
Morpho-Molecular Evidence Reveals Four Novel Species of Gymnopus (Agaricales, Omphalotaceae) from China

 , ,
, , 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Abbreviations for Specific Names and Additional Mycological Glossary
2.2. Specimen
2.3. Morphological Studies
2.4. DNA Extraction, Amplification and Sequencing
2.5. Phylogenetic Analyses
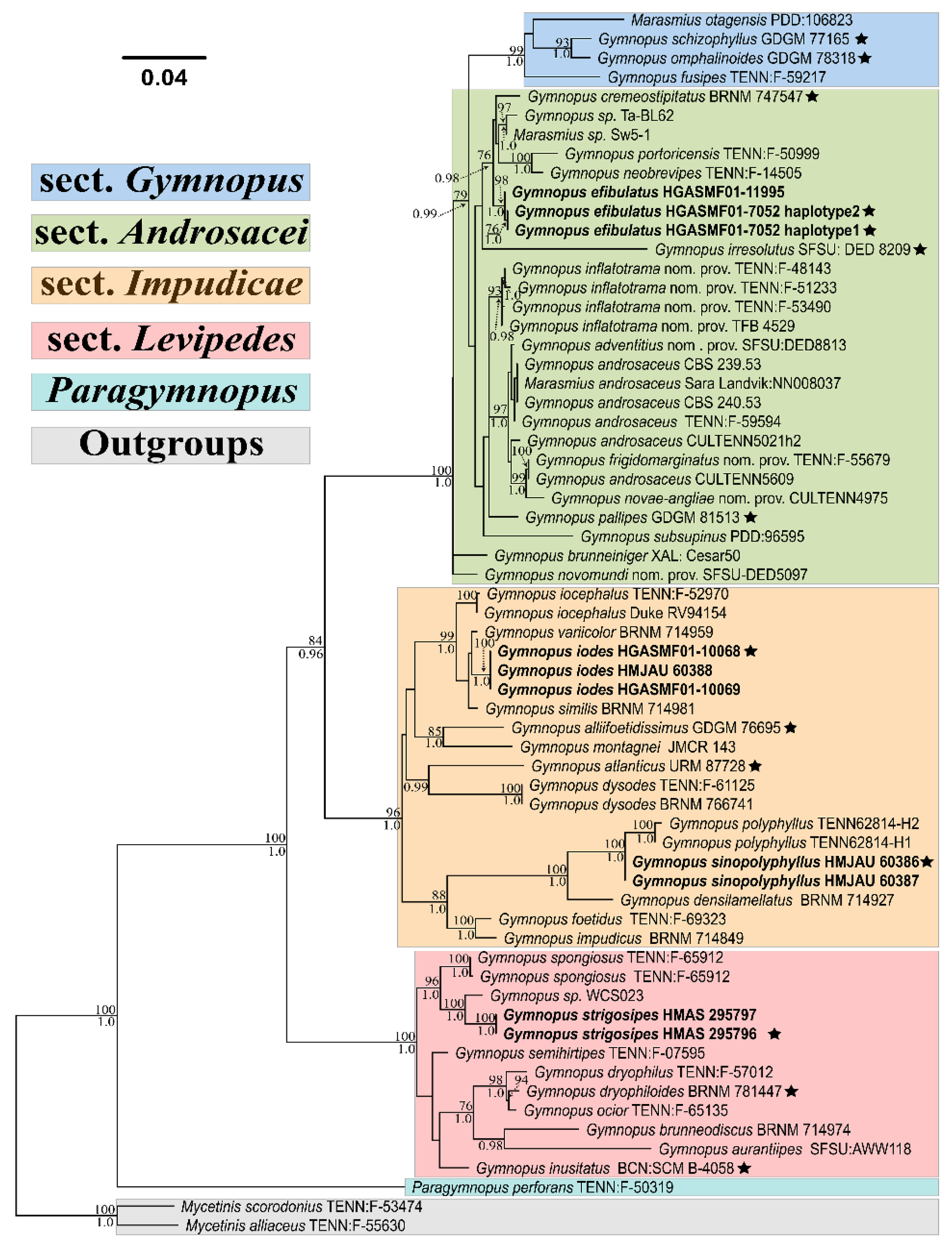
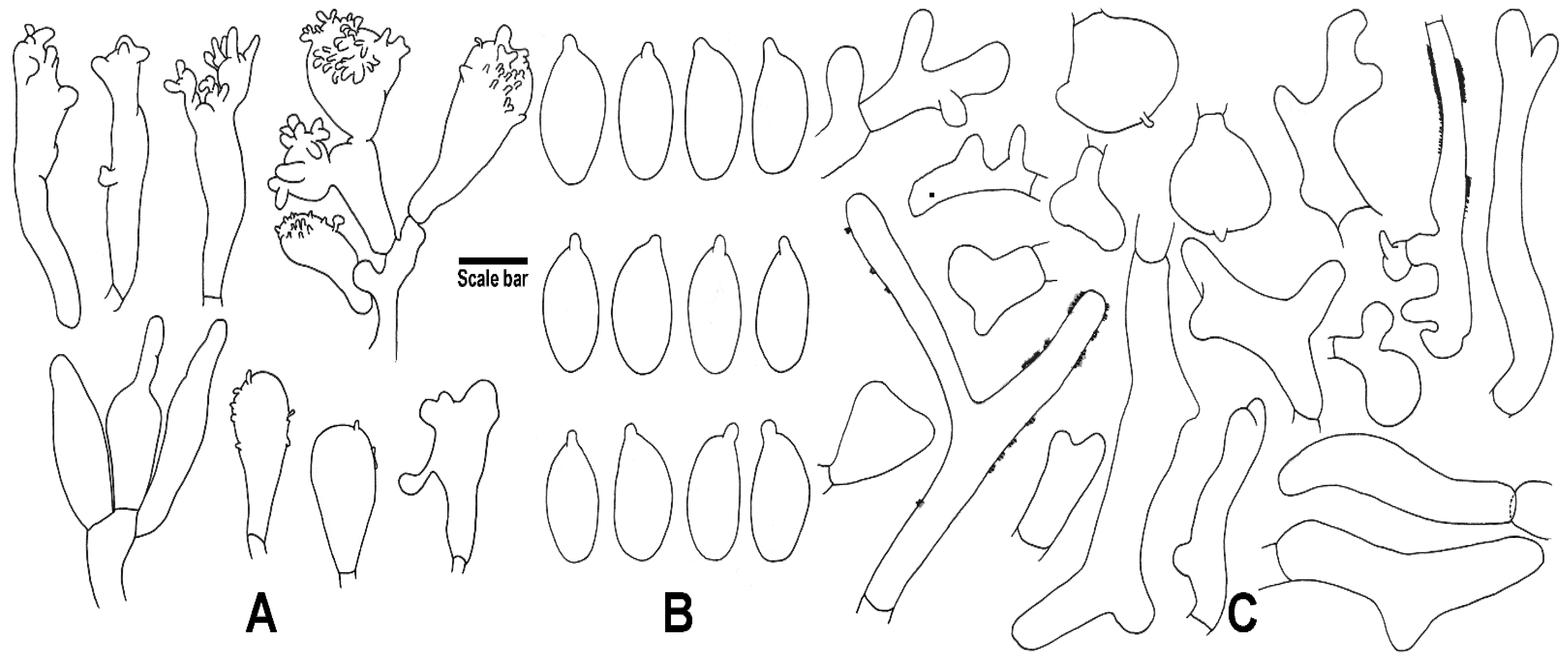
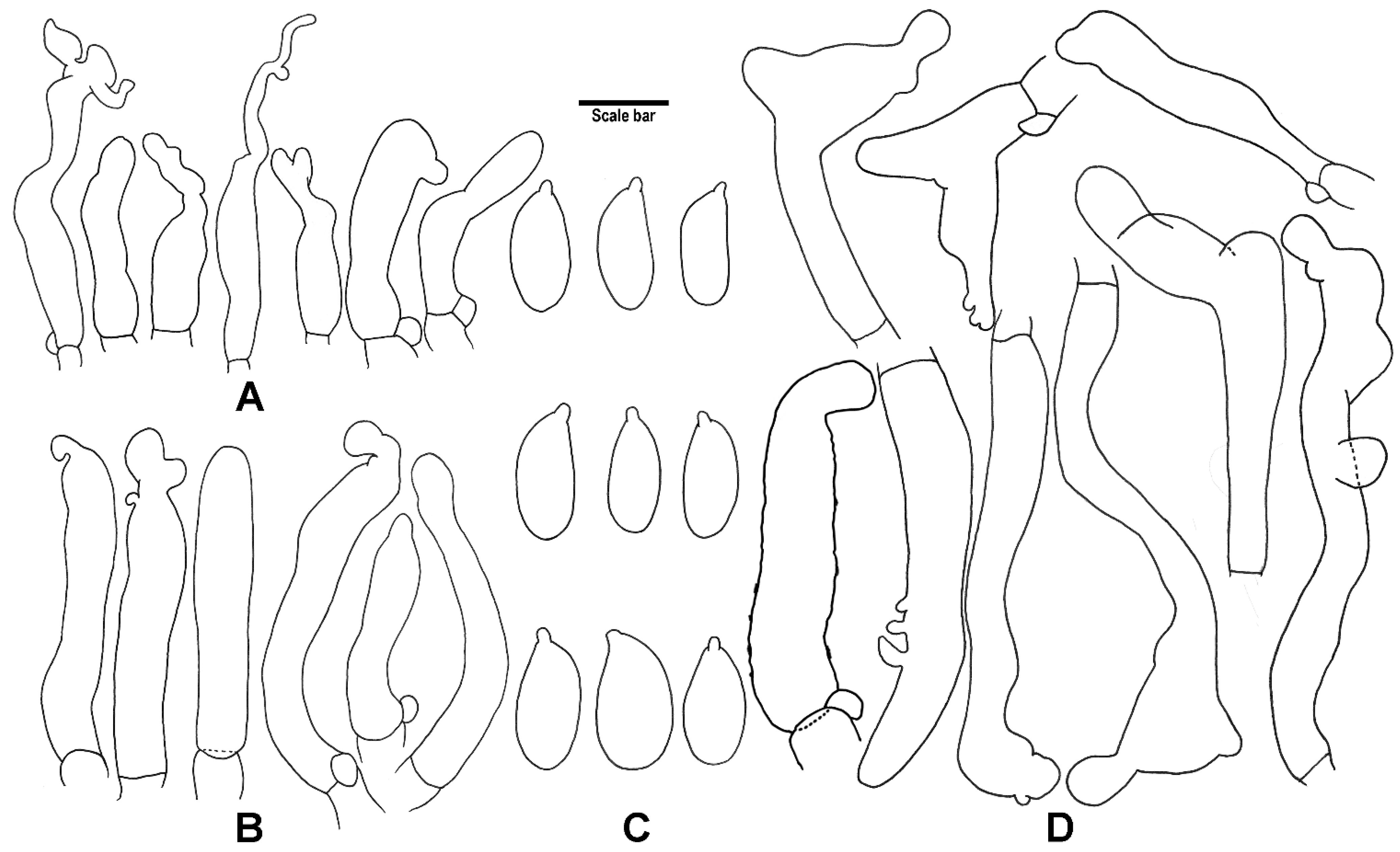
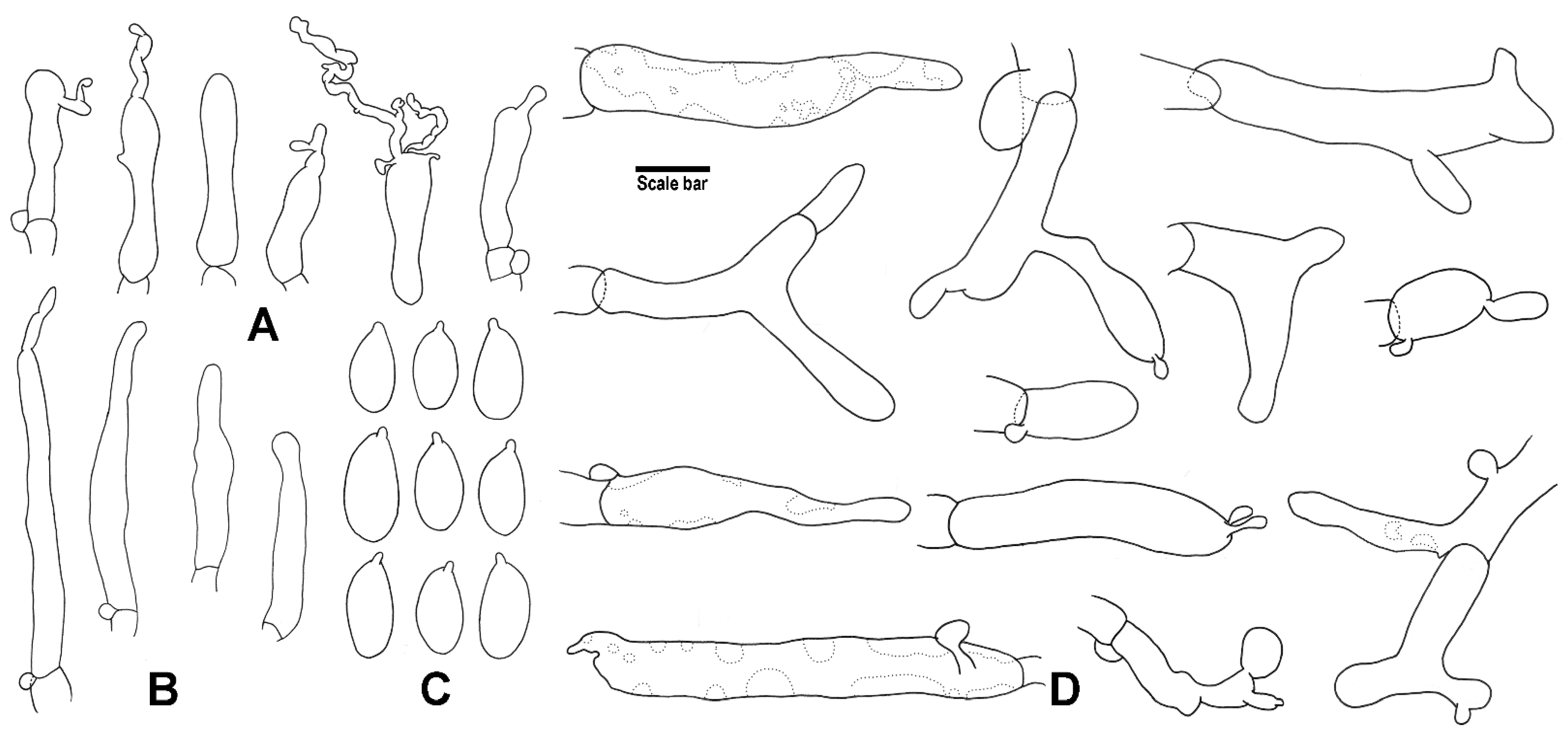
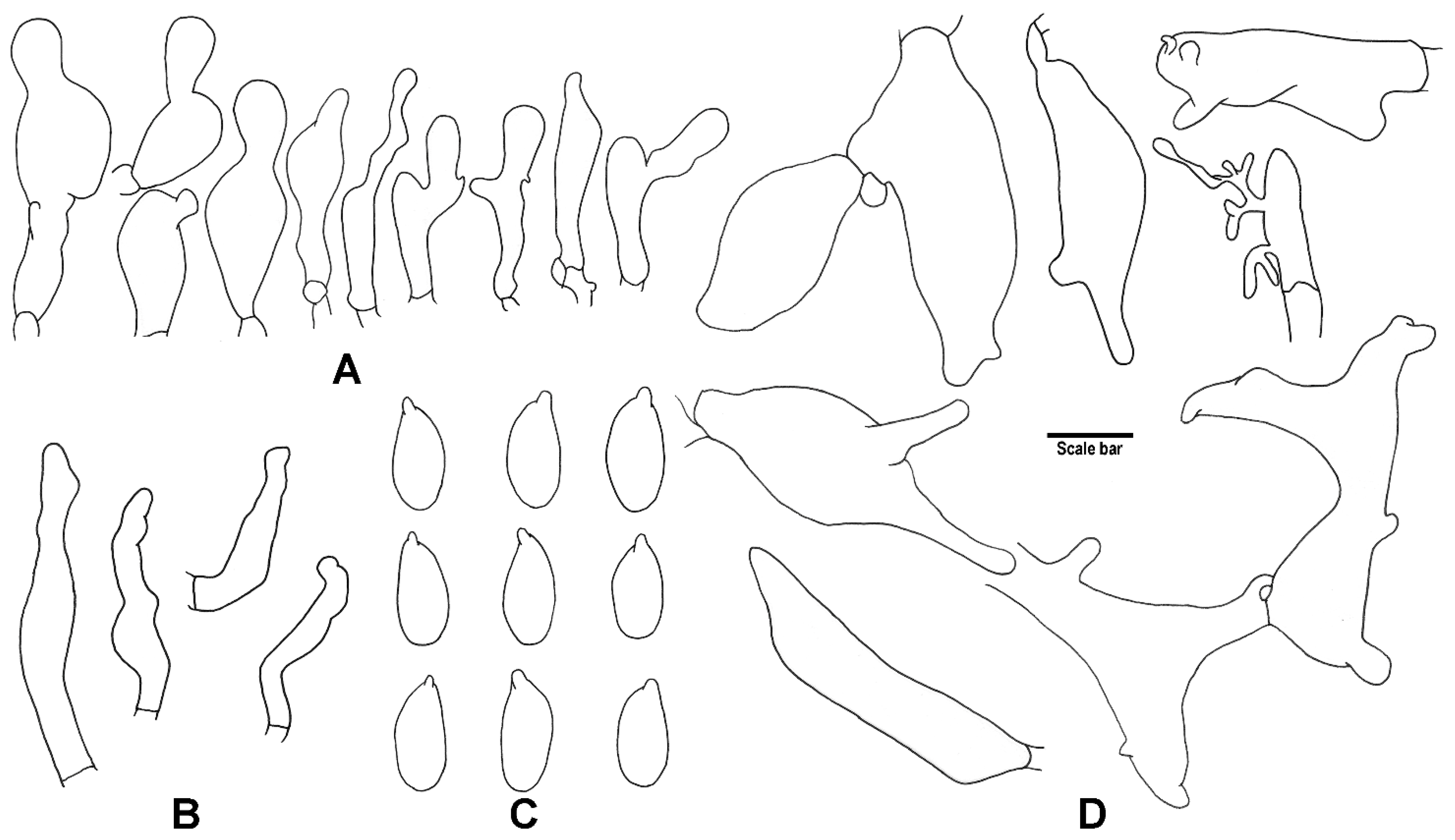
3. Results
4. Taxonomy
- Terminal cells of pileipellis broad, mostly inflated, mixed with irregularly branched elements and some resembling Dryophila-type structures ……………………………………………………………………………………………………………………………………………………………………………….. 2.
- −
- Terminal cells of pileipellis coralloid, more or less diverticulate, lobed to irregularly branched, or with Dryophila-type structures ……………………………………………………………………………………………………………………………………………………………………………….. 3.
- 2.
- Pileus generally deeply umbilicate; lamellae ventricose ………………………………………………………………………………………….... G. omphalinoides
- −
- Pileus more or less depressed; lamellae linear to arcuate ………………………………………………………………………………………….... G. schizophyllus
- 3.
- Rhizomorphs present, cheilocystidia consist of Siccus-type broom cells, stipitipellis with dextrinoid hyphae ………………………………………………. 4.
- −
- Rhizomorphs absent, cheilocystidia never a Siccus-type broom cell, stipitipellis without dextrinoid hyphae………………………………………………… 5.
- 4.
- Clamp connections present ……………………………………………………………………………………………………………………………………. G. pallipes
- −
- Clamp connections absent …………………………………………………………………………………………………………………………………….G. efibulatus
- 5.
- Basidiomata with unpleasant odour …………………………………………………………………………………………………………………………………... 6.
- −
- Basidiomata with negligible odour ……………………………………………………………………………………………………………………………………. 10.
- 6.
- Lamellae not close or crowded …………………………………………………………………………………………………………………………………………. 7.
- −
- Lamellae close or crowded ……………………………………………………………………………………………………………………………………………..... 9.
- 7.
- Pileus general white overall ………………………………………………………………………………………………………………………... G. alliifoetidissimus
- −
- Pileus not white …………………………………………………………………………………………………………………………………………………………… 8.
- 8.
- Pileus light brown, orange white to greyish orange when old ……………………………………………………………………………………………. G. similis
- −
- Pileus almost reddish lilac overall when drying ………………………………………………………………………………………………………………. G. iodes
- 9.
- Pileipellis consist of incrusted hyphae ………………………………………………………………………………………………………………. G. densilamellatus
- −
- Pileipellis without incrusted hyphae ……………………………………………………………………………………………………………....... G. sinopolyphyllus
- 10.
- Caulocystidia present ……………………………………………………………………………………………………………………………………... G. strigosipes
- −
- Caulocystidia not recorded …………………………………………………………………………………………………………………………………………..... 11.
- 11.
- Stipe smooth or tomentose …………………………………………………………………………………………………………………………………………... 12.
- −
- Stipe with hairs …………………………………………………………………………………………………………………………………………………….......... 15.
- 12.
- Pileipellis made up of smooth hyphae ……………………………………………………………………………………………………………………………… 13.
- −
- Pileipellis with incrusted hyphae …………………………………………………………………………………………………………………………………….... 14.
- 13.
- Basidia sterigmata extremely long, up to 32 µm ……………………………………………………………………………………………………… G. macrosporus
- −
- Basidia sterigmata normally long ………………………………………………………………………………………………………………………........... G. tiliicola
- 14.
- Basidia sterigmata extremely long, up to 33 µm …………………………………………………………………………………………………………… G. longus
- −
- Basidia sterigmata normally long ………………………………………………………………………………………………………………………….. G. globulosus
- 15.
- Pileus tomentose or pileipellis with incrusted hyphae …………………………………………………………………………………………………………… 16.
- −
- Pileus without tomenta and pileipellis made up of smooth hyphae ……………………………………………………………………………………………… 17.
- 16.
- Pileus tomentose, pileipellis made up of smooth hyphae …………………………………………………………………………………………… G. tomentosus
- −
- Pileus without tomenta, pileipellis with incrusted hyphae …………………………………………………………………………………… G. longisterigmaticus
- 17.
- Pileus estriate …………………………………………………………………………………………………………………………………………….... G. erythropus
- −
- Pileus striate …………………………………………………………………………………………………………………………………………………………....... 18.
- 18.
- Stipe longitudinally striate ………………………………………………………………………………………………………………………………....... G. striatus
- −
- Stipe smooth ………………………………………………………………………………………………………………………………………………. G. changbaiensis
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antonín, V.; Noordeloos, M.E. A monograph of Marasmius, Collybia and related genera in Europe. Part 2: Collybia, Gymnopus, Rhodocollybia, Crinipellus, Chaetocalathus and additions to Marasmiellus. Libr. Bot. 1997, 17, 1–256. [Google Scholar]
- Antonín, V.; Noordeloos, M.E. A Monograph of Marasmioid and Collybioid Fungi in Europe; IHV-Verlag: Berlin, Germany, 2010; 480p. [Google Scholar]
- Antonín, V.; Finy, P.; Tomšovský, M. Taxonomy of the Gymnopus inusitatus group and the new G. inusitatus var. cystidiatus from Hungary. Mycotaxon 2012, 119, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Antonín, V.; Sedlák, P.; Tomšovský, M. Taxonomy and phylogeny of European Gymnopus subsection Levipedes (Basidiomycota, Omphalotaceae). Persoonia 2013, 31, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrill, W.A. (Agaricales) Polyporaceae-Agaricaceae. In North American Flora; The New York Botanical Garden: New York, NY, USA, 1916; Volume 9, pp. 1–542. [Google Scholar]
- Halling, R.E. The genus Collybia (Agaricales) in the Northeastern United States and adjacent Canada. Mycol. Mem. 1983, 8, 1–148. [Google Scholar]
- Mata, J.L.; Hughes, K.W.; Petersen, R.H. An investigation of Omphalotaceae (Fungi: Euagarics) with emphasis on the genus Gymnopus. Sydowia 2007, 58, 191–289. [Google Scholar]
- Petersen, R.H.; Hughes, K.W. Two additional species of Gymnopus (Euagarics, Basidiomycotina). MycoKeys 2019, 45, 1–24. [Google Scholar] [CrossRef] [Green Version]
- César, E.; Montoya, L.; Bandala, V.M.; Ramos, A. Three new marasmioid-gymnopoid rhizomorph-forming species from Mexican mountain cloud forest relicts. Mycol. Prog. 2020, 19, 1017–1029. [Google Scholar] [CrossRef]
- Ryoo, R.; Antonín, V.; Ka, K.H. Marasmioid and gymnopoid fungi of the Republic of Korea. 8. Gymnopus section Levipedes. Mycobiology 2020, 48, 252–262. [Google Scholar] [CrossRef]
- Antonín, V.; Ryoo, R.; Ka, K.H. Marasmioid and gymnopoid fungi of the Republic of Korea. 7. Gymnopus sect. Androsacei. Mycol. Prog. 2014, 13, 703–718. [Google Scholar] [CrossRef]
- Ryoo, R.; Antonín, V.; Ka, K.H.; Tomšovský, M. Marasmioid and gymnopoid fungi of the Republic of Korea. 8. Gymnopus section Impudicae. Phytotaxa 2016, 286, 75–88. [Google Scholar] [CrossRef]
- Vizzini, A.; Antonin, V.; Sesli, E.; Contu, M. Gymnopus trabzonensis sp. nov. (Omphalotaceae) and Tricholoma virgatum var. fulvoumbonatum var. nov. (Tricholomataceae), two new white-spored agarics from Turkey. Phytotaxa 2015, 226, 119–130. [Google Scholar] [CrossRef]
- Corner, E.J.H. The agaric genera Marasmius, Chaetocalathus, Crinipellis, Heimiomyces, Resupinatus, Xerula and Xerulina in Malesia. Nova Hedwig. Beih. 1996, 111, 1–175. [Google Scholar]
- Wilson, A.W.; Desjardin, D.E.; Horak, E. Agaricales of Indonesia. 5. The genus Gymnopus from Java and Bali. Sydowia 2004, 56, 137–210. [Google Scholar]
- Li, J.P.; Li, Y.; Li, T.H.; Antonín, V.; Hosen, M.I.; Song, B.; Xie, M.L.; Feng, Z. A preliminary report of Gymnopus sect. Impudicae (Omphalotaceae) from China. Phytotaxa 2021, 497, 263–276. [Google Scholar] [CrossRef]
- Li, J.P.; Song, B.; Feng, Z.; Wang, J.; Deng, C.Y.; Yang, Y.H. A new species of Gymnopus sect. Androsacei (Omphalotaceae, Agaricales) from China. Phytotaxa 2021, 521, 1–14. [Google Scholar] [CrossRef]
- Li, J.P.; Antonín, V.; Gates, G.; Jiang, L.; Li, T.H.; Li, Y.; Song, B.; Deng, C.Y. Emending Gymnopus sect. Gymnopus (Agaricales, Omphalotaceae) by including two new species from southern China. Mycokeys 2022, 87, 183–204. [Google Scholar] [CrossRef]
- Cooper, J.; Leonard, P. Three new species of foetid Gymnopus in New Zealand. MycoKeys 2013, 7, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Halling, R.E. Notes on Collybia V. Gymnopus section Levipedes in tropical South America, with comments on Collybia. Brittonia 1996, 48, 487–494. [Google Scholar] [CrossRef]
- Coimbra, V.R.M.; Pinheiro, F.G.B.; Wartchow, F.; Gibertoni, T.B. Studies on Gymnopus sect. Impudicae (Omphalotaceae, Agaricales) from Northern Brazil: Two new species and notes on G. Montagnei. Mycol. Prog. 2015, 14, e110. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Perry, B.A. The gymnopoid fungi (Basidiomycota, Agaricales) from the Republic of São Tomé and Príncipe, West Africa. Mycosphere 2017, 8, 1317–1391. [Google Scholar] [CrossRef]
- Morrone, J.J. The biogeographical Andean Subregion: A proposal exemplified by Arthropod taxa (Arachnida: Crustacea, and Hexapoda). Neotropica 1996, 42, 103–114. [Google Scholar]
- Katinas, L.; Morrone, J.J.; Crisci, J.V. Track analysis reveals the composite nature of the Andean biota. Aust. J. Bot. 1999, 47, 111–130. [Google Scholar] [CrossRef]
- Antonín, V.; Halling, R.E.; Noordeloos, M.E. Generic concepts within the groups of Marasmius and Collybia sensu lato. Mycotaxon 1997, 63, 359–368. [Google Scholar]
- Vilgalys, R. Speciation and species concepts in the Collybia dryophila complex. Mycologia 1991, 83, 758–773. [Google Scholar] [CrossRef]
- Vilgalys, R.; Miller, O.K. Mating relationships within the Collybia dryophila group in Europe. Trans. Br. Mycol. Soc. 1987, 89, 295–300. [Google Scholar] [CrossRef]
- Wilson, A.W.; Desjardin, D.E. Phylogenetic relationships in the gymnopoid and marasmioid fungi (Basidiomycetes. Euagarics clade). Mycologia 2005, 97, 667–679. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.M.; Ge, Z.W.; Yang, Z.L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Antonín, V. Contribution to a monograph of marasmioid and collybioid fungi in Europe. Czech Mycol. 2008, 60, 21–27. [Google Scholar] [CrossRef]
- Oliveira, J.J.S.; Vargas-Isla, R.; Cabral, T.S.; Rodrigues, D.P.; Ishikawa, N.K. Progress on the phylogeny of the Omphalotaceae: Gymnopus s. str., Marasmiellus s. str., Paragymnopus gen. nov. and Pusillomyces gen. nov. Mycol. Prog. 2019, 18, 713–739. [Google Scholar] [CrossRef]
- Petersen, R.H.; Hughes, K.W. Collybiopsis and its type species, Co. ramealis. Mycotaxon 2021, 136, 263–349. [Google Scholar] [CrossRef]
- Hu, J.J.; Zhao, G.P.; Tuo, Y.L.; Rao, G.; Zhang, Z.H.; Qi, Z.X.; Yue, L.; Liu, Y.J.; Zhang, T.; Li, Y.; et al. Morphological and molecular evidence reveal eight new species of Gymnopus from Northeast China. J. Fungi 2022, 8, 349. [Google Scholar] [CrossRef]
- Karsten, P.A. Myceteo aliquot in Mongolia et China boreali a cl. C. N. Potanin lecti. Hedwigia 1892, 31, 38–40. [Google Scholar]
- Index Herbariorum. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 13 March 2022).
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; 252p. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; Volume 37, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Pruitt, K.D.; Schoch, C.L.; Sherry, S.T.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2021, 49, D92–D96. [Google Scholar] [CrossRef]
- Petersen, R.H.; Hughes, K.W. Micromphale sect. Perforantia (Agaricales, Basidiomycetes); expansion and phylogenetic placement. MycoKeys 2016, 18, 1–122. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Mata, J.L.; Hughes, K.W.; Petersen, R.H. Phylogenetic placement of Marasmiellus juniperinus. Mycoscience 2004, 45, 214–221. [Google Scholar] [CrossRef]
- Hughes, K.W.; Petersen, R.H.; Lickey, E.B. Using heterozygosity to estimate a percentage DNA sequence similarity for environmental species’ delimitation across basidiomycete fungi. New Phytol. 2009, 182, 795–798. [Google Scholar] [CrossRef]
- Mitsuhashi, S.; Shindo, C.; Shigetomi, K.; Miyamoto, T.; Ubukata, M. (+)-Epogymnolactam, a novel autophagy inducer from mycelial culture of Gymnopus sp. Phytochemistry 2015, 114, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Osono, T.; Matsuoka, S.; Hirose, D. Diversity and geographic distribution of ligninolytic fungi associated with Castanopsis sieboldii leaf litter in Japan. Front. Microbiol. 2020, 11, 595427. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.H.; Hughes, K.W. An investigation on Mycetinis (Euagarics, Basidiomycota). MycoKeys 2017, 24, 1–138. [Google Scholar] [CrossRef] [Green Version]
- Desjardin, D.E.; Retnowati, A.; Horak, E. Agaricales of Indonesia. 2. A preliminary monograph of Marasmius from Java and Bali. Sydowia 2000, 54, 92–194. [Google Scholar]
- Desjardin, D.E.; Horak, E. Marasmius and Gloiocephala in the South Pacific Region: Papua New Guinea, New Caledonia, and New Zealand taxa. In Taxonomic Monographs of Agaricales II; Petrini, O., Petrini, L.E., Horak, E., Eds.; Schweizerbart and Borntraeger science publishers: Stuttgart, Germany, 1997; Volume 168, pp. 1–81. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Collection No. | GenBank Accession No | References | |
|---|---|---|---|---|
| ITS | LSU | |||
| G. adventitius nom. prov. | SFSU: DED8813 | KY026760 | KY026760 | [42] |
| G. alliifoetidissimus * | GDGM 76695 | MT023348 | MT017526 | [16] |
| G. androsaceus | CULTENN5609 | KY026750 | KY026750 | [42] |
| G. androsaceus | CBS 240.53 | MH857175 | MH868714 | [43] |
| G. androsaceus | CBS 239.53 | MH857174 | MH868713 | [43] |
| G. androsaceus | CULTENN5021h2 | KY026748 | KY026748 | [42] |
| G. androsaceus | TENN: F-59594 | KY026663 | KY026663 | [42] |
| G. atlanticus * | URM 87728 | KT222654 | KY302698 | [21] |
| G. aurantiipes | SFSU: AWW118 | AY263432 | AY639410 | [15] |
| G. brunneiniger | XAL: Cesar50 | MT232388 | MW187069 | [9] |
| G. brunneodiscus | BRNM 714974 | MH589973 | MH589988 | [10] |
| G. cremeostipitatus * | BRNM 747547 | KF251071 | KF251091 | [11] |
| G. densilamellatus | BRNM 714927 | KP336685 | KP336694 | [12] |
| G. dryophiloides * | BRNM 781447 | MH589967 | MH589985 | [10] |
| G. dryophilus | TENN: F-57012 | DQ241781 | AY640619 | [29] |
| G. dysodes | TENN: F-61125 | KY026666 | FJ750265 | [42] |
| G. dysodes | BRNM 766741 | KP336693 | KP336701 | [12] |
| G. efibulatus * | HGASMF01-7052 haplotype1 | OM970865 | OM970865 | This study |
| G. efibulatus * | HGASMF01-7052 haplotype2 | OM970866 | OM970866 | This study |
| G. efibulatus | HGASMF01-11995 | OM970873 | OM970877 | This study |
| G. foetidus | TENN: F-69323 | KY026739 | KY026739 | [42] |
| G. frigidomarginatus nom. prov. | TENN: F-55679 | KY026648 | KY026648 | [42] |
| G. fusipes | TENN: F-59217 | AY256710 | AY256710 | [44] |
| G. impudicus | BRNM 714849 | LT594119 | LT594119 | [12] |
| G. inflatotrama nom. prov. | TENN: F-48143 | KY026619 | KY026619 | [42] |
| G. inflatotrama nom. prov. | TFB 4529 | KY026744 | KY026744 | [42] |
| G. inflatotrama nom. prov. | TENN: F-53490 | KY026640 | KY026640 | [42] |
| G. inflatotrama nom. prov. | TENN: F-51233 | KY026632 | KY026632 | [42] |
| G. inusitatus * | BCN: SCM B-4058 | JN247553 | JN247557 | [3] |
| G. iocephalus | Duke RV94154 | DQ449986 | unavailable | [7] |
| G. iocephalus | TENN: F-52970 | DQ449984 | KY019630 | [7] |
| G. iodes | HGASMF01-10069 | OM970868 | OM970868 | This study |
| G. iodes * | HGASMF01-10068 | OM970869 | OM970869 | This study |
| G. iodes | HMJAU 60388 | OM970870 | OM970870 | This study |
| G. irresolutus * | SFSU: DED 8209 | MF100973 | unavailable | [22] |
| G. montagnei | JMCR 143 | DQ449988 | AF261327 | [7] |
| G. neobrevipes | TENN: F-14505 | MH673477 | MH673477 | [8] |
| G. novae-angliae nom. prov. | CULTENN4975 | KY026745 | KY026745 | [42] |
| G. novomundi nom. prov. | SFSU-DED5097 | KY026759 | KY026759 | [42] |
| G. ocior | TENN: F-65135 | KY026678 | KY026678 | [42] |
| G. omphalinoides * | GDGM 78318 | MW134044 | MW134730 | [18] |
| G. pallipes * | GDGM 81513 | MW582856 | OK087327 | [17,18] |
| G. polyphyllus | TENN62814-H1 | FJ596894 | unavailable | [45] |
| G. polyphyllus | TENN62814-H2 | FJ596895 | unavailable | [45] |
| G. schizophyllus * | GDGM 77165 | MW134043 | MW134729 | [18] |
| G. semihirtipes | TENN: F-07595 | OK376741 | unavailable | GenBank |
| G. similis | BRNM 714981 | KP336690 | KP336697 | [12] |
| G. sinopolyphyllus | HMJAU 60387 | OM970871 | OM970871 | This study |
| G. sinopolyphyllus * | HMJAU 60386 | OM970872 | OM970872 | This study |
| G. sp. | WCS023 | AB968433 | unavailable | [46] |
| G. sp. | Ta-BL62 | LC505290 | LC505290 | [47] |
| G. spongiosus | TENN: F-65912 | KY026687 | KY026687 | [42] |
| G. spongiosus | TENN: F-65912 | KY026686 | KY026686 | [42] |
| G. strigosipes * | HMAS 295796 | OM970874 | OM970874 | This study |
| G. strigosipes | HMAS 295797 | OM970867 | OM970867 | This study |
| G. subsupinus | PDD: 96595 | KM975399 | KM975375 | GenBank |
| G. variicolor | BRNM 714959 | LT594121 | KP348011 | [12] |
| Ma. androsaceus | Sara Landvik: NN008037 | JN943605 | JN941145 | [11] |
| Ma. otagensis | PDD: 106823 | MT974597 | MT974601 | [18] |
| Ma. sp. | Sw5-1 | LC504952 | unavailable | GenBank |
| My. alliaceus | TENN: F-55630 | KY696752 | KY696752 | [48] |
| My. scorodonius | TENN: F-53474 | KY696748 | KY696748 | [48] |
| Pa. perforans | TENN: F-50319 | KY026625 | KY026625 | [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.-P.; Pan, M.-C.; Li, Y.; Deng, C.-Y.; Wang, X.-M.; Zhang, B.-X.; Li, C.-T.; Li, Y. Morpho-Molecular Evidence Reveals Four Novel Species of Gymnopus (Agaricales, Omphalotaceae) from China. J. Fungi 2022, 8, 398. https://doi.org/10.3390/jof8040398
Li J-P, Pan M-C, Li Y, Deng C-Y, Wang X-M, Zhang B-X, Li C-T, Li Y. Morpho-Molecular Evidence Reveals Four Novel Species of Gymnopus (Agaricales, Omphalotaceae) from China. Journal of Fungi. 2022; 8(4):398. https://doi.org/10.3390/jof8040398
Chicago/Turabian StyleLi, Ji-Peng, Mei-Chen Pan, Yi Li, Chun-Ying Deng, Xiao-Min Wang, Bang-Xi Zhang, Chang-Tian Li, and Yu Li. 2022. "Morpho-Molecular Evidence Reveals Four Novel Species of Gymnopus (Agaricales, Omphalotaceae) from China" Journal of Fungi 8, no. 4: 398. https://doi.org/10.3390/jof8040398
APA StyleLi, J. -P., Pan, M. -C., Li, Y., Deng, C. -Y., Wang, X. -M., Zhang, B. -X., Li, C. -T., & Li, Y. (2022). Morpho-Molecular Evidence Reveals Four Novel Species of Gymnopus (Agaricales, Omphalotaceae) from China. Journal of Fungi, 8(4), 398. https://doi.org/10.3390/jof8040398