
Two New Species of Sidera (Hymenochaetales, Basidiomycota) from Southwest China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
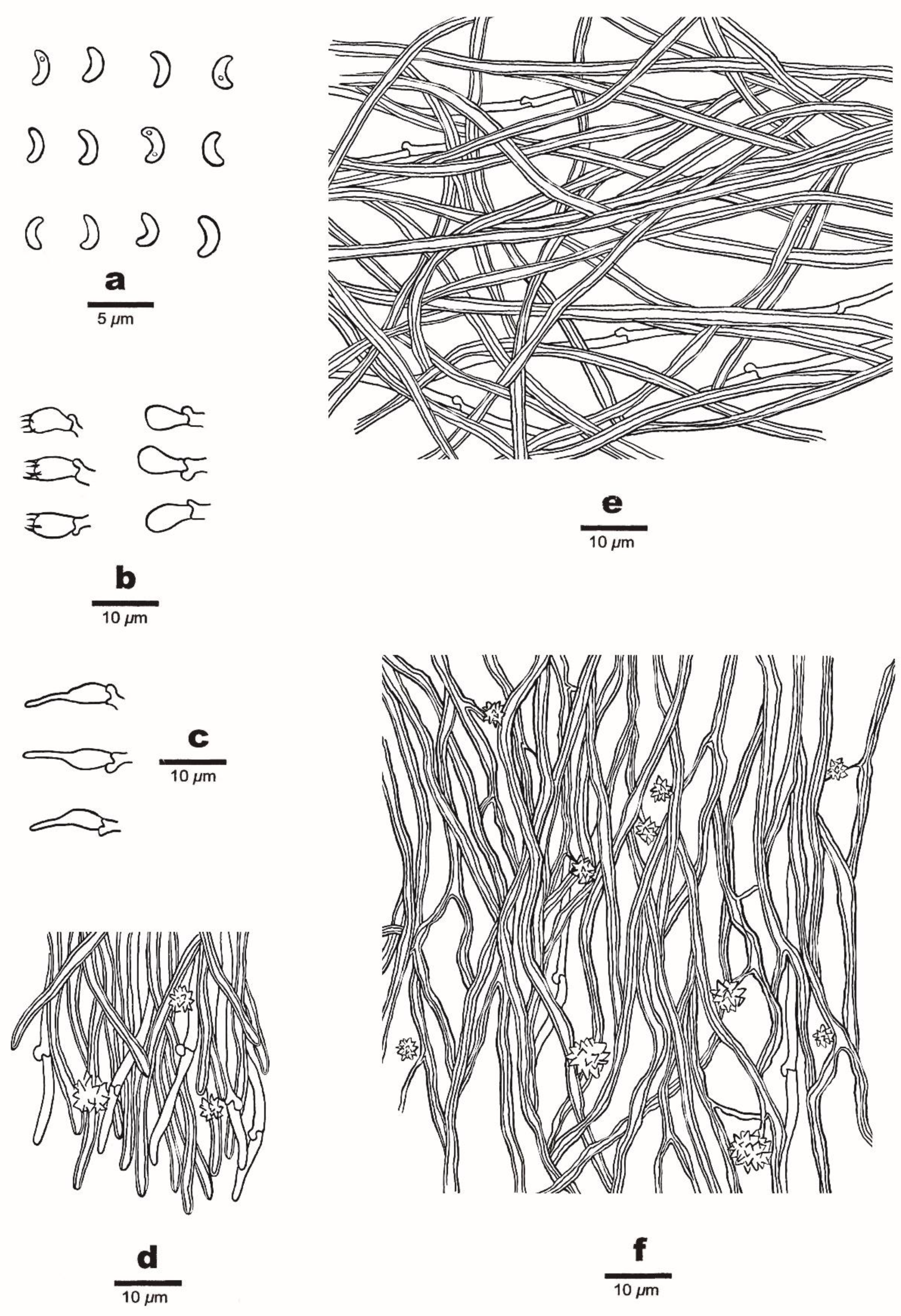
2.2. Morphological Studies
2.3. DNA Extraction, PCR, and Sequencing
2.4. Phylogenetic Analyses
3. Results
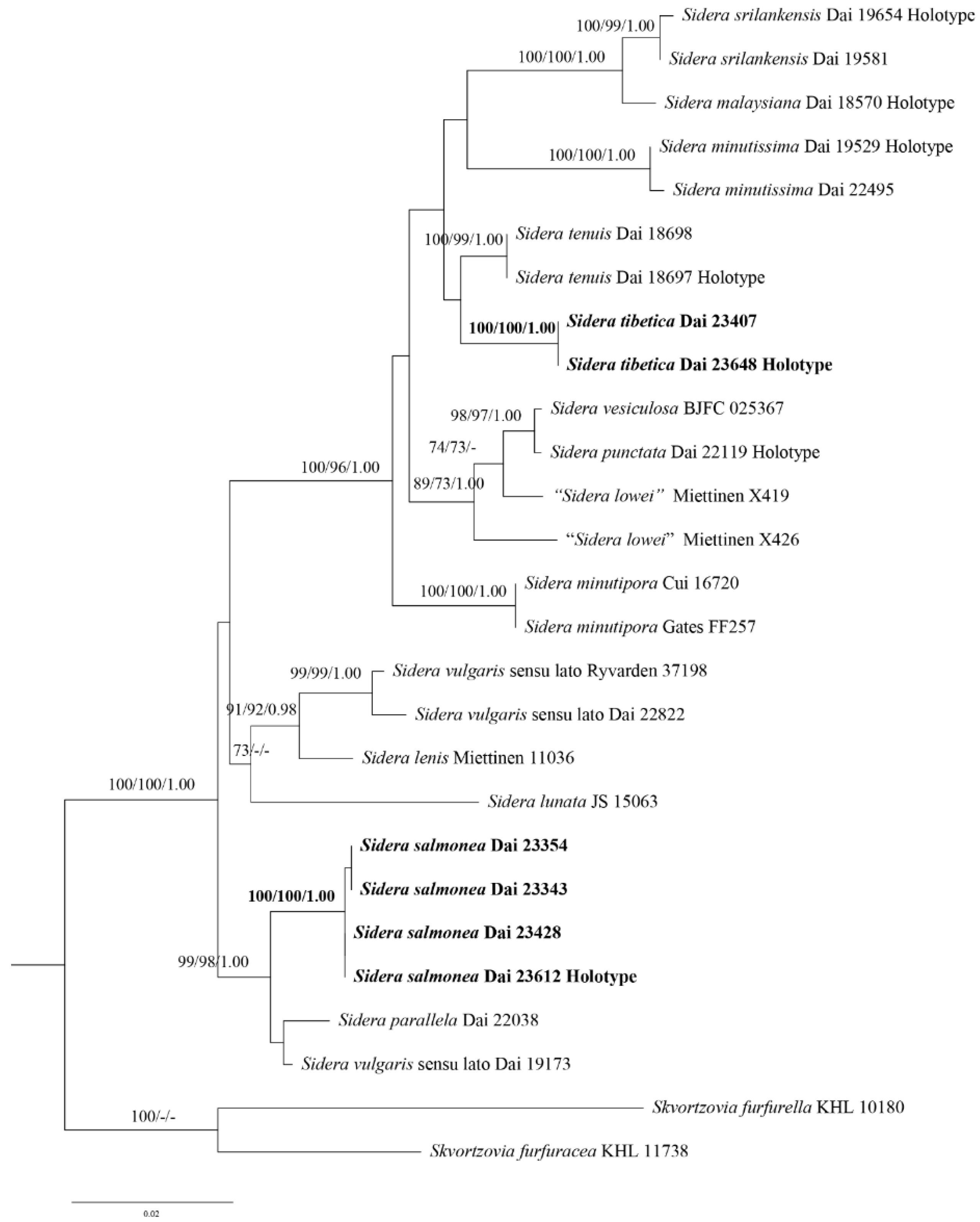
3.1. Phylogenetic Analyses
3.2. Taxonomy
- 1.
- 2.
| 1. Hymenium grandinioid to odontioid | S. lunata |
| 1. Hymenium poroid | 2 |
| 2. Hyphal system monomitic | 3 |
| 2. Hyphal system dimitic | 6 |
| 3. Basidiospores mostly < 1 μm in width | 4 |
| 3. Basidiospores mostly > 1 μm in width | 5 |
| 4. Pores 7–9 per mm; basidiospores 2.9–3.7 μm long | S. vesiculosa |
| 4. Pores 6–7 per mm; basidiospores 3.9–4.5 μm long | S. roseo-bubalina |
| 5. Pores 6–8 per mm; cystidioles present, some branched | S. lowei |
| 5. Pores 8–9 per mm; cystidioles absent | S.punctata |
| 6. Basidiospores > 1.5 μm in width | S. lenis |
| 6. Basidiospores < 1.5 μm in width | 7 |
| 7. Skeletal hyphae becoming swollen in KOH | 8 |
| 7. Skeletal hyphae almost unchanged in KOH | 10 |
| 8. Pores 5–7 per mm; basidiospores 3.7–4.3 μm long | S. minutipora |
| 8. Pores 9–11 per mm; basidiospores 2.9–3.3 μm long | 9 |
| 9. Basidiospores allantoid, skeletal hyphae distinctly swollen in KOH | S. inflata |
| 9. Basidiospores lunate, skeletal hyphae slightly swollen in KOH | S. malaysiana |
| 10. Tramal hyphae parallel along the tubes | S. parallela |
| 10. Tramal hyphae interwoven | 11 |
| 11. Generative hyphae at dissepiments even | 12 |
| 11. Generative hyphae at dissepiments with swollen tips | 14 |
| 12. Basidiospores > 3.5 μm long | S. srilankensis |
| 12. Basidiospores < 3.5 μm long | 13 |
| 13. Sterile margin distinct, white; basidiospore length/width > 3 | S. salmonea |
| 13. Sterile margin indistinct to almost absent; basidiospore length/width < 3 | S. tibetica |
| 14. Basidiospores < 3.6 μm long | S. vulgaris |
| 14. Basidiospores > 3.8 μm long | 15 |
| 15. Sterile margin distinct, fimbriate; basidiospore length/width < 4 | S. minutissima |
| 15. Sterile margin indistinct to almost absent; basidiospore length/width > 4 | S. tenuis |
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miettinen, O.; Larsson, K.H. Sidera. A new genus in Hymenochaetales with poroid and hydnoid species. Mycol. Prog. 2011, 10, 131–141. [Google Scholar] [CrossRef]
- Liu, Z.B.; Zhou, M.; Yuan, Y.; Dai, Y.C. Global diversity and taxonomy of Sidera (Hymenochaetales, Basidiomycota): Four new species and keys to species of the genus. J. Fungi 2021, 7, 251. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.C. Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 2010, 45, 131–343. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Italy, 1996; pp. 1–6. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2018; Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 15 January 2021).
- Li, H.J.; Cui, B.K.; Dai, Y.C. Taxonomy and multi-gene phylogeny of Datronia (Polyporales, Basidiomycota). Persoonia 2014, 32, 170–182. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gefand, D.H., Sninsky, J.J., White, J.T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, B.K.; Song, J.; Dai, Y.C. Fragiliporiaceae, a new family of Polyporales (Basidiomycota). Fungal Divers. 2015, 70, 115–126. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. Bioedit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. Available online: https://www.mesquiteproject.org/ (accessed on 7 March 2022).
- Chen, J.J.; Cui, B.K. Phlebiporia bubalina gen. et. sp. nov. (Meruliaceae, Polyporales) from Southwest China with a preliminary phylogeny based on rDNA sequences. Mycol. Prog. 2014, 13, 563–573. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analyses and post analyses of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. Available online: http://www.phylo.org/sub_sections/portal (accessed on 4 August 2009).
- Ronquist, F.; Teslenko, M.; Mark, P.; Avres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes3.2: Efficient Bayesian phylogenetic inference and model choice, across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A. Molecular Evolution, Phylogenetics and Epidemiology. FigTree ver. 1.4.4 Software. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 7 March 2022).
- Du, R.; Wu, F.; Gate, G.M.; Dai, Y.C.; Tian, X.M. Taxonomy and phylogeny of Sidera (Hymenochaetales, Basidiomycota): Four new species and keys to species of the genus. MycoKeys 2020, 68, 115–135. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.C.; Yu, C.J.; Wang, H.C. Polypores from eastern Xizang (Tibet), western China. Ann. Bot. Fenn. 2007, 44, 135–145. [Google Scholar]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar]
- Yuan, H.S.; Dai, Y.C. Polypores from northern and central Yunnan Province, Southwestern China. Sydowia 2008, 60, 147–159. [Google Scholar]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, J.L.; Cui, B.K. Two new species of Fomitiporia (Hymenochaetales, Basidiomycota) from Tibet, southwest China. Mycologia 2016, 108, 1010–1017. [Google Scholar] [CrossRef]
- Zhou, L.W.; Vlasák, J.; Qin, W.M.; Dai, Y.C. Global diversity and phylogeny of the Phellinus igniarius complex (Hymenochaetales, Basidiomycota) with the description of five new species. Mycologia 2016, 108, 192–204. [Google Scholar] [CrossRef]
- Chen, J.J.; Cui, B.K.; Zhou, L.W.; Korhonen, K.; Dai, Y.C. Phylogeny, divergence time estimation, and biogeography of the genus Heterobasidion (Basidiomycota, Russulales). Fungal Divers. 2015, 71, 185–200. [Google Scholar] [CrossRef]
- Tian, X.M.; Yu, H.Y.; Zhou, L.W.; Decock, C.; Vlasák, J.; Dai, Y.C. Phylogeny and taxonomy of the Inonotus linteus complex. Fungal Divers. 2013, 58, 159–169. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Growing Habit | Hymenophore | Hyphal System | Cystidioles | Skeletal Hyphae in KOH | Spores Shape | Spore Size (µm) |
|---|---|---|---|---|---|---|---|
| S. inflata | Annual | Poroid, 9–10/mm | Dimitic | Present | Swollen | Allantoid | 3–3.3 × 0.9–1.1 |
| S. lenis | Perennial | Poroid, 4–6/mm | Dimitic | Present | Swollen | Allantoid to lunate | 3.9–4.9 × 1.5–2 |
| S. lowei | Annual | Poroid, 6–8/mm | Monomitic | Present, some branched | – | Allantoid | 3.5–5 × 1–1.2 |
| S. lunata | Annual | Hydnoid, 8–9/mm | Monomitic | Present | – | Allantoid | 2.5–3.8 × 1.6–1.9 |
| S. malaysiana | Annual | Poroid, 9–11/mm | Dimitic | Present | Swollen | Lunate | 2.9–3.2 × 1–1.2 |
| S. minutipora | Annual | Poroid, 5–7/mm | Dimitic | Present | Swollen | Allantoid | 3.7–4.3 × 1–1.3 |
| S. minutissima | Annual | Poroid, 7–9/mm | Dimitic | Present | Almost unchanged | Allantoid | 3.8–4.4 × 0.9–1.3 |
| S. parallela | Annual | Poroid, 6–8/mm | Dimitic | Present | Almost unchanged | Lunate | 2.8–3.3 × 0.9–1.2 |
| S. punctata | Annual | Poroid, 8–9/mm | Monomitic | Absent | – | Allantoid to lunate | 3.8–4.8 × 1–1.3 |
| S. roseo-bubalina | Annual | Poroid, 6–7/mm | Monomitic | Present | – | Lunate | 3.9–4.5 × 0.8–1 |
| S. salmonea | Annual | Poroid, 7–9/mm | Dimitic | Present | Almost unchanged | Lunate | 3–3.5 × 0.9–1.1 |
| S. srilankensis | Annual | Poroid, 6–8/mm | Dimitic | Present | Almost unchanged | Lunate | 3.5–4 × 1–1.3 |
| S. tibetica | Annual | Poroid, 7–8/mm | Dimitic | Present | Almost unchanged | Lunate | 2.9–3.1 × 1–1.1 |
| S. tenuis | Annual | Poroid, 8–10/mm | Dimitic | Present | Almost unchanged | Allantoid | 4.2–5 × 0.8–1 |
| S. vesiculosa | Annual | Poroid, 7–9/mm | Monomitic | Present | – | Allantoid to lunate | 2.9–3.7 × 0.6–1 |
| S. vulgaris | Perennial | Poroid, 6–8/mm | Dimitic | Present, some branched | Almost unchanged | Allantoid to lunate | 2.9–3.6 × 0.9–1.4 |
| Species | Specimen No. | Locality | GenBank Accession No. | |
|---|---|---|---|---|
| ITS | nLSU | |||
| S. lenis | Miettinen 11036 | Finland | FN907914 | FN907914 |
| “S. lowei” | Miettinen X419 | Venezuela | FN907917 | FN907917 |
| “S. lowei” | Miettinen X426 | New Zealand | FN907919 | FN907919 |
| S. lunata | JS 15063 | Norway | DQ873593 | DQ873593 |
| S. malaysiana | Dai 18570 | Malaysia | MW198481 | MW192007 |
| S. minutipora | Gates FF257 | Australia | FN907922 | FN907922 |
| S. minutipora | Cui 16720 | Australia | MN621349 | MN621348 |
| S. minutissima | Dai 19529 | Sri Lanka | MN621352 | MN621350 |
| S. minutissima | Dai 22495 | China | OM974248 * | OM974240 * |
| S. parallela | Dai 22038 | China | MW477793 * | MW474964 * |
| S. punctata | Dai 22119 | China | MW418438 | MW418437 |
| S. salmonea | Dai 23343 | China | OM974249 * | OM974241 * |
| S. salmonea | Dai 23354 | China | OM974250 * | OM974242 * |
| S. salmonea | Dai 23428 | China | OM974251 * | OM974243 * |
| S. salmonea | Dai 23612 | China | – | OM974247 * |
| S. srilankensis | Dai 19581 | Sri Lanka | MN621345 | MN621347 |
| S. srilankensis | Dai 19654 | Sri Lanka | MN621344 | MN621346 |
| S. tenuis | Dai 18697 | Australia | MK331865 | MK331867 |
| S. tenuis | Dai 18698 | Australia | MK331866 | MK331868 |
| S. tibetica | Dai 23407 | China | OM974252 * | OM974244 * |
| S. tibetica | Dai 23648 | China | OM974253 * | OM974245 * |
| S. vesiculosa | BJFC025367 | Singapore | MH636565 | MH636567 |
| S. vulgaris sensu lato | Ryvarden 37198 | New Zealand | FN907918 | FN907918 |
| S. vulgaris sensu lato | Dai 19173 | Canada | MW198477 * | MW192005 * |
| S. vulgaris sensu lato | Dai 22822 | China | OM974254 * | OM974246 * |
| Skvortzovia furfuracea | KHL 11738 | Finland | DQ873648 | DQ873648 |
| S. furfurella | KHL 10180 | Puerto Rico | DQ873649 | DQ873649 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-B.; Zhou, M.; Wu, F.; Yu, J. Two New Species of Sidera (Hymenochaetales, Basidiomycota) from Southwest China. J. Fungi 2022, 8, 385. https://doi.org/10.3390/jof8040385
Liu Z-B, Zhou M, Wu F, Yu J. Two New Species of Sidera (Hymenochaetales, Basidiomycota) from Southwest China. Journal of Fungi. 2022; 8(4):385. https://doi.org/10.3390/jof8040385
Chicago/Turabian StyleLiu, Zhan-Bo, Meng Zhou, Fang Wu, and Jian Yu. 2022. "Two New Species of Sidera (Hymenochaetales, Basidiomycota) from Southwest China" Journal of Fungi 8, no. 4: 385. https://doi.org/10.3390/jof8040385
APA StyleLiu, Z. -B., Zhou, M., Wu, F., & Yu, J. (2022). Two New Species of Sidera (Hymenochaetales, Basidiomycota) from Southwest China. Journal of Fungi, 8(4), 385. https://doi.org/10.3390/jof8040385