn against fusarial stalk rot, ear rot and seedling blight. Acta Phytopath. Acad. Sci. Hung. 1986, 21, 231–249. [Google Scholar]
- Miedaner, T.; Bolduan, C.; Melchinger, A.E. Aggressiveness and mycotoxin production of eight isolates each of Fusarium graminearum and Fusarium verticillioides for ear rot on susceptible and resistant early maize inbred lines. Eur. J. Plant Pathol. 2010, 127, 113–123. [Google Scholar] [CrossRef]
- EC Commission Recommendation 2006/576/EC, 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2, and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Communities 2006, 229, 7–9.
- EC Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 20, 364–365.
- EC Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. OJL 2002, 30, 10.
- EC (2007) Commission regulation (EC) No 1126/2007 of 28 September 2007, amending regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union 2007, L255, 14–17.
- Hong, S.B.; Go, S.J.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 2005, 97, 1316–1329. [Google Scholar] [CrossRef]
- Toth, B.; Kaszonyi, G.; Bartok, T.; Varga, J.; Mesterhazy, A. Common resistance of wheat to members of the Fusarium graminearum species complex and F.culmorum. Plant Breed. 2008, 127, 1–8. [Google Scholar] [CrossRef]
- Baird, R.; Abbas, H.K.; Windham, G.; Williams, P.; Baird, S.; Ma, P.; Kelley, R.; Hawkins, L.; Scruggs, M. Identification of select fumonisin forming Fusarium species using PCR applications of the polyketide synthase gene, and its relationship to fumonisin production in vitro. Int. J. Mol. Sci. 2008, 9, 554–570. [Google Scholar] [CrossRef] [Green Version]
- Young, H.C. The toothpick method of inoculating corn for ear and stalk rots. Phytopathology 1943, 33, 16. [Google Scholar]
- Anonymous. Analytical Procedure of Bonafarm Babolna Feed UPLC-MS/MS Method, Approved by the National Accreditation Authority under the Code NAH-1254-14-1560/2016, Issued on 14 December 2016, Approval for Mycotoxins Was Issued under No. BBVM-111:2015. Available online: www.babolnatakarmany.hu/labor (accessed on 28 January 2022).
- Sváb, J. Biometriai Módszerek a Kutatásban (Methods for Biometrics in Research), 3rd ed.; Mezogazdasági Kiadó (Agr. Publ. House): Budapest, Hungary, 1981; p. 557. ISBN 963 231 0136. [Google Scholar]
- Weber, E. Grundriss der Biologischen Statistik. (Fundaments of the Biological Statistics); VEB Fisher Verlag: Jena, Germany, 1967. [Google Scholar]
- Logrieco, A.F.; Battilani, P.; Leggieri, C.M.; Haesaert, G.; Jiang, Y.; Lanubile, A.; Mahuku, G.; Mesterhazy, A.; Ortega-Beltran, A.; Pasti, M.A.; et al. Perspectives on global mycotoxin issues and management from the MycoKey Maize Working Group. Plant Dis. 2021, 105, 525–537. [Google Scholar] [CrossRef]
- Probst, C.; Bandyopadhyay, R.; Cotty, P.J. Diversity of aflatoxin-producing fungi and their impact on food safety in sub-Saharan Africa. Int. J. Food Microbiol. 2014, 174, 113–122. [Google Scholar] [CrossRef]
- Presello, D.A.; Botta, G.; Iglesias, J.; Eyherabide, G.H. Effect of disease severity on yield and grain fumonisin concentration of maize hybrids inoculated with Fusarium verticillioides. Crop Prot. 2008, 27, 572–576. [Google Scholar] [CrossRef]
- Lanubile, A.; Pasini, L.; Lo Pinto, M.; Battilani, P.; Prandini, A.; Marocco, A. Evaluation of broad spectrum sources of resistance to Fusarium verticillioides and advanced maize breeding lines. World Mycotoxin J. 2011, 4, 43–51. [Google Scholar] [CrossRef]
- Womack, E.D.; Williams, W.P.; Windham, G.L.; Xu, W. Mapping Quantitative Trait Loci associated with resistance to aflatoxin accumulation in maize inbred mp719. Front. Microbiol. 2020, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Schoeman, A.; Flett, B.C.; Janse van Rensburg, B.; Ncube, E.; Viljoen, A. Pathogenicity and toxigenicity of Fusarium verticillioides isolates collected from maize roots, stems, and ears in South Africa. Eur. J. Plant Pathol. 2018, 152, 677–689. [Google Scholar] [CrossRef]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, UK, 1971; pp. 1–237. [Google Scholar]
- Christensen, C.M.; Kaufmann, H.H. Grain Storage, the Role of Fungi in Quality Loss; Library of Congress Catalog Card Number: 70-76174; University of Minnesota Press: Minneapolis, MN, USA, 1969; 153p. [Google Scholar]
- Wit, M.; Waśkiewicz, A.; Goliński, P.; Chełkowski, J.; Warzecha, R.; Ochodzki, P.; Wakuliński, W. Occurrence of fumonisin FB1 in kernels and rachis of maize cobs infected by Fusarium verticillioides. Prog. Plant Prot. 2010, 50, 1832–1836. [Google Scholar]
- Leslie, J.F.; Moretti, A.; Mesterházy, Á.; Ameye, M.; Audenaert, K.; Singh, P.K.; Richard-Forget, F.; Chulze, S.N.; Del Ponte, E.M.; Chala, A.; et al. Key global actions for mycotoxin management in wheat and other small grains. Toxins 2021, 13, 725. [Google Scholar] [CrossRef] [PubMed]
- Ráduly, Z.; Szabó, L.; Madar, A.; Pócsi, I.; Csernoch, L. Toxicological and medical aspects of Aspergillus-derived mycotoxins entering the feed and food chain. Front. Microbiol. 2020, 10, 2908. [Google Scholar] [CrossRef] [Green Version]
- Miedaner, T.; Juroszek, P. Global warming and increasing maize cultivation demand comprehensive efforts in disease and insect resistance breeding in north-western Europe. Plant Pathol. 2021, 70, 1032–1046. [Google Scholar] [CrossRef]
- Medina, A.; Rodriguez, A.; Sultan, Y.; Magan, N. Climate change factors and Aspergillus flavus: Effects on gene expression, growth, and aflatoxin production. World Mycotoxin J. 2015, 8, 171–179. [Google Scholar] [CrossRef]
- Damianidis, D.; Ortiz, B.V.; Bowen, K.L.; Windham, G.L.; Hoogenboom, G.; Hagan, A.; Knappenberger, T.; Abbas, H.K.; Scully, B.T.; Mourtzinis, S. Minimum temperature, rainfall, and agronomic management impacts on corn grain aflatoxin contamination. Agron. J. 2018, 110, 1697–1708. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.L.; Williams, W.P.; Windham, G.L.; Menkir, A.; Chen, Z.-Y. Evaluation of African bred maize germplasm lines for resistance to aflatoxin accumulation. Agronomy 2016, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Royon, F. Mycotoxins Are Concentrated in Distillers’ Grains. Hoards-Dairyman, Feeding. 2016, p. 731. Available online: https://www.slideshare.net/FernandoDiazRoyonDVM/mycotoxins-are-concentrated-in-distillers-grain (accessed on 28 January 2022).
- Mesterhazy, A. How to Avoid Toxigenic Problems in Animal Husbandry? Open Access J. Vet. Sci. Res. 2020, 5, 1–3. [Google Scholar] [CrossRef]
- Rose, L.J.; Sheila Okoth, S.; Flett, B.C.; van Rensburg, B.J.; Viljoen, A. Preharvest Management Strategies and Their Impact on Mycotoxigenic Fungi and Associated Mycotoxins, Mycotoxins-Impact and Management Strategies; Njobeh, P.B., Stepman, F., Eds.; Intech Open: London, UK, 2019; Available online: https://www.intechopen.com/books/mycotoxins-impact-and-management-strategies/preharvest-management-strategies-and-their-impact-on-mycotoxigenic-fungi-and-associated-mycotoxins (accessed on 28 January 2022).
- Zhang, Y.-H.; Caupert, J. Survey of Mycotoxins in U.S. Distiller’s Dried Grains with Solubles from 2009 to 2011. J. Agric. Food Chem. 2012, 60, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; David, A. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Mycotoxins from the Field to the Table. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parts of Row | Rows/Isolates | Check | ||
|---|---|---|---|---|
| 2 | Fg *. 2 | Fv. 2. | Af. 2. | Check2 |
| way | ||||
| 1 | Fg. 1 | Fv. 1 | Af. 1. | Check1 |
| Row 1 | Row 2 | Row 3 | Row 4 | |
| Mycotoxin | Precursor Ion | Product Ions | Lens Parameters | Time Parameters | |||||
|---|---|---|---|---|---|---|---|---|---|
| Q1 Mass (Da) | Adduct | Daughter | Q3 Mass (Da) | DP (V) | CE (V) | CXP (V) | Dwell (ms) | RT (min) | |
| DON | 355 | [M+AcO]− | 1 | 59.2 | −40 | −40 | −8 | 150 | 4.32 |
| [M+AcO]− | 2 | 295.2 | −40 | −16 | −14 | 150 | |||
| AFB1 | 313 | [M+H]+ | 1 | 285.0 | 176 | 35 | 14 | 20 | 5.47 |
| [M+H]+ | 2 | 241.0 | 176 | 53 | 14 | 20 | |||
| FB1+ | 722 | [M+H]+ | 1 | 704.0 | 241 | 41 | 42 | 20 | 5.92 |
| [M]+ | 2 | 352.0 | 231 | 49 | 24 | 20 | |||
| FB2 | 706 | [M+H]+ | 1 | 688.0 | 216 | 39 | 36 | 20 | 6.34 |
| [M]+ | 2 | 336.0 | 221 | 51 | 26 | 20 | |||
| Hybrid | Toxigenic Species | Check | Mean Ear Rot | Mean of Rankings | Variance in Rankings | ||
|---|---|---|---|---|---|---|---|
| Fg | Fv | Af | |||||
| Szegedi 521 | 5.50 | 0.20 | 0.11 | 0.42 | 1.56 | 5.00 | 18.67 |
| Korimbos | 8.48 | 0.04 | 0.05 | 0.12 | 2.17 | 1.75 | 0.25 |
| DKC 4590 | 11.18 | 0.36 | 0.15 | 0.50 | 3.05 | 9.50 | 25.67 |
| P9537 | 11.64 | 0.32 | 0.14 | 0.25 | 3.09 | 7.25 | 11.58 |
| P9241 | 12.21 | 0.25 | 0.10 | 0.23 | 3.20 | 5.00 | 0.67 |
| 4517 | 12.09 | 0.87 | 0.17 | 0.60 | 3.43 | 11.75 | 28.25 |
| DKC 4717 | 13.29 | 0.45 | 0.24 | 0.35 | 3.58 | 10.50 | 9.67 |
| Cardixxio Duo | 13.88 | 0.23 | 0.29 | 0.26 | 3.66 | 8.25 | 17.58 |
| DKC 5542 | 14.04 | 0.32 | 0.10 | 0.37 | 3.71 | 8.00 | 11.33 |
| Siló Star | 15.05 | 0.26 | 0.16 | 0.20 | 3.92 | 7.25 | 12.25 |
| P9903 | 15.27 | 0.32 | 0.21 | 0.44 | 4.06 | 11.25 | 2.92 |
| DKC 4943 | 15.22 | 0.32 | 0.39 | 0.32 | 4.06 | 12.00 | 14.00 |
| Fornad | 14.34 | 1.18 | 0.29 | 0.63 | 4.11 | 14.50 | 13.67 |
| P9911 | 19.18 | 0.30 | 0.13 | 0.49 | 5.03 | 10.25 | 14.92 |
| DKC 5830 | 20.38 | 0.27 | 0.30 | 0.21 | 5.29 | 10.50 | 35.00 |
| DKC 4541 | 19.79 | 1.12 | 0.35 | 0.68 | 5.49 | 16.50 | 1.67 |
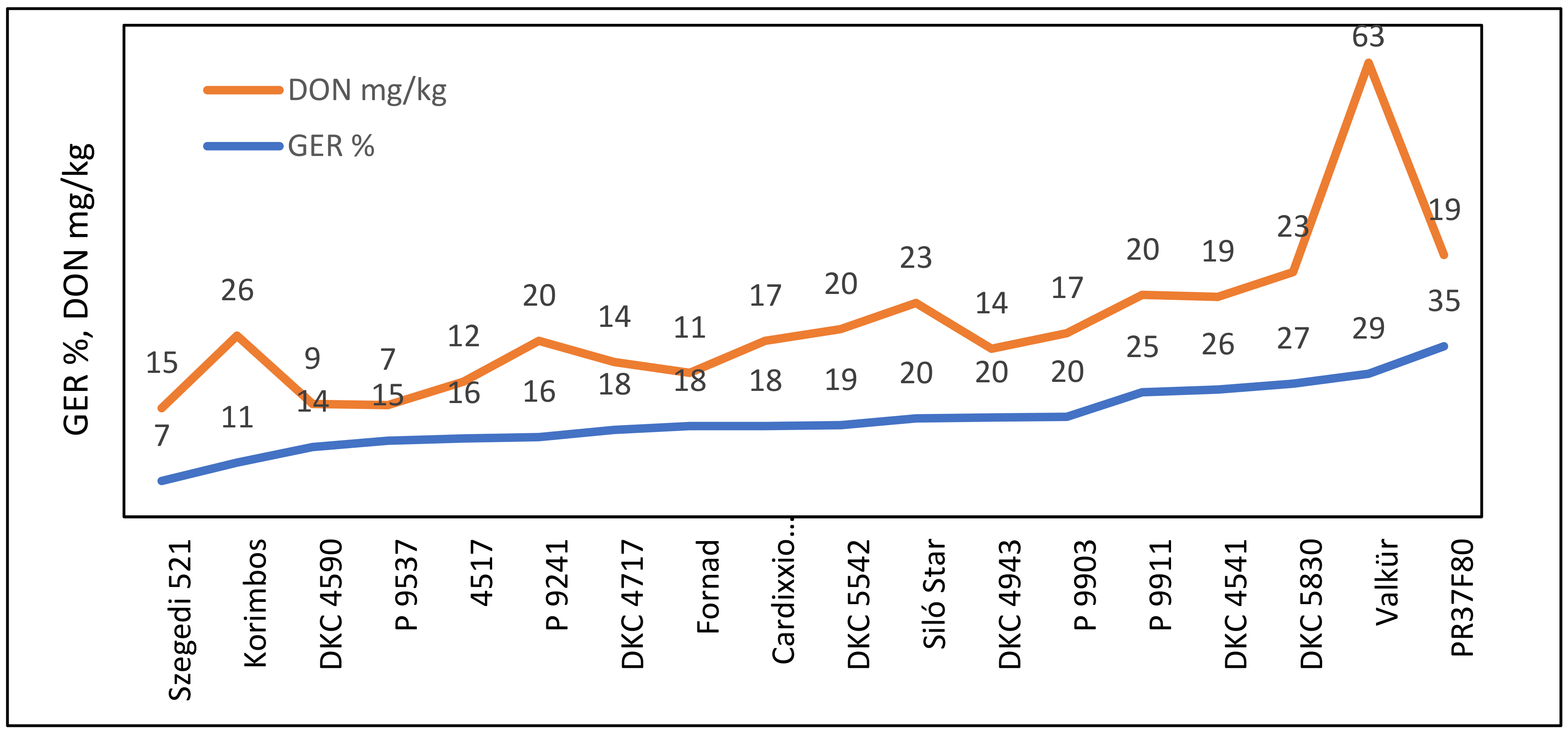
| Valkür | 21.83 | 0.14 | 0.03 | 0.03 | 5.51 | 5.25 | 61.58 |
| PR37F80 | 26.20 | 0.86 | 0.48 | 0.58 | 7.03 | 16.50 | 3.00 |
| Mean | 14.98 | 0.43 | 0.20 | 0.37 | 4.00 | 9.50 | 15.70 |
| LSD 5% | 7.69 | 0.45 | 0.18 | 0.25 | 1.92 | ||
| Correlations | Fg | Fv | Af | Check | Mean | Ranks | |
| Fv | 0.3190 | ||||||
| Af | 0.5127 * | 0.5746 * | |||||
| Check | 0.1234 | 0.8271 *** | 0.4918 * | ||||
| Mean | 0.994 *** | 0.4128 | 0.5691 * | 0.2187 | |||
| Ranks | 0.5810 * | 0.8337 *** | 0.8299 *** | 0.7893 *** | 0.6577 ** | ||
| Variance | 0.2106 | −0.1995 | −0.2822 | −0.3369 | 0.1702 | −0.1790 | |
| *** p = 0.001, ** p = 0.01, * p = 0.05 | Fg = F. graminearum, Fv = F. verticillioides, Af = . flavus, Bold names: good general resistance | ||||||
| Risk group | Low | Low to Medium | Medium to High | High | |||
| Hybrid | Tox. Species, Ear Rot % | Toxins | Control | FAO | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fg | Fv | Af | DON mg/kg | FB1+B2 mg/kg | AFB1 μg/kg | F. Ear Rot % | Af Ear Rot % | DON mg/kg | FB1+B2 mg/kg | AFB1 μg/kg | No. | |
| Szegedi 521 | 5.50 | 0.20 | 0.11 | 14.80 | 2.70 | 289.75 | 0.42 | 0.01 | 0.15 | 0.65 | 90.50 | 560 |
| Korimbos | 8.48 | 0.04 | 0.05 | 25.8 | 0.81 | 298.75 | 0.12 | 0.00 | 2.40 | 0.42 | 27.50 | 575 |
| DKC 4590 | 11.18 | 0.36 | 0.15 | 8.75 | 2.31 | 21.50 | 0.50 | 0.05 | 0.19 | 2.76 | 1.65 | 360 |
| P9537 | 11.64 | 0.32 | 0.14 | 7.27 | 3.05 | 21.75 | 0.25 | 0.00 | 0.00 | 1.53 | 11.00 | 370 |
| 4517 | 12.09 | 0.87 | 0.17 | 11.58 | 11.44 | 77.75 | 0.60 | 0.00 | 0.00 | 7.05 | 32.00 | 520 |
| P9241 | 12.21 | 0.25 | 0.10 | 19.57 | 1.79 | 19.75 | 0.23 | 0.00 | 0.00 | 0.71 | 14.00 | 350 |
| DKC 4717 | 13.29 | 0.45 | 0.24 | 13.81 | 1.11 | 40.25 | 0.35 | 0.00 | 0.05 | 1.92 | 29.50 | 390 |
| Cardixxio Duo | 13.88 | 0.23 | 0.29 | 17.32 | 1.22 | 29.50 | 0.26 | 0.00 | 0.18 | 0.18 | 2.50 | 470 |
| DKC 5542 | 14.04 | 0.32 | 0.10 | 19.54 | 1.29 | 338.50 | 0.37 | 0.00 | 0.00 | 2.71 | 33.00 | 540 |
| Fornad | 14.34 | 1.18 | 0.29 | 10.84 | 4.47 | 137.25 | 0.63 | 0.09 | 1.05 | 1.55 | 6.50 | 420 |
| Siló Star | 15.05 | 0.26 | 0.16 | 23.47 | 1.06 | 202.25 | 0.20 | 0.00 | 1.30 | 0.31 | 38.90 | 490 |
| DKC 4943 | 15.22 | 0.32 | 0.39 | 13.98 | 0.94 | 44.25 | 0.32 | 0.05 | 0.32 | 0.43 | 21.50 | 400 |
| P9903 | 15.27 | 0.32 | 0.21 | 17.04 | 1.69 | 38.50 | 0.44 | 0.00 | 0.10 | 0.41 | 2.50 | 390 |
| P9911 | 19.18 | 0.30 | 0.13 | 19.79 | 6.47 | 59.25 | 0.49 | 0.17 | 0.00 | 1.60 | 10.50 | 450 |
| DKC 4541 | 19.79 | 1.12 | 0.35 | 18.85 | 3.18 | 57.50 | 0.68 | 0.04 | 0.05 | 1.30 | 8.50 | 370 |
| DKC 5830 | 20.38 | 0.27 | 0.30 | 22.72 | 0.98 | 322.00 | 0.21 | 0.00 | 0.00 | 0.09 | 2.50 | 560 |
| Valkür | 21.83 | 0.14 | 0.03 | 63.27 | 1.64 | 13.00 | 0.03 | 0.00 | 0.61 | 0.51 | 5.00 | 730 |
| PR37F80 | 26.20 | 0.86 | 0.47 | 18.57 | 7.66 | 135.25 | 0.58 | 0.00 | 0.00 | 0.91 | 19.00 | 420 |
| Mean | 14.98 | 0.43 | 0.20 | 19.28 | 2.99 | 119.26 | 0.37 | 0.02 | 0.36 | 1.39 | 19.81 | |
| LSD 5% | 7.69 | 0.45 | 0.18 | 21.5 | 4.00 | 66.29 | 0.25 | |||||
| Risk group | Low | Low to medium | Medium to high | High | ||||||||
| Hybrid | Toxigenic Species, Ear Rot % | Ranks | Ranks | ||||
|---|---|---|---|---|---|---|---|
| Fg+ | Fv | Af | Check | Mean | Mean | Variance | |
| Konfites | 10.15 | 0.61 | 0.35 | 0.21 | 2.83 | 11.00 | 46.00 |
| ES Harmonium | 11.49 | 0.23 | 0.15 | 0.25 | 3.03 | 7.00 | 38.67 |
| Sy Talisman | 13.50 | 0.40 | 0.27 | 0.27 | 3.61 | 9.75 | 36.92 |
| Korimbos | 14.35 | 0.14 | 0.12 | 0.08 | 3.67 | 3.25 | 2.25 |
| P0725 | 15.91 | 0.45 | 0.11 | 0.17 | 4.16 | 7.00 | 8.67 |
| Koregraf | 16.62 | 0.42 | 0.24 | 0.10 | 4.35 | 6.75 | 4.92 |
| DKC 5830 | 17.08 | 0.52 | 0.44 | 0.17 | 4.55 | 11.00 | 18.00 |
| ES Lagoon | 19.76 | 0.32 | 0.24 | 0.19 | 5.13 | 8.75 | 8.92 |
| Armagnac | 19.79 | 0.23 | 0.11 | 0.13 | 5.06 | 5.00 | 8.00 |
| Illango | 19.94 | 0.53 | 0.36 | 0.18 | 5.25 | 12.25 | 6.92 |
| P9718E | 21.86 | 0.49 | 0.25 | 0.09 | 5.67 | 9.25 | 17.58 |
| Kathedralis | 22.10 | 0.67 | 0.58 | 0.21 | 5.89 | 15.25 | 7.58 |
| DKC 4541 | 24.59 | 1.61 | 0.54 | 0.30 | 6.76 | 16.50 | 5.67 |
| Valkür | 28.78 | 0.10 | 0.05 | 0.05 | 7.25 | 4.25 | 42.25 |
| P9415 | 32.04 | 0.60 | 0.28 | 0.24 | 8.29 | 14.50 | 1.00 |
| SY Zephir | 33.92 | 0.37 | 0.19 | 0.16 | 8.66 | 9.00 | 22.67 |
| Kleopatras | 35.02 | 0.44 | 0.10 | 0.16 | 8.93 | 8.75 | 38.92 |
| Sy Zoan | 35.36 | 0.47 | 0.16 | 0.18 | 9.04 | 11.75 | 20.92 |
| Mean | 21.79 | 0.48 | 0.25 | 0.17 | 5.67 | 9.50 | 18.66 |
| LSD 5% | 8.60 | 0.55 | 0.15 | 0.08 | 2.16 | 5.98 | |
| Correlations | Fg + | Fv | Af | Check | Mean | Ranks | |
| Fv | 0.110 | ||||||
| Afl | −0.161 | 0.744 *** | |||||
| Check | −0.091 | 0.62 ** | 0.539 * | ||||
| Mean | 0.998 *** | 0.169 | −0.108 | −0.048 | |||
| Ranks, mean | 0.254 | 0.799 *** | 0.824 *** | 0.704 *** | 0.307 | ||
| Ranks, variance | −0.032 | −0.257 | −0.282 | 0.049 | −0.046 | −0.203 | |
| *** p = 0.001, ** p = 0.01, * p = 0.05, Fg+ = F. graminearum; Fv = F. verticillioides; Af = A. flavus. | |||||||
| Risk group: | Low | Low to medium | Medium to high | High | |||
| Hybrid | Ear Rot % Art+. | Toxin Content Art. | Ear Rot, Control | Toxin Content, Control | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fg % x | Fv % | Af % | DON mg/kg | FB1+B2 mg/kg | AFB11 μg/kg | F. Check | Asp. Check | DON mg/kg | FB1+B2 mg/kg | AFB1 μg/kg | FAO No. | |
| Konfites | 10.15 | 0.61 | 0.35 | 18.15 | 3.49 | 53 | 0.21 | 0.000 | 1.70 | 1.88 | 2 | 430 |
| ES Harmonium | 11.49 | 0.23 | 0.15 | 37.60 | 2.10 | 62 | 0.25 | 0.000 | 0.13 | 0.81 | 4 | 380 |
| Sy Talisman | 13.50 | 0.40 | 0.27 | 7.48 | 1.80 | 71 | 0.27 | 0.000 | 0.72 | 0.54 | 2 | 250 |
| Korimbos | 14.35 | 0.14 | 0.12 | 76.08 | 5.67 | 44 | 0.08 | 0.000 | 3.49 | 0.30 | 408 | 575 |
| P0725 | 15.91 | 0.45 | 0.11 | 28.58 | 4.12 | 613 | 0.17 | 0.005 | 0.00 | 0.12 | 794 | 560 |
| Koregraf | 16.62 | 0.42 | 0.24 | 30.72 | 2.47 | 37 | 0.10 | 0.000 | 0.00 | 0.30 | 352 | 410 |
| DKC 5830 | 17.08 | 0.52 | 0.44 | 47.02 | 3.25 | 245 | 0.17 | 0.000 | 2.18 | 5.63 | 0 | 560 |
| ES Lagoon | 19.76 | 0.32 | 0.24 | 38.87 | 3.11 | 896 | 0.19 | 0.005 | 0.00 | 2.23 | 20 | 460 |
| Armagnac | 19.79 | 0.23 | 0.11 | 25.27 | 0.87 | 98 | 0.13 | 0.005 | 2.33 | 1.58 | 9 | 490 |
| Illango | 19.94 | 0.53 | 0.36 | 46.75 | 2.62 | 1068 | 0.18 | 0.008 | 0.05 | 0.75 | 1143 | 530 |
| P9718E | 21.86 | 0.49 | 0.25 | 101.01 | 1.52 | 638 | 0.09 | 0.000 | 0.17 | 1.17 | 3 | 390 |
| Kathedralis | 22.10 | 0.67 | 0.58 | 13.38 | 3.76 | 361 | 0.21 | 0.000 | 0.27 | 1.73 | 0 | 490 |
| DKC 4541 | 24.59 | 1.61 | 0.54 | 48.77 | 4.00 | 174 | 0.30 | 0.005 | 3.59 | 3.06 | 0 | 370 |
| Valkür | 28.78 | 0.10 | 0.05 | 82.64 | 1.55 | 205 | 0.05 | 0.000 | 1.28 | 0.81 | 0 | 731 |
| P9415 | 32.04 | 0.60 | 0.28 | 49.14 | 1.38 | 38 | 0.24 | 0.000 | 0.58 | 2.32 | 0 | 350 |
| SY Zephir | 33.92 | 0.37 | 0.19 | 28.38 | 1.75 | 160 | 0.16 | 0.005 | 0.63 | 2.36 | 4 | 390 |
| Kleopatras | 35.02 | 0.44 | 0.10 | 74.22 | 1.81 | 310 | 0.16 | 0.000 | 0.35 | 2.31 | 114 | 630 |
| Sy Zoan | 35.36 | 0.47 | 0.16 | 96.35 | 6.66 | 1258 | 0.18 | 0.008 | 0.00 | 1.65 | 0 | 560 |
| Mean | 21.79 | 0.48 | 0.25 | 47.24 | 2.88 | 352 | 0.17 | 0.002 | 0.97 | 1.64 | 159 | |
| LSD 5% | 8.60 | 0.55 | 0.15 | 54.90 | 3.789 | 528 | 0.08 | |||||
| Risk group: | Low | Low to medium | Medium to high | High | ||||||||
| Hybrid | Fg/DON | Fv/FB1+B2 | Af/AFB1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Fg % | mg/kg | DON/Fg % | Fv % | mg/kg | FB1+2/Fv % | Af % | mg/kg | AFB1/Af % | |
| Korimbos | 11.42 | 47.71 | 4.18 | 0.09 | 3.24 | 36.85 | 0.08 | 171.25 | 2091.60 |
| DKC 5830 | 18.73 | 32.03 | 1.71 | 0.40 | 2.11 | 5.31 | 0.37 | 283.63 | 776.61 |
| DKC 4541 | 22.19 | 31.47 | 1.42 | 1.37 | 3.59 | 2.63 | 0.45 | 115.88 | 258.82 |
| Valkür | 25.30 | 65.05 | 2.57 | 0.12 | 1.59 | 13.10 | 0.04 | 109.13 | 2816.13 |
| Mean | 13.08 | 44.06 | 3.37 | 0.49 | 2.63 | 5.34 | 0.22 | 169.97 | 759.81 |
| LSD 5% | 6.02 | ns | 0.59 | ns | 0.11 | ns | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesterhazy, A.; Szieberth, D.; Toldine, E.T.; Nagy, Z.; Szabó, B.; Herczig, B.; Bors, I.; Tóth, B. Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus. J. Fungi 2022, 8, 293. https://doi.org/10.3390/jof8030293
Mesterhazy A, Szieberth D, Toldine ET, Nagy Z, Szabó B, Herczig B, Bors I, Tóth B. Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus. Journal of Fungi. 2022; 8(3):293. https://doi.org/10.3390/jof8030293
Chicago/Turabian StyleMesterhazy, Akos, Denes Szieberth, Eva Tóth Toldine, Zoltan Nagy, Balázs Szabó, Beata Herczig, Istvan Bors, and Beata Tóth. 2022. "Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus" Journal of Fungi 8, no. 3: 293. https://doi.org/10.3390/jof8030293
APA StyleMesterhazy, A., Szieberth, D., Toldine, E. T., Nagy, Z., Szabó, B., Herczig, B., Bors, I., & Tóth, B. (2022). Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus. Journal of Fungi, 8(3), 293. https://doi.org/10.3390/jof8030293