
Peronosporales Species Associated with Strawberry Crown Rot in the Czech Republic


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Species Determination
2.3. Evaluation of Soil-Inhabiting Species Spectra of Peronosporales Present in Particular Localities
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jarvis, W.R. The infection of strawberry and raspberry fruits by Botrytis cinerea Fr. Ann. Appl. Biol. 1962, 50, 569–575. [Google Scholar] [CrossRef]
- Pegg, G.F. The impact of Verticillium diseases in agriculture. Phytopathol. Mediterr. 1984, 23, 176–192. [Google Scholar]
- Eikemo, H.; Stensvand, A.; Davik, J.; Tronsmo, A.M. Resistance to crown rot (Phytophthora cactorum) in strawberry cultivars and in offspring from crosses between cultivars differing in susceptibility to the disease. Ann. Appl. Biol. 2003, 142, 83–89. [Google Scholar] [CrossRef]
- Deutschmann, F. Eine Wurzelfäule an Erdbeeren, hervorgerufen durch Phytophthora cactorum (Leb. et Cohn) Schroet. Nachr. Dtsch. Pflanzenschutzd. 1954, 6, 7–9. [Google Scholar]
- Hantula, J.; Lilja, A.; Parikka, P. Genetic variation and host specificity of Phytophthora cactorum isolated in Europe. Mycol. Res. 1997, 101, 565–572. [Google Scholar] [CrossRef]
- Winterbottom, C.; Westerlund, F.; Mircetich, J.; Galper, L.; Welch, N. Evaluation of relative resistance of different strawberry cultivars to Phytophthora and Verticillium dahliae as a potential alternative to methyl bromide. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, Orlando, FL, USA, 4–6 November 1996; pp. 36-31–36-35. [Google Scholar]
- Delmas, C.E.; Mazet, I.D.; Jolivet, J.; Delière, L.; Delmotte, F. Simultaneous quantification of sporangia and zoospores in a biotrophic oomycete with an automatic particle analyzer: Disentangling dispersal and infection potentials. J. Microbiol. Methods 2014, 107, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Raftoyannis, Y.; Dick, M.W. Zoospore encystment and pathogenicity of Phytophthora and Pythium species on plant roots. Microbiol. Res. 2006, 161, 1–8. [Google Scholar] [CrossRef]
- Darmono, T.W.; Parke, J.L. Chlamydospores of Phytophthora cactorum: Their production, structure, and infectivity. Can. J. Bot. 1990, 68, 640–645. [Google Scholar] [CrossRef]
- Harris, D.C. Survival of Phytophthora syringae oospores in and on apple orchard soil. Trans. Br. Mycol. Soc. 1985, 85, 153–155. [Google Scholar] [CrossRef]
- Sneh, B.; McIntosh, D.L. Studies on the behavior and survival of Phytophthora cactorum in soil. Can. J. Bot. 1974, 52, 795–802. [Google Scholar] [CrossRef]
- Abad, Z.G.; Abad, J.A.; Coffey, M.D.; Oudemans, P.V.; Man in’t Veld, W.A.; de Gruyter, H.; Cunnington, J.; Louws, F.J. Phytophthora bisheria sp. nov., a new species identified in isolates from the Rosaceous raspberry, rose and strawberry in three continents. Mycologia 2008, 100, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Barboza, E.A.; Fonseca, M.E.N.; Boiteux, L.S.; Reis, A. First worldwide report of a strawberry fruit rot disease caused by Phytophthora capsici isolates. Plant Dis. 2017, 101, 259. [Google Scholar] [CrossRef]
- Hickman, C.J. The red core root disease of the strawberry caused by Phytophthora fragariae n.sp. J. Pomol. Hortic. Sci. 1941, 18, 89–118. [Google Scholar] [CrossRef]
- Irzykowska, L.; Irzykowski, W.; Jarosz, A.; Golebniak, B. Association of Phytophthora citricola with leather rot disease of strawberry. J. Phytopathol. 2005, 153, 680–685. [Google Scholar] [CrossRef]
- Parikka, P.K.; Vestberg, M.; Karhu, S.T.; Haikonen, T.; Hautsalo, J. Possibilities for biological control of red core (Phytophthora fragariae). VIII Int. Strawb. Symp. 2017, 1156, 751–756. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Uematsu, S.; Takeuchi, T.; Shirai, K.; Ishiguro, Y.; Suga, H.; Kageyama, K. Two new species, Phytophthora nagaii sp. nov. and P. fragariaefolia sp. nov., causing serious diseases on rose and strawberry plants, respectively, in Japan. J. Gen. Plant Pathol. 2014, 80, 348–365. [Google Scholar] [CrossRef]
- Rytkönen, A.; Lilja, A.; Vercauteren, A.; Sirkiä, S.; Parikka, P.; Soukainen, M.; Hantula, J. Identity and potential pathogenicity of Phytophthora species found on symptomatic Rhododendron plants in a Finnish nursery. Can. J. Plant Pathol. 2012, 34, 255–267. [Google Scholar] [CrossRef]
- Suzui, T.; Makino, T.; Ogoshi, A. Phytophthora rot of strawberry caused by Phytophthora nicotianae var. parasitica in Shizuoka. Jap. J. Phytopathol. 1980, 46, 169–178. [Google Scholar] [CrossRef] [Green Version]
- De Cock, A.W.; Lodhi, A.M.; Rintoul, T.L.; Bala, K.; Robideau, G.P.; Abad, Z.G.; Coffey, M.D.; Shahzad, S.; Lévesque, C.A. Phytopythium: Molecular phylogeny and systematics. Persoonia 2015, 34, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Uzuhashi, S.; Tojo, M.; Kakishima, M. Phylogeny of the genus Pythium and description of new genera. Mycoscience 2010, 51, 337–365. [Google Scholar] [CrossRef]
- Eden, M.A.; Hill, R.A. First record of Pythium mastophorum in New Zealand and its pathogenicity relative to other Pythium spp. N. Z. J. Crop Hortic. Sci. 1998, 26, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, Y.; Otsubo, K.; Watanabe, H.; Suzuki, M.; Nakayama, K.; Fukuda, T.; Fujinaga, M.; Suga, H.; Kageyama, K. Root and crown rot of strawberry caused by Pythium helicoides and its distribution in strawberry production areas of Japan. J. Gen. Plant Pathol. 2014, 80, 423–429. [Google Scholar] [CrossRef]
- Marin, M.V.; Seijo, T.; Mertely, J.; Peres, N.A. First report of crown rot caused by Phytopythium helicoides on strawberry in the Americas. Plant Dis. 2019, 103, 2696. [Google Scholar] [CrossRef]
- Mouden, N.; Benkirane, R.; Ouazzani Touhami, A.; Douira, A. Fungal species associated with collapsed strawberry plants cultivated in strawberries plantations in Morocco. Int. J. Curr. Res. 2016, 8, 29108–29117. [Google Scholar]
- Shennan, C.; Muramoto, J.; Koike, S.; Baird, G.; Fennimore, S.; Samtani, J.; Bolda, M.; Dara, S.; Daugovish, O.; Lazarovits, G.; et al. Anaerobic soil disinfestation is an alternative to soil fumigation for control of some soilborne pathogens in strawberry production. Plant Pathol. 2018, 67, 51–66. [Google Scholar] [CrossRef]
- Toljamo, A.R.; Granlund, L.J.; Kokko, H.I. DNA barcode identification and virulence testing of Phytophthora spp. and Pythium spp. isolated from soil of strawberry fields. VIII Int. Strawb. Symp. 2017, 1156, 727–734. [Google Scholar] [CrossRef]
- Watanabe, T. Pathogenicity of Pythium myriotylum isolated from strawberry roots in Japan. Ann. Phytopath. Soc. Jpn. 1977, 43, 306–309. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Hashimoto, K.; Sato, M. Pythium species associated with strawberry roots in Japan, and their role in the strawberry stunt disease. Phytopathology 1977, 67, 1324–1332. [Google Scholar] [CrossRef]
- Fang, X.L.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Severity of crown and root diseases of strawberry and associated fungal and oomycete pathogens in Western Australia. Australas. Plant Pathol. 2011, 40, 109–119. [Google Scholar] [CrossRef]
- Fang, X.L.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Comparisons of virulence of pathogens associated with crown and root diseases of strawberry in Western Australia with special reference to the effect of temperature. Sci. Hortic. 2011, 131, 39–48. [Google Scholar] [CrossRef]
- LaMondia, J. Strawberry black root rot. Adv. Strawb. Res. 2004, 23, 1–10. [Google Scholar]
- Martin, F.N. Strawberry root rot and the recovery of Pythium and Rhizoctonia spp. In Proceedings of the Annual International Research Conference on Methyl Bromide Alternatives and Emissions Reductions, San Diego, CA, USA, 1–4 November 1999. [Google Scholar]
- Burdon, J.J.; Chilvers, G.A. Epidemiology of damping-off disease (Pythium irregulare) in relation to density of Lepidium sativum seedlings. Ann. Appl. Biol. 1975, 81, 135–143. [Google Scholar] [CrossRef]
- Green, H.; Jensen, D.F. Disease progression by active mycelial growth and biocontrol of Pythium ultimum var. ultimum studied using a rhizobox system. Phytopathology 2000, 90, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redekar, N.R.; Eberhart, J.L.; Parke, J.L. Diversity of Phytophthora, Pythium, and Phytopythium species in recycled irrigation water in a container nursery. Phytobiomes J. 2019, 3, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Lookabaugh, E.C.; Ivors, K.L.; Shew, B.B. Mefenoxam sensitivity, aggressiveness, and identification of Pythium species causing root rot on floriculture crops in North Carolina. Plant Dis. 2015, 99, 1550–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Htun, Z.M.; Rotchanapreeda, T.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Sae-Chew, P.; Payattikul, P.; Yurayart, C.; Limsivilai, O.; et al. Loop-mediated isothermal amplification (LAMP) for identification of Pythium insidiosum. Int. J. Infect. Dis. 2020, 101, 149–159. [Google Scholar] [CrossRef]
- Olson, J.D.; Damicone, J.P.; Kahn, B.A. Identification and characterization of isolates of Pythium and Phytophthora spp. from snap beans with cottony leak. Plant Dis. 2016, 100, 1446–1453. [Google Scholar] [CrossRef]
- Radmer, L.; Anderson, G.; Malvick, D.M.; Kurle, J.E.; Rendahl, A.; Mallik, A. Pythium, Phytophthora, and Phytopythium spp. associated with soybean in Minnesota, their relative aggressiveness on soybean and corn, and their sensitivity to seed treatment fungicides. Plant Dis. 2017, 101, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Themann, K.; Werres, S. Baiting of Phytophthora sp. with the Rhododendron leaf test. In Proceedings of the First International Meeting on Phytophthoras in Forest and Wildland Ecosystems, Grants Pass, OR, USA, 30 August–3 September 1999. [Google Scholar]
- Balcì, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathol. 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Tsao, P.H. Factors affecting isolation and quantitation of Phytophthora from soil. In Phytophthora: Its Biology, Taxonomy, Ecology, and Pathology, 1st ed.; Erwin, D.C., Bartnicki-Garcia, S., Tsao, P.H., Eds.; American Phytopathological Society: St. Paul, MN, USA, 1983; pp. 219–236. [Google Scholar]
- Kageyama, K. Molecular taxonomy and its application to ecological studies of Pythium species. J. Gen. Plant Pathol. 2014, 80, 314–326. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lévesque, C.A.; de Cock, A.W. Molecular phylogeny and taxonomy of the genus. Pythium. Mycol. Res. 2004, 108, 1363–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkowski, J.E.; de Boer, R.F.; Norng, S.; Thomson, F.; Minchinton, E.J. Pythium species associated with root rot complex in winter-grown parsnip and parsley crops in south eastern Australia. Australas. Plant Pathol. 2013, 42, 403–411. [Google Scholar] [CrossRef]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Community composition and distribution of Phytophthora species across adjacent native and non-native forests of South Africa. Fungal Ecol. 2018, 36, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Khaliq, I.; Hardy, G.E.S.J.; White, D.; Burgess, T.I. eDNA from roots: A robust tool for determining Phytophthora communities in natural ecosystems. FEMS Microbiol. Ecol. 2018, 94, fiy048. [Google Scholar] [CrossRef] [Green Version]
- Matsiakh, I.; Kramarets, V.; Cleary, M. Occurrence and diversity of Phytophthora species in declining broadleaf forests in western Ukraine. For. Pathol. 2021, 51, e12662. [Google Scholar] [CrossRef]
- Nechwatal, J.; Bakonyi, J.; Cacciola, S.O.; Cooke, D.E.L.; Jung, T.; Nagy, Z.Á.; Vannini, A.; Vettraino, A.M.; Brasier, C.M. The morphology, behaviour and molecular phylogeny of Phytophthora taxon Salixsoil and its redesignation as Phytophthora lacustris sp. nov. Plant Pathol. 2013, 62, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First report of root rot caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Scanu, B.; Linaldeddu, B.; Pérez-Sierra, A.; Deidda, A.; Franceschini, A. Phytophthora ilicis as a leaf and stem pathogen of Ilex aquifolium in Mediterranean islands. Phytopathol. Mediterr. 2014, 53, 480–490. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide, 1st ed.; American Phytopathological Society: St Paul, MN, USA, 1996; p. 592. [Google Scholar]
- Mostowfizadeh-Ghalamfarsa, R.; Panabieres, F.; Banihashemi, Z.; Cooke, D.E.L. Phylogenetic relationship of Phytophthora cryptogea Pethybr. & Laff and P. drechsleri Tucker. Fungal Biol. 2010, 114, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Allasia, V.; Venard, P.; Notter, S.; Vernière, C.; Panabières, F. Intraspecific variation in Phytophthora citrophthora from citrus trees in Eastern Corsica. Eur. J. Plant Pathol. 2003, 109, 791–805. [Google Scholar] [CrossRef]
- Nemec, S. Histopathology of Pythium infected strawberry roots. Can. J. Bot. 1972, 50, 1091–1096. [Google Scholar] [CrossRef]
- Van der Plaats-Niterink, A.J. Monograph of the Genus Pythium, 1st ed.; Centraalbureau voor Schimmelcultures: Baarn, The Netherlands, 1981; p. 244. [Google Scholar]
- Badali, F.; Abrinbana, M.; Abdollahzadeh, J.; Khaledi, E. Molecular and morphological taxonomy of Pythium species isolated from soil in West Azarbaijan province (N.W. Iran). Rostaniha 2016, 17, 78–91. [Google Scholar] [CrossRef]
- De Cock, A.W.A.M.; Lévesque, C.A. New species of Pythium and Phytophthora. Stud. Mycol. 2004, 50, 481–487. [Google Scholar]
- Ekwamu, A.; Mukalazi, J.; Buruchara, R.A. Pythium Species in Bean Fields in Uganda, 1st ed.; LAP LAMBERT Academic Publishing: Chisinau, Moldova, 2013; p. 136. [Google Scholar]
- Hosseini Badrbani, A.; Abbasi, S.; Bolboli, Z.; Jamali, S.; Sharifi, R. Morphological and molecular characterization of Oomycetes associated with root and crown rot of cucurbits in Kermanshah province, Iran. Mycol. Iran. 2018, 5, 15–27. [Google Scholar] [CrossRef]
- Chen, J.J.; Yu, J.; Zhou, Y.H.; Yang, H.J. Pythium glomeratum and Py. nodosum, two new records from China. J. Microbiol. Exp. 2020, 8, 24–28. [Google Scholar] [CrossRef]
- Le, D.P.; Smith, M.K.; Aitken, E.A.B. An assessment of Pythium spp. associated with soft rot disease of ginger (Zingiber officinale) in Queensland, Australia. Australas. Plant Pathol. 2016, 45, 377–387. [Google Scholar] [CrossRef]
- Li, Y.P.; You, M.P.; Barbetti, M.J. Species of Pythium associated with seedling root and hypocotyl disease on common bean (Phaseolus vulgaris) in Western Australia. Plant Dis. 2014, 98, 1241–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLeod, A.; Botha, W.J.; Meitz, J.C.; Spies, C.F.; Tewoldemedhin, Y.T.; Mostert, L. Morphological and phylogenetic analyses of Pythium species in South Africa. Mycol. Res. 2009, 113, 933–951. [Google Scholar] [CrossRef]
- Nechwatal, J.; Oßwald, W.F. Pythium montanum sp. nov., a new species from a spruce stand in the Bavarian Alps. Mycol. Prog. 2003, 2, 73–80. [Google Scholar] [CrossRef]
- Paul, B.; Galland, D.; Bhatnagar, T.; Dulieu, H. A new species of Pythium isolated from the Burgundy region in France. FEMS Microbiol. Lett. 1998, 158, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, S.K.; Zhou, Y.H.; Lamour, K. Oomycetes baited from streams in Tennessee 2010–2012. Mycologia 2013, 105, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Weiland, J.E.; Beck, B.R.; Davis, A. Pathogenicity and virulence of Pythium species obtained from forest nursery soils on Douglas-fir seedlings. Plant Dis. 2013, 97, 744–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belbahri, L.; McLeod, A.; Paul, B.; Calmin, G.; Moralejo, E.; Spies, C.F.J.; Botha, W.J.; Clemente, A.; Descals, E.; Sánchez-Hernández, E.; et al. Intraspecific and within-isolate sequence variation in the ITS rRNA gene region of Pythium mercuriale sp. nov. (Pythiaceae). FEMS Microbiol. Lett. 2008, 284, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouket, A.C.; Babai-Ahari, A.; Arzanlou, M.; Tojo, M. Morphological and molecular characterization of Phytopythium litorale and Pp. oedochilum from Iran. Nova Hedwig. 2015, 102, 257–270. [Google Scholar] [CrossRef]
- Javadi, N.; Sharifnabi, B. Phytopythium litorale, the causal agent of almond root and crown rot in Iran. In Proceedings of the 22nd Iranian Plant Protection Congress, Karaj, Iran, 27–30 August 2016. [Google Scholar]
- Nam, B.; Choi, Y.J. Phytopythium and Pythium species (Oomycota) isolated from freshwater environments of Korea. Mycobiology 2019, 47, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Redekar, N.R.; Bourret, T.B.; Eberhart, J.L.; Johnson, G.E.; Pitton, B.J.L.; Haver, D.L.; Oki, L.R.; Parke, J.L. The population of oomycetes in a recycled irrigation water system at a horticultural nursery in southern California. Water Res. 2020, 183, 116050. [Google Scholar] [CrossRef]
- Rezaei, S.; Abrinbana, M.; Ghosta, Y. Taxonomic and pathogenic characterization of Phytopythium species from West Azarbaijan, Iran, and description of two new species. Mycologia 2021, 113, 612–628. [Google Scholar] [CrossRef] [PubMed]
- Lischke, H.; Guisan, A.; Fischlin, A.; Williams, J.; Bugmann, H. Vegetation responses to climate change in the Alps: Modeling studies. In Views from the Alps. Regional Perspectives on Climate Change, 1st ed.; Cebon, P., Dahinden, U., Davies, H., Imboden, D., Jaeger, C.C., Eds.; MIT Press: Cambridge, MA, USA, 1998; pp. 309–350. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Pánek, M.; Střížková, I.; Zouhar, M.; Kudláček, T.; Tomšovský, M. Mixed-mating model of reproduction revealed in European Phytophthora cactorum by ddRADseq and effector gene sequence data. Microorganisms 2021, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Pánek, M.; Fér, T.; Mráček, J.; Tomšovský, M. Evolutionary relationships within the Phytophthora cactorum species complex in Europe. Fungal Biol. 2016, 120, 836–851. [Google Scholar] [CrossRef] [PubMed]
- Hantula, J.; Lilja, A.; Nuorteva, H.; Parikka, P.; Werres, S. Pathogenicity, morphology and genetic variation of Phytophthora cactorum from strawberry, apple, rhododendron, and silver birch. Mycol. Res. 2000, 104, 1062–1068. [Google Scholar] [CrossRef]
- Bhat, R.G.; Colowit, P.M.; Tai, T.H.; Aradhya, M.K.; Browne, G.T. Genetic and pathogenic variation in Phytophthora cactorum affecting fruit and nut crops in California. Plant Dis. 2006, 90, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eikemo, H.; Klemsdal, S.S.; Riisberg, I.; Bonants, P.; Stensvand, A.; Tronsmo, A.M. Genetic variation between Phytophthora cactorum isolates differing in their ability to cause crown rot in strawberry. Mycol. Res. 2004, 108, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, D.E.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Christova, P.; Lyubenova, A.; Kostov, K.; Slavov, S. Diversity and pathogenicity of Phytophthora species, isolated from Osam River. J. Mt. Agric. Balk. 2018, 21, 179–191. [Google Scholar]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef] [Green Version]
- Ilieva, E.; Jamart, G.; Kamoen, O. Characterisation of some isolates of Phytophthora cryptogea. Parasitica 1992, 48, 113–122. [Google Scholar]
- Meszka, B.; Michalecka, M. Identification of Phytophthora spp. isolated from plants and soil samples on strawberry plantations in Poland. J. Plant Dis. Prot. 2016, 123, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora species in protected natural areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Olson, H.A.; Benson, D.M. Host specificity and variations in aggressiveness of North Carolina isolates of Phytophthora cryptogea and P. drechsleri in greenhouse ornamental plants. Plant Dis. 2013, 97, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, M.; Gerhardson, B. Isolates of Phytophthora cryptogea pathogenic to wheat and some other crop plants. J. Phytopathol. 1990, 129, 303–315. [Google Scholar] [CrossRef]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Phytophthora beyond agriculture. Annu. Rev. Phytopathol. 2012, 50, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Cooke, D.E.; Duncan, J.M.; Hansen, E.M. Multiple new phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides–P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Schoebel, C.N.; Stewart, J.; Grünwald, N.J.; Rigling, D.; Prospero, S. Population history and pathways of spread of the plant pathogen Phytophthora plurivora. PLoS ONE 2014, 9, e85368. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.H.; Chang, M.S.; Su, H.J. Isolates of Phytophthora cinnamomi from Taiwan as evidence for an Asian origin of the species. Trans. Br. Mycol. Soc. 1978, 71, 496–499. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, M.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A survey in natural forest ecosystems of Vietnam reveals high diversity of both new and described Phytophthora taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Lilja, A.; Rytkönen, A.; Hantula, J.; Müller, M.; Kurkela, T. Phytophthora cactorum, P. ramorum, P. plurivora, Melampsoridium hiratsukamum, Dothiostroma septosporum and Chalara fraxinea, non-native pathogens in Finland. In Proceedings of the 7th Meeting of IUFRO Working Party 7.03.04 Diseases and Insects in Forest Nurseries, Hilo, HI, USA, 13–17 July 2010; pp. 55–62. [Google Scholar]
- Laviola, C.; Somma, V.; Evola, C. Present status of Phytophthora species in the Mediterranean area, especially in relation to citrus. EPPO Bull. 1990, 20, 1–9. [Google Scholar] [CrossRef]
- Pánek, M.; Střížková, I. A comparison of the virulence of selected Pythium, Globisporangium, Phytopythium and Phytophthora species against strawberry plants. J. Plant Dis. Prot. 2021, 128, 1447–1458. [Google Scholar] [CrossRef]
- Sutton, J.C.; Sopher, C.R.; Owen-Going, T.N.; Liu, W.; Grodzinski, B.; Hall, J.C.; Benchimol, R.L. Etiology and epidemiology of Pythium root rot in hydroponic crops: Current knowledge and perspectives. Summa Phytopathol. 2006, 32, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Lugauskas, A.; Repečkiene, J.; Uselis, N.; Rašinskiene, A. Problems on a longtime strawberry growing in one plot. Hortum Cultus 2003, 2, 59–68. [Google Scholar]
- Rojas, J.A.; Jacobs, J.L.; Napieralski, S.; Karaj, B.; Bradley, C.A.; Chase, T.; Esker, P.D.; Giesler, L.J.; Jardine, D.J.; Malvick, D.K.; et al. Oomycete species associated with soybean seedlings in North America—part I: Identification and pathogenicity characterization. Phytopathology 2017, 107, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Zitnick-Anderson, K.K.; Nelson, B.D., Jr. Identification and pathogenicity of Pythium on soybean in North Dakota. Plant Dis. 2015, 99, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Granke, L.L.; Hausbeck, M.K. Effects of temperature, concentration, age, and algaecides on Phytophthora capsici zoospore infectivity. Plant Dis. 2010, 94, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littrell, R.H.; McCarter, S.M. Effect of soil temperature on virulence of Pythium aphanidermatum and Pythium myriotylum to rye and tomato. Phytopathology 1970, 60, 704–707. [Google Scholar] [CrossRef]
- Grove, G.G.; Madden, L.V.; Ellis, M.A. Splash dispersal of Phytophthora cactorum from infected strawberry fruit. Phytopathology 1985, 75, 611–615. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S. Taxonomic notes on three Pythium species. Trans. Br. Mycol. Soc. 1986, 86, 659–663. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Chen, W.D.; Yang, X.B. Occurrence of Pythium species in long-term maize and soybean monoculture and maize/soybean rotation. Mycol. Res. 1998, 102, 1450–1452. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Li, Y.Q.; Fan, H.; Wang, Y.C.; Zheng, X.B. Molecular detection of Phytophthora capsici in infected plant tissues, soil and water. Plant Pathol. 2006, 55, 770–775. [Google Scholar] [CrossRef]
- Van Poucke, K.; Haegeman, A.; Goedefroit, T.; Focquet, F.; Leus, L.; Jung, M.H.; Nave, C.; Redondo, M.A.; Husson, C.; Kostov, K.; et al. Unravelling hybridization in Phytophthora using phylogenomics and genome size estimation. IMA Fungus 2021, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Martin, F. Meiotic instability of Pythium sylvaticum as demonstrated by inheritance of nuclear markers and karyotype analysis. Genetics 1995, 139, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, H.; Zhou, Q.; Qian, K.; van der Lee, T.; Huang, S. Changing ploidy as a strategy: The Irish potato famine pathogen shifts ploidy in relation to its sexuality. Mol. Plant-Microbe Interact. 2017, 30, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Christova, P.K.; Kostov, K.V.; Lyubenova, A.B.; Slavov, S.B. A new hybrid of Phytophthora from Southeast Europe. Mycologia 2021, 113, 734–747. [Google Scholar] [CrossRef]
- Sheltzer, J.M.; Blank, H.M.; Pfau, S.J.; Tange, Y.; George, B.M.; Humpton, T.J.; Brito, I.L.; Hiraoka, Y.; Niwa, O.; Amon, A. Aneuploidy drives genomic instability in yeast. Science 2011, 333, 1026–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaver, A.C.; Dombrowski, P.G.; Sweeney, J.Y.; Treis, T.; Zappala, R.M.; Sniegowski, P.D. Fitness evolution and the rise of mutator alleles in experimental Escherichia coli populations. Genetics 2002, 162, 557–566. [Google Scholar] [CrossRef]
- Sniegowski, P.D.; Gerrish, P.J.; Lenski, R.E. Evolution of high mutation rates in experimental populations of E. coli. Nature 1997, 387, 703–705. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.K.; Leibler, S. Benefits of phenotypic plasticity for population growth in varying environments. Proc. Natl. Acad. Sci. USA 2018, 115, 12745–12750. [Google Scholar] [CrossRef] [Green Version]
- De Cara, M.; Palmero, D.; Durán, C.; Lacasa, C.; Santos, M.; Coffey, M.D.; Tello, J.C. Phytophthora parasitica showing host specificity and pathogenic ability on tomato and sweet pepper. In Microorganisms in Industry and Environment from Scientific and Industrial Research to Consumer Products, 1st ed.; Mendez-Vilas, A., Ed.; World Scientific Publishing Co.: Lisbon, Portugal, 2010; pp. 101–105. [Google Scholar]
- Wang, Y.; Meng, Y.; Zhang, M.; Tong, X.; Wang, Q.; Sun, Y.; Quan, J.; Govers, F.; Shan, W. Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host specificity. Mol. Plant Pathol. 2011, 12, 187–201. [Google Scholar] [CrossRef]
- McHau, G.R.A.; Coffey, M.D. An integrated study of morphological and isozyme patterns found within a worldwide collection of Phytophthora citrophthora and a redescription of the species. Mycol. Res. 1994, 98, 1291–1299. [Google Scholar] [CrossRef]
- Spies, C.F.; Meitz-Hopkins, J.C.; Langenhoven, S.D.; Pretorius, M.C.; McLeod, A. Two clonal lineages of Phytophthora citrophthora from citrus in South Africa represent a single phylogenetic species. Mycologia 2014, 106, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Pánek, M.; Helmer, Š.; Ali, A. Use of metalaxyl against some soil plant pathogens of the class Peronosporomycetes—A review and two case studies. Plant Prot. Sci. 2022, in press. [Google Scholar] [CrossRef]


{kind=link}
| Species Name | No. of Isolates Obtained | % of Particular Species on the Total No. of Isolates |
|---|---|---|
| Globisporangium irregulare | 3 | 1.71 |
| Globisporangium ultimum | 3 | 1.71 |
| Phytophthora bilorbang | 1 | 0.57 |
| Phytophthora cactorum | 113 | 64.57 |
| Phytophthora citrophthora | 2 | 1.14 |
| Phytophthora cryptogea | 1 | 0.57 |
| Phytophthora lacustris | 1 | 0.57 |
| Phytophthora plurivora | 1 | 0.57 |
| Phytopythium citrinum | 1 | 0.57 |
| Phytopythium mercuriale | 1 | 0.57 |
| Phytopythium montanum | 1 | 0.57 |
| Phytopythium vexans | 5 | 2.86 |
| Phytopythium litorale | 1 | 0.57 |
| Pythium aphanidermatum | 1 | 0.57 |
| Pythium conidiophorum | 1 | 0.57 |
| Pythium dissotocum complex | 20 | 11.43 |
| Pythium heterothallicum | 3 | 1.71 |
| Pythium intermedium | 4 | 2.29 |
| Pythium mamillatum | 2 | 1.14 |
| Pythium nodosum | 5 | 2.86 |
| Pythium perplexum | 1 | 0.57 |
| Pythium rostratifingens | 1 | 0.57 |
| Pythium salpingophorum | 1 | 0.57 |
| Pythium torulosum | 2 | 1.14 |
| Presence of Pythiaceae Pathogens | No. of Localities | % of Localities |
|---|---|---|
| Not any species found | 7 | 17.1 |
| Only P. cactorum found | 13 | 31.7 |
| Only other species than P. cactorum found | 9 | 22.0 |
| P. cactorum and other species found | 12 | 29.3 |
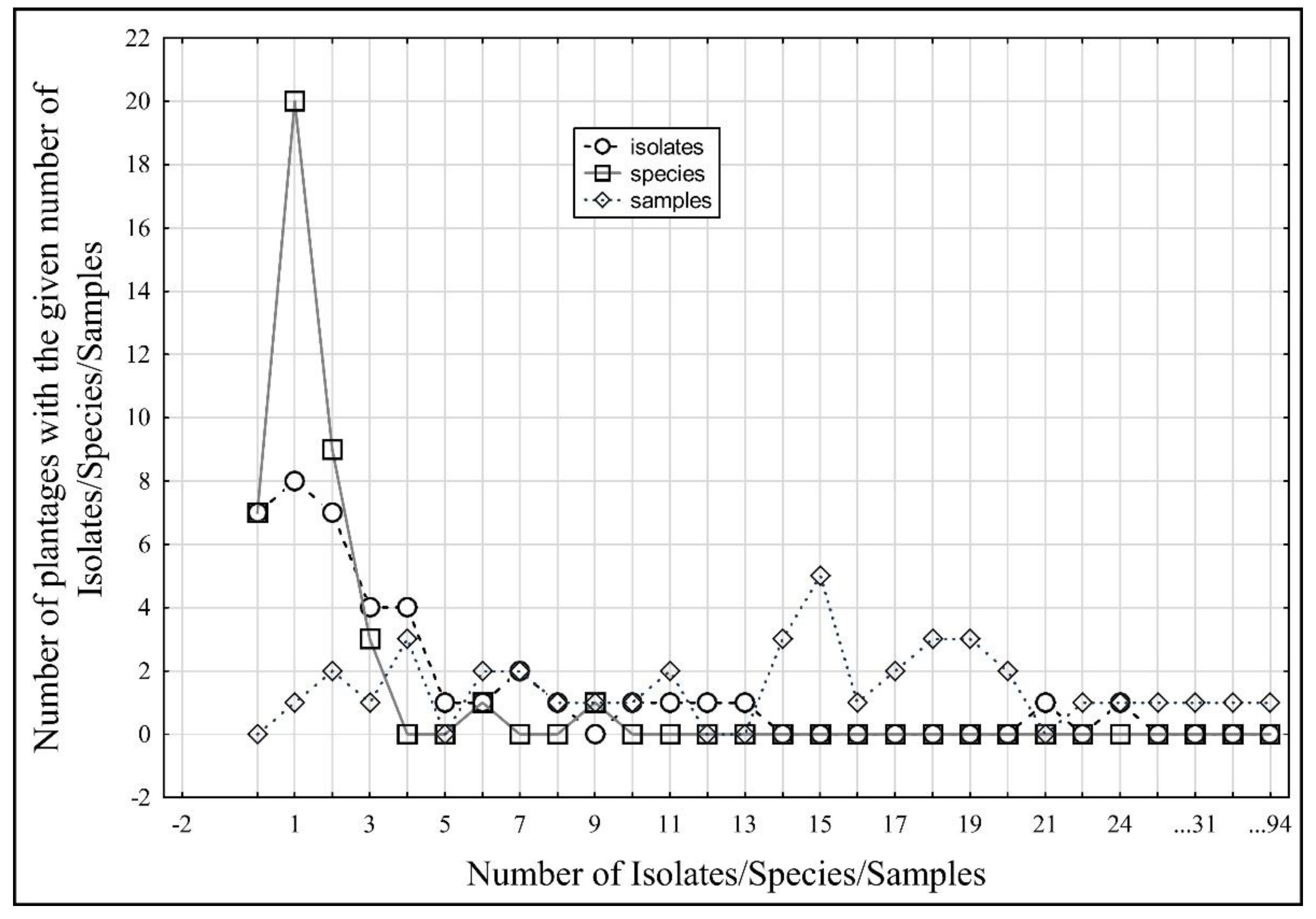
| No. | No. of Plantations with the Given Number of Sampled Plants | No. of Plantations with the Given Number of Isolates | No. of Plantations with the Given Number of Species |
|---|---|---|---|
| 0 | 0 | 7 | 7 |
| 1 | 1 | 8 | 20 |
| 2 | 2 | 7 | 9 |
| 3 | 1 | 4 | 3 |
| 4 | 3 | 4 | 0 |
| 5 | 0 | 1 | 0 |
| 6 | 2 | 1 | 1 |
| 7 | 2 | 2 | 0 |
| 8 | 1 | 1 | 0 |
| 9 | 1 | 0 | 1 |
| 10 | 1 | 1 | 0 |
| 11 | 2 | 1 | 0 |
| 12 | 0 | 1 | 0 |
| 13 | 0 | 1 | 0 |
| 14 | 3 | 0 | 0 |
| 15 | 5 | 0 | 0 |
| 16 | 1 | 0 | 0 |
| 17 | 2 | 0 | 0 |
| 18 | 3 | 0 | 0 |
| 19 | 3 | 0 | 0 |
| 20 | 2 | 0 | 0 |
| 21 | 0 | 1 | 0 |
| 22 | 1 | 0 | 0 |
| 23 | 0 | 0 | 0 |
| 24 | 1 | 1 | 0 |
| No. of Isolates | No. of Species Identified | No. of Plants Sampled | |
|---|---|---|---|
| No. of isolates | 1.000000 | 0.742487 | 0.485586 |
| No. of species identified | 0.742487 | 1.000000 | 0.191530 |
| No. of plants sampled | 0.485586 | 0.191530 | 1.000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pánek, M.; Maňasová, M.; Wenzlová, J.; Zouhar, M.; Mazáková, J. Peronosporales Species Associated with Strawberry Crown Rot in the Czech Republic. J. Fungi 2022, 8, 346. https://doi.org/10.3390/jof8040346
Pánek M, Maňasová M, Wenzlová J, Zouhar M, Mazáková J. Peronosporales Species Associated with Strawberry Crown Rot in the Czech Republic. Journal of Fungi. 2022; 8(4):346. https://doi.org/10.3390/jof8040346
Chicago/Turabian StylePánek, Matěj, Marie Maňasová, Jana Wenzlová, Miloslav Zouhar, and Jana Mazáková. 2022. "Peronosporales Species Associated with Strawberry Crown Rot in the Czech Republic" Journal of Fungi 8, no. 4: 346. https://doi.org/10.3390/jof8040346
APA StylePánek, M., Maňasová, M., Wenzlová, J., Zouhar, M., & Mazáková, J. (2022). Peronosporales Species Associated with Strawberry Crown Rot in the Czech Republic. Journal of Fungi, 8(4), 346. https://doi.org/10.3390/jof8040346