
Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates and Specimens
2.2. DNA Extraction, PCR Amplifications and Sequencing
2.3. Molecular Phylogenetic Analyses
2.3.1. Sequencing and Sequence Alignment
2.3.2. Phylogenetic Analyses
2.4. Morphological Observations
3. Results
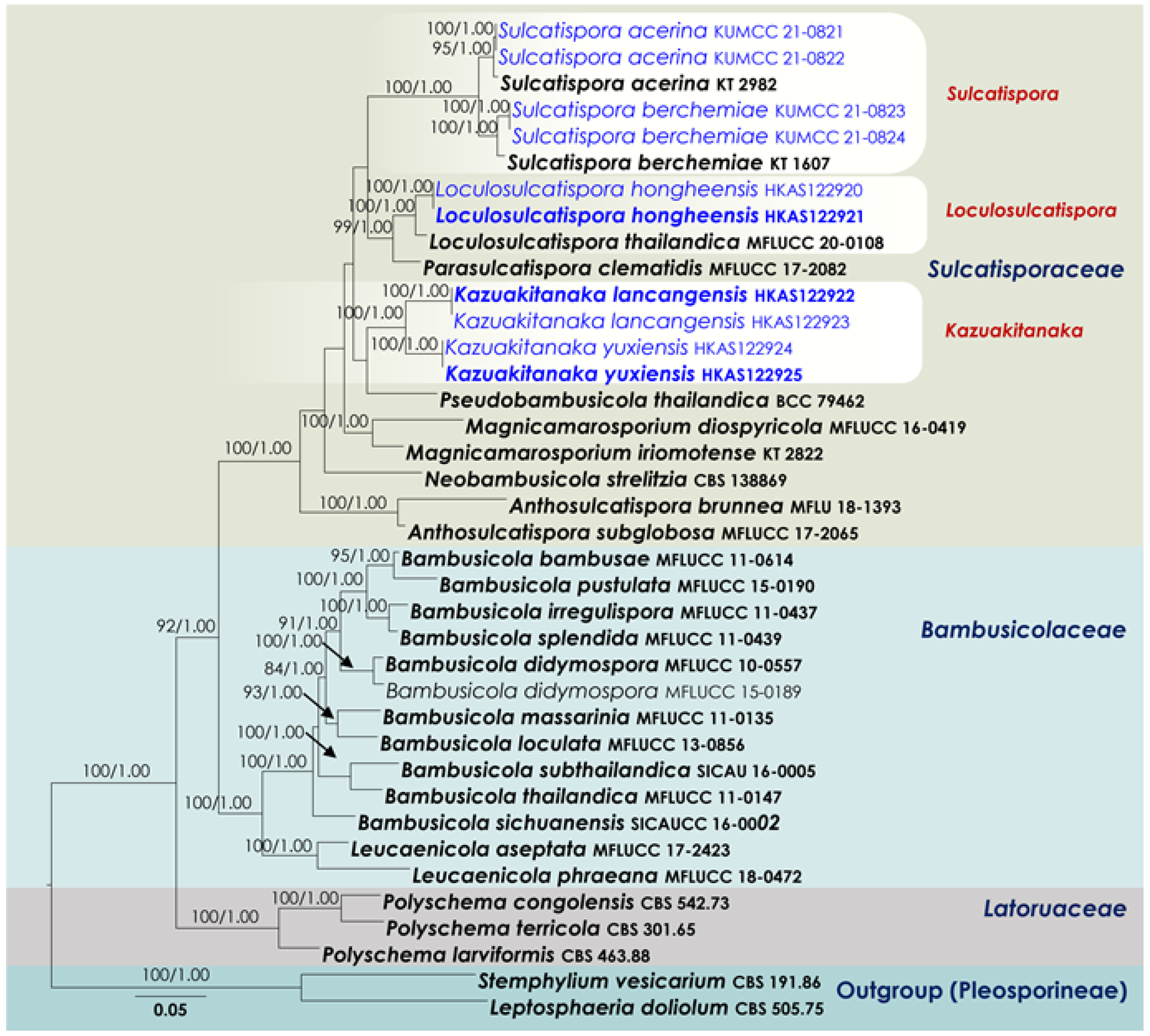
3.1. Phylogenetic Analyses
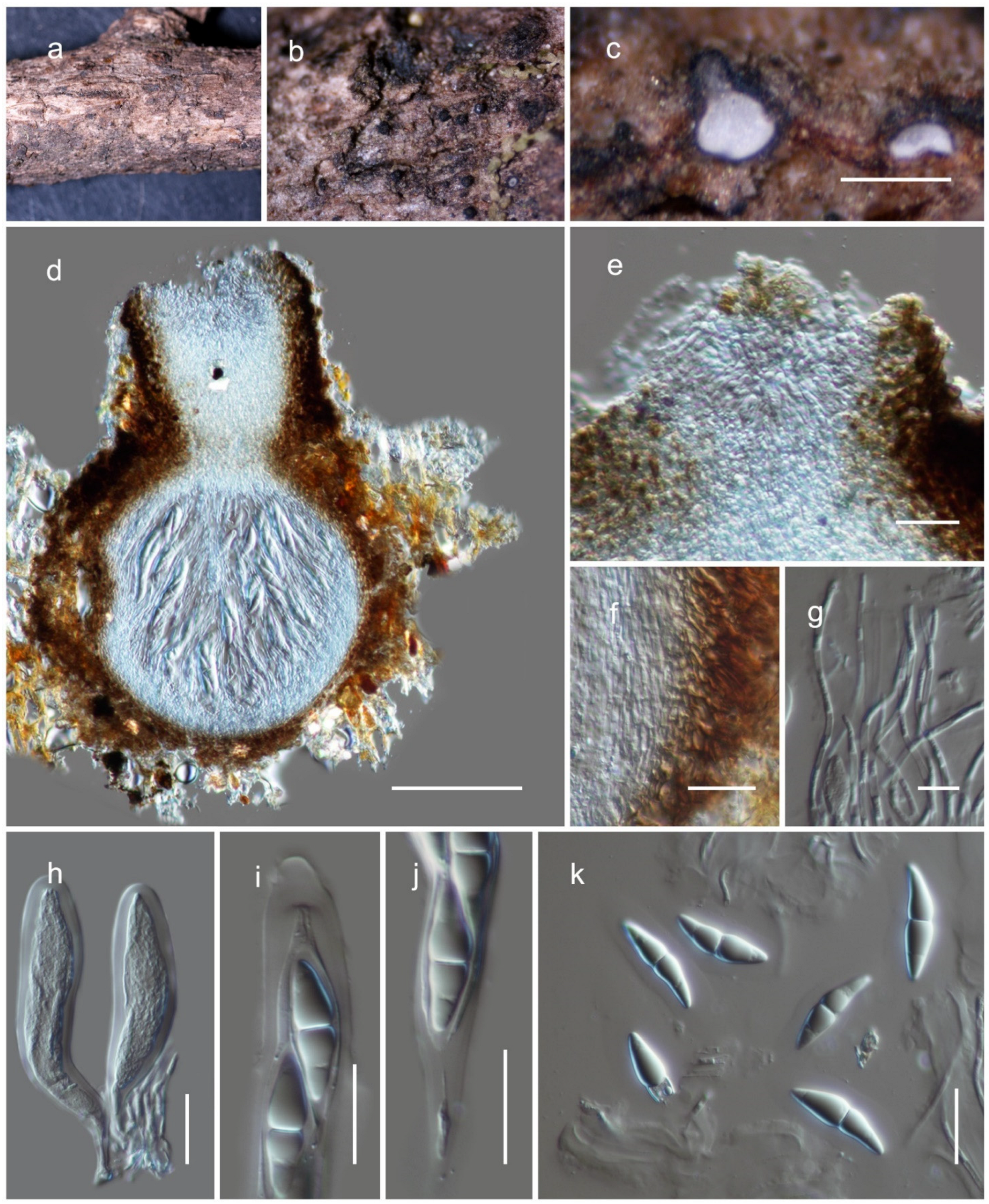
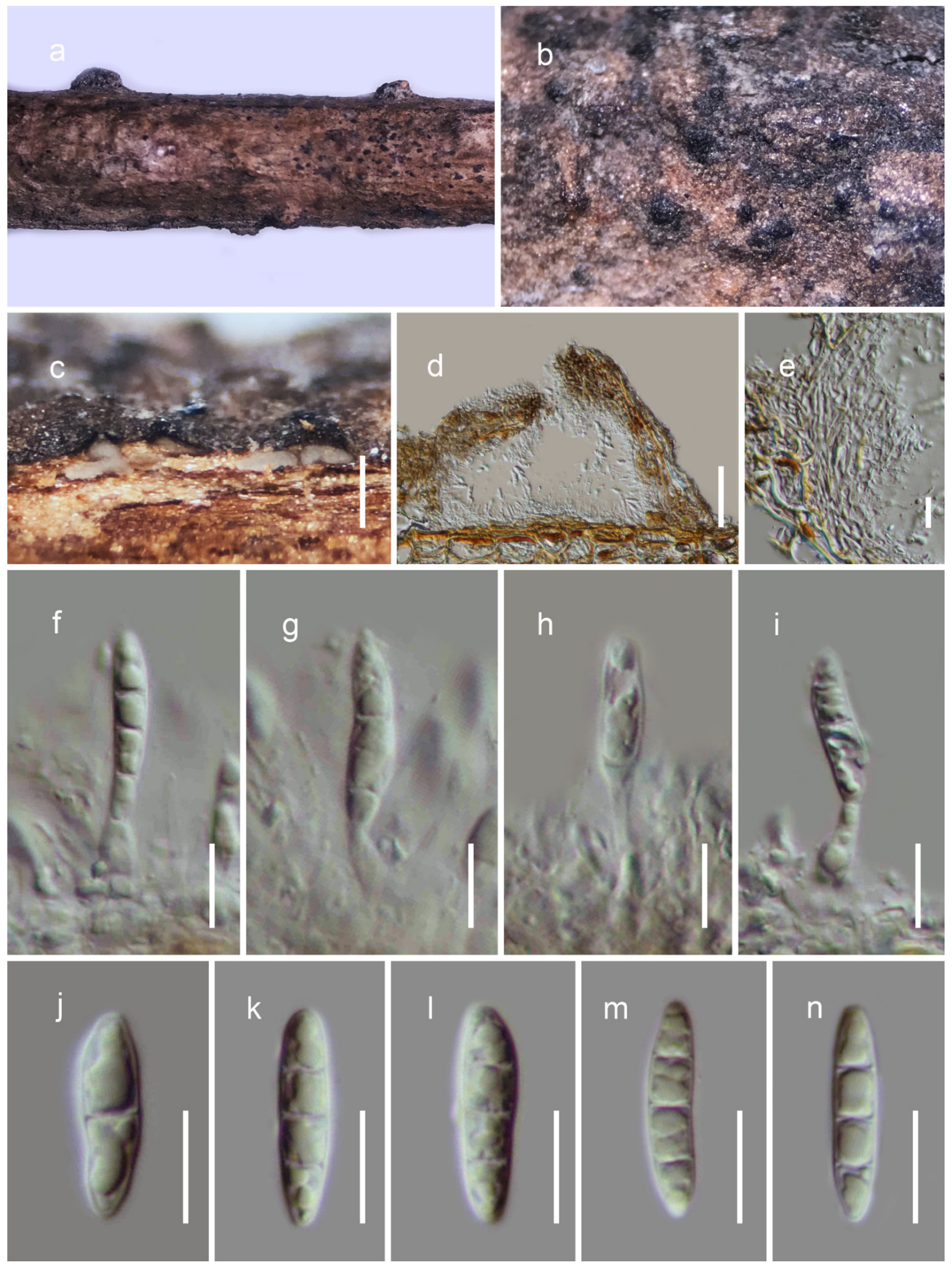
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bánki, O.; Roskov, Y.; Döring, M.; Ower, G.; Vandepitte, L.; Hobern, D.; Remsen, D.; Schalk, P.; DeWalt, R.E.; Keping, M.; et al. Catalogue of Life Checklist (Version 2022-01-14); Catalogue of Life: Leiden, NL, USA, 2022. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Bezerra, J. Editorial: Fungal Systematics and Biogeography. Front. Microbiol. 2022, 12, 827725. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef] [Green Version]
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Schmit, J.P.; Mueller, G.M. An estimate of the lower limit of global fungal diversity. Biodivers. Conserv. 2007, 16, 99–111. [Google Scholar] [CrossRef]
- Dighton, J. Fungi in Ecosystem Processes, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; 434p. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Jeewon, R.; Xu, J.C.; Lumyong, S.; Wanasinghe, D.N. Morpho-phylo taxonomy of novel dothideomycetous fungi associated with dead woody twigs in Yunnan Province, China. Front. Microbiol. 2021, 12, e654683. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa-2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Jiang, H.B.; Hyde, K.D.; Jayawardena, R.S.; Doilom, M.; Xu, J.; Phookamsak, R. Taxonomic and phylogenetic characterizations reveal two new species and two new records of Roussoella (Roussoellaceae, Pleosporales) from Yunnan, China. Mycol. Prog. 2019, 18, 577–591. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Hyde, K.D.; Wanasinghe, D.N.; Worthy, F.R.; Karunarathna, S.C. Addition to Melanommataceae: A new geographical record of Alpinaria rhododendri from Shangri La, China. Asian J. Mycol. 2020, 3, 335–344. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the systematics of microfungi colonizing dead woody twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, L.S.; Maharachchikumbura, S.S.N.; Mortimer, P.E.; Hyde, K.D.; Kang, J.C. Acrocordiella yunnanensis sp. nov. (Requienellaceae, Xylariales) from Yunnan, China. Phytotaxa 2021, 487, 103–113. [Google Scholar] [CrossRef]
- Ren, G.C.; Wanasinghe, D.N.; Monkai, J.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C.; Pang, A.; Gui, H. Novel saprobic Hermatomyces species (Hermatomycetaceae, Pleosporales) from China (Yunnan Province) and Thailand. MycoKeys 2021, 82, 57–79. [Google Scholar] [CrossRef]
- Yang, E.; Tibpromma, S.; Karunarathna, S.C.; Phookamsak, R.; Xu, J.; Zhao, Z.; Karunanayake, C.; Promputtha, I. Taxonomy and phylogeny of novel and extant taxa in Pleosporales associated with Mangifera indica from Yunnan, China (Series I). J. Fungi 2022, 8, 152. [Google Scholar] [CrossRef]
- Xie, N.; Phookamsak, R.; Jiang, H.; Zeng, Y.; Zhang, H.; Xu, F.; Lumyong, S.; Xu, J.; Hongsanan, S. Morpho-molecular characterization of five novel taxa in Parabambusicolaceae (Massarineae, Pleosporales) from Yunnan, China. J. Fungi 2022, 8, 108. [Google Scholar] [CrossRef]
- Su, H.; Hyde, K.D.; Maharachchikumbura, S.S.; Ariyawansa, H.A.; Luo, Z.; Promputtha, I.; Tian, Q.; Lin, C.; Shang, Q.; Zhao, Y. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Su, H.; Luo, Z.; Liu, X.; Su, X.; Hu, D.; Zhou, D.; Bahkali, A.H.; Hyde, K.D. Lentithecium cangshanense sp. nov. (Lentitheciaceae) from freshwater habitats in Yunnan Province, China. Phytotaxa 2016, 267, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Luo, Z.; Jeewon, R.; Bhat, D.J.; Bao, D.; Li, W.; Hao, Y.; Su, H.; Hyde, K.D. Morphology and multigene phylogeny reveal new genus and species of Torulaceae from freshwater habitats in northwestern Yunnan, China. Mycol. Prog. 2018, 17, 531–545. [Google Scholar] [CrossRef]
- Luo, Z.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Bao, D.; Zhao, Y.; Chai, H.; Su, H.; Su, X.; Hyde, K.D. Molecular phylogeny and morphological characterization of asexual fungi (Tubeufiaceae) from freshwater habitats in Yunnan, China. Cryptog. Mycol. 2017, 38, 27–53. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.Y. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Huang, S.; Maharachchikumbura, S.S.; Jeewon, R.; Bhat, D.J.; Chomnunti, P.; Hyde, K.D.; Lumyong, S. Morphological and molecular taxonomy of Jahnula dianchia sp. nov. (Jahnulales) from submerged wood in Dianchi Lake, Yunnan China. Mycol. Prog. 2018, 17, 547–555. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jeewon, R.; Gareth Jones, E.; Boonmee, S.; Kaewchai, S.; Manawasinghe, I.S.; Lumyong, S.; Hyde, K.D. Novel palmicolous taxa within Pleosporales: Multigene phylogeny and taxonomic circumscription. Mycol. Prog. 2018, 17, 571–590. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Bezerra, J.D.P.; Mortimer, P.E. (Eds.) Fungal Systematics and Biogeography; Frontiers Media SA: Lausanne, Switzerland, 2022; 446p. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Wanasinghe, D.N.; Wei, D.; Monkai, J.; Yasanthika, E.; Gui, H.; Mortimer, P.E.; Xu, J.; Hyde, K.D. Loculosulcatispora thailandica gen. et sp. nov. (Sulcatisporaceae), saprobic on woody litter in Thailand. Phytotaxa 2020, 475, 67–78. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Senwanna, C.; Cheewangkoon, R. Saprobic Dothideomycetes in Thailand: Phaeoseptum hydei sp. nov., a new terrestrial ascomycete in Phaeoseptaceae. Phytotaxa 2020, 449, 149–163. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nylander, J.A.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.0. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 14 March 2022).
- Phukhamsakda, C.; Jeewon, R.; McKenzie, E.H.C.; Xu, J.C. Morphology and phylogeny of Phaeoseptum mali sp. nov. (Phaeoseptaceae, Pleosporales) on bark of Malus halliana. Asian J. Mycol. 2019, 2, 118–128. [Google Scholar] [CrossRef]
- Liu, N.G.; Hyde, K.D.; Bhat, D.J.; Jumpathong, J.; Liu, J.K. Morphological and phylogenetic studies of Pleopunctum gen. nov. (Phaeoseptaceae, Pleosporales) from China. Mycosphere 2019, 10, 757–775. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; De Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Jones, E.B.G. Decaisnella formosa sp. nov. (Ascomycota, Massariaceae) from an Australian sandy beach. Can. J. Bot. 2003, 81, 598–600. [Google Scholar] [CrossRef]
- Mugambi, G.K.; Huhndorf, S.M. Molecular phylogenetics of Pleosporales: Melanommataceae and Lophiostomataceae recircumscribed (Plesporomycetidae, Dothideomycetes, Ascomycota). Stud. Mycol. 2009, 64, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fournier, J.; Phookamsak, R.; Bahkali, A.H.; Hyde, K.D. Halotthiaceae fam. nov. (Pleosporales) accommodates the new genus Phaeoseptum and several other aquatic genera. Mycologia 2013, 105, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Species Fungorum. Available online: http://www.speciesfungorum.org/Names/Names.asp (accessed on 8 March 2022).
- Koukol, O.; Delgado, G. Why morphology matters: The negative consequences of hasty descriptions of putative novelties in asexual ascomycetes. IMA Fungus 2021, 12, 26. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, E.B.G.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal planet description sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; Bhat, D.J.; Hongsanan, S.; Tibpromma, S.; Yang, J.B.; Promputtha, I. Magnicamarosporium diospyricola sp. nov. (Sulcatisporaceae) from Thailand. Mycosphere 2017, 8, 512–520. [Google Scholar] [CrossRef]
- Rupcic, Z.; Chepkirui, C.; Hernández-Restrepo, M.; Crous, P.W.; Luangsa-Ard, J.J.; Stadler, M. New nematicidal and antimicrobial secondary metabolites from a new species in the new genus, Pseudobambusicola thailandica. MycoKeys 2018, 33, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Wanasinghe, D.N.; Papizadeh, M.; Goonasekara, I.D.; Camporesi, E.; Bhat, D.J.; McKenzie, E.H.C.; Phillips, A.J.L.; Diederich, P.; et al. Taxonomy and phylogeny of dematiaceous coelomycetes. Fungal Divers. 2016, 77, 1–316. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain 1 | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| SSU | LSU | ITS | tef1 | rpb2 | ||
| Alfoldia vorosii | CBS 145501 T | MK589346 | MK589354 | JN859336 | MK599320 | - |
| Amorocoelophoma cassiae | MFLUCC 17-2283 T | NG_065775 | NG_066307 | NR_163330 | MK360041 | MK434894 |
| Angustimassarina acerina | MFLUCC 14-0505 T | NG_063573 | KP888637 | NR_138406 | KR075168 | - |
| Angustimassarina populi | MFLUCC 13-0034 T | NG_061204 | KP888642 | KP899137 | KR075164 | - |
| Angustimassarina quercicola | MFLUCC 14-0506 T | NG_063574 | KP888638 | KP899133 | KR075169 | - |
| Anthosulcatispora brunnea | MFLU 18-1393 T | - | MH644791 | MH644792 | - | - |
| Anthosulcatispora subglobosa | MFLUCC 17-2065 T | MT226705 | MT214592 | MT310636 | MT394649 | MT394706 |
| Bambusicola bambusae | MFLUCC 11-0614 T | JX442039 | JX442035 | JX442031 | KP761722 | KP761718 |
| Bambusicola didymospora | MFLUCC 10-0557 T | KU872110 | KU863105 | KU940116 | KU940188 | KU940163 |
| Bambusicola didymospora | MFLUCC 15-0189 | KU872111 | KU863106 | KU940117 | KU940189 | KU940164 |
| Bambusicola irregulispora | MFLUCC 11-0437 T | JX442040 | JX442036 | JX442032 | KP761723 | KP761719 |
| Bambusicola loculata | MFLUCC 13-0856 T | KP761735 | KP761729 | KP761732 | KP761724 | KP761715 |
| Bambusicola massarinia | MFLUCC 11-0135 T | KU872115 | KU863111 | KU940122 | KU940192 | KU940169 |
| Bambusicola pustulata | MFLUCC 15-0190 T | KU872112 | KU863107 | KU940118 | KU940190 | KU940165 |
| Bambusicola sichuanensis | SICAUCC 16-0002 T | MK253528 | MK253532 | MK253473 | MK262828 | MK262830 |
| Bambusicola splendida | MFLUCC 11-0439 T | JX442042 | JX442038 | JX442034 | KP761726 | KP761717 |
| Bambusicola subthailandica | SICAU 16-0005 T | MK253529 | MK253533 | MK253474 | MK262829 | MK262831 |
| Bambusicola thailandica | MFLUCC 11-0147 T | KU872113 | KU863108 | KU940119 | KU940191 | KU940166 |
| Crassiclypeus aquaticus | CBS 143643 T | LC312472 | LC312530 | LC312501 | LC312559 | LC312588 |
| Decaisnella formosa | BCC 25616 | GQ925833 | GQ925846 | - | GU479851 | - |
| Decaisnella formosa | BCC 25617 | GQ925834 | GQ925847 | - | GU479850 | - |
| Flabellascoma cycadicola | CBS 143644 T | LC312473 | LC312531 | LC312502 | LC312560 | LC312589 |
| Forliomyces uniseptata | MFLUCC 15-0765 T | NG_061234 | NG_059659 | NR_154006 | KU727897 | - |
| Gloniopsis praelonga | CBS 112415 | FJ161134 | FJ161173 | - | FJ161090 | FJ161113 |
| Guttulispora crataegi | MFLUCC 13-0442 T | KP899125 | KP888639 | KP899134 | KR075161 | - |
| Halotthia posidoniae | BBH 22481 | GU479752 | GU479786 | - | - | - |
| Hysterium angustatum | MFLUCC 16-0623 | GU397359 | FJ161180 | - | FJ161096 | MH535875 |
| Kazuakitanaka lancangensis | HKAS122922 T | ON009088 | ON009104 | ON009120 | ON009263 | - |
| Kazuakitanaka lancangensis | HKAS122923 | ON009089 | ON009105 | ON009121 | ON009264 | - |
| Kazuakitanaka yuxiensis | HKAS122924 | ON009092 | ON009108 | ON009124 | ON009267 | ON009290 |
| Kazuakitanaka yuxiensis | HKAS122925 T | ON009093 | ON009109 | ON009125 | ON009268 | ON009291 |
| Leptoparies palmarum | CBS 143653 T | LC312485 | LC312543 | LC312514 | LC312572 | LC312601 |
| Leptosphaeria doliolum | CBS 505.75 T | GU296159 | GU301827 | NR_155309 | GU349069 | KY064035 |
| Leucaenicola aseptata | MFLUCC 17-2423 T | NG_065776 | MK347963 | NR_163332 | MK360059 | MK434891 |
| Leucaenicola phraeana | MFLUCC 18-0472 T | NG_065784 | MK348003 | MK347785 | MK360060 | MK434867 |
| Lignosphaeria fusispora | MFLUCC 11-0377 T | - | KP888646 | NR_164233 | - | - |
| Loculosulcatispora thailandica | MFLUCC 20-0108 T | MT383968 | MT383964 | MT376742 | MT380476 | MT380475 |
| Loculosulcatispora hongheensis | HKAS122920 | ON009090 | ON009106 | ON009122 | ON009265 | ON009288 |
| Loculosulcatispora hongheensis | HKAS122921 T | ON009091 | ON009107 | ON009123 | ON009266 | ON009289 |
| Magnicamarosporium diospyricola | MFLUCC 16-0419 T | NG_065102 | KY554212 | NR_153486 | KY554209 | KY554208 |
| Magnicamarosporium iriomotense | KT 2822 T | AB797219 | AB807509 | AB809640 | AB808485 | - |
| Mauritiana rhizophorae | BCC 28866 | GU371832 | GU371824 | - | GU371817 | - |
| Misturatosphaeria aurantiacinotata | GKM 1238 T | - | NG_059927 | - | GU327761 | - |
| Neobambusicola strelitzia | CBS 138869 T | - | KP004495 | NR_137945 | MG976037 | - |
| Parasulcatispora clematidis | MFLUCC 17-2082 T | - | MT214593 | MT310637 | MT394650 | - |
| Phaeoseptum aquaticum | CBS 123113 T | - | JN644072 | KY940803 | - | - |
| Phaeoseptum carolshearerianum | NFCCI-4221 T | MK307816 | MK307813 | MK307810 | MK309874 | MK309877 |
| Phaeoseptum carolshearerianum | NFCCI-4384 | MK307818 | MK307815 | MK307812 | MK309876 | MK309879 |
| Phaeoseptum hydei | MFLUCC 17-0801 T | MT240624 | MT240623 | MT240622 | MT241506 | - |
| Phaeoseptum mali | HKAS122916 | ON009082 | ON009098 | ON009114 | ON009257 | ON009282 |
| Phaeoseptum mali | HKAS122917 | ON009083 | ON009099 | ON009115 | ON009258 | ON009283 |
| Phaeoseptum mali | MFLUCC 17-2108 T | - | MK625197 | MK659580 | MK647990 | - |
| Phaeoseptum manglicola | NFCCI-4666 T | MK307817 | MK307814 | MK307811 | MK309875 | MK309878 |
| Phaeoseptum terricola | MFLUCC 10-0102 T | MH105780 | MH105779 | MH105778 | MH105781 | - |
| Pleopunctum ellipsoideum | MFLUCC 19-0390 T | MK804514 | MK804517 | MK804512 | MK828510 | - |
| Pleopunctum pseudoellipsoideum | KUMCC 21-0820 | ON009084 | ON009100 | ON009116 | ON009259 | ON009284 |
| Pleopunctum pseudoellipsoideum | HKAS122915 | ON009085 | ON009101 | ON009117 | ON009260 | ON009285 |
| Pleopunctum pseudoellipsoideum | MFLUCC 19-0391 T | - | MK804518 | MK804513 | MK828511 | - |
| Pleopunctum thailandicum | MFLUCC 21-0039 T | - | MZ198896 | MZ198894 | MZ172461 | - |
| Pleopunctum menglaense | KUMCC 21-0025 | ON009086 | ON009102 | ON009118 | ON009261 | ON009286 |
| Pleopunctum menglaense | KUMCC 21-0026 T | ON009087 | ON009103 | ON009119 | ON009262 | ON009287 |
| Polyschema congolensis | CBS 542.73 T | - | EF204502 | MH860770 | - | EF204486 |
| Polyschema larviformis | CBS 463.88 T | - | EF204503 | - | - | - |
| Polyschema terricola | CBS 301.65 T | EF204519 | EF204504 | NR_160100 | - | EF204487 |
| Pseudoaurantiascoma kenyense | GKM 1195 T | - | NG_059928 | - | GU327767 | - |
| Pseudobambusicola thailandica | BCC 79462 | - | MG926560 | MG926559 | MG926562 | MG926561 |
| Pseudolophiostoma cornisporum | CBS 143654 T | LC312486 | LC312544 | LC312515 | LC312573 | LC312602 |
| Ramusculicola thailandica | MFLUCC 13-0284 T | KP899131 | KP888647 | KP899141 | KR075167 | - |
| Sporormurispora atraphaxidis | MFLUCC 17-0742 T | NG_061296 | NG_059880 | NR_157546 | - | - |
| Stemphylium vesicarium | CBS 191.86 T | DQ247812 | DQ247804 | MH861935 | DQ471090 | KC584471 |
| Sulcatispora acerina | KUMCC 21-0821 | ON009096 | ON009112 | ON009128 | ON009271 | ON009294 |
| Sulcatispora acerina | KUMCC 21-0822 | ON009097 | ON009113 | ON009129 | ON009272 | ON009295 |
| Sulcatispora acerina | KT 2982 T | LC014605 | LC014610 | LC014597 | LC014615 | - |
| Sulcatispora berchemiae | KT 1607 T | AB797244 | AB807534 | AB809635 | AB808509 | - |
| Sulcatispora berchemiae | KUMCC 21-0823 | ON009094 | ON009110 | ON009126 | ON009269 | ON009292 |
| Sulcatispora berchemiae | KUMCC 21-0824 | ON009095 | ON009111 | ON009127 | ON009270 | ON009293 |
| Sulcosporium thailandicum | MFLUCC 12-0004 T | KT426564 | KT426563 | MG520958 | - | - |
| Teichospora melanommoides | CBS 140733 T | - | KU601585 | NR_154632 | KU601610 | - |
| Teichospora pusilla | CBS 140731 T | - | KU601586 | NR_154633 | KU601605 | - |
| Teichospora rubriostiolata | CBS 140734 T | - | KU601590 | NR_154634 | KU601609 | KU601599 |
| Thyridaria macrostomoides | GKM 1033 | - | GU385190 | - | GU327776 | - |
| Thyridaria macrostomoides | GKM 1159 | - | GU385185 | - | GU327778 | - |
| Thyridaria macrostomoides | GKM 224N | - | GU385191 | - | GU327777 | - |
| Vaginatispora appendiculata | MFLUCC 16-0314 T | KU743219 | KU743218 | KU743217 | KU743220 | - |
| Westerdykella ornata | CBS 379.55 | GU296208 | GU301880 | AY943045 | GU349021 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanasinghe, D.N.; Ren, G.-C.; Xu, J.-C.; Cheewangkoon, R.; Mortimer, P.E. Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China. J. Fungi 2022, 8, 375. https://doi.org/10.3390/jof8040375
Wanasinghe DN, Ren G-C, Xu J-C, Cheewangkoon R, Mortimer PE. Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China. Journal of Fungi. 2022; 8(4):375. https://doi.org/10.3390/jof8040375
Chicago/Turabian StyleWanasinghe, Dhanushka N., Guang-Cong Ren, Jian-Chu Xu, Ratchadawan Cheewangkoon, and Peter E. Mortimer. 2022. "Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China" Journal of Fungi 8, no. 4: 375. https://doi.org/10.3390/jof8040375
APA StyleWanasinghe, D. N., Ren, G. -C., Xu, J. -C., Cheewangkoon, R., & Mortimer, P. E. (2022). Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China. Journal of Fungi, 8(4), 375. https://doi.org/10.3390/jof8040375