
Elevated Ozone Concentration and Nitrogen Addition Increase Poplar Rust Severity by Shifting the Phyllosphere Microbial Community


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Ozone Fumigation Treatment and Nitrogen Addition
2.3. Evaluation of Poplar Foliar Rust Severity
2.4. Leaf Samples Collection, DNA Extraction and Illumina Amplicon Sequencing
2.5. Bioinformatics and Statistical Analysis
3. Results
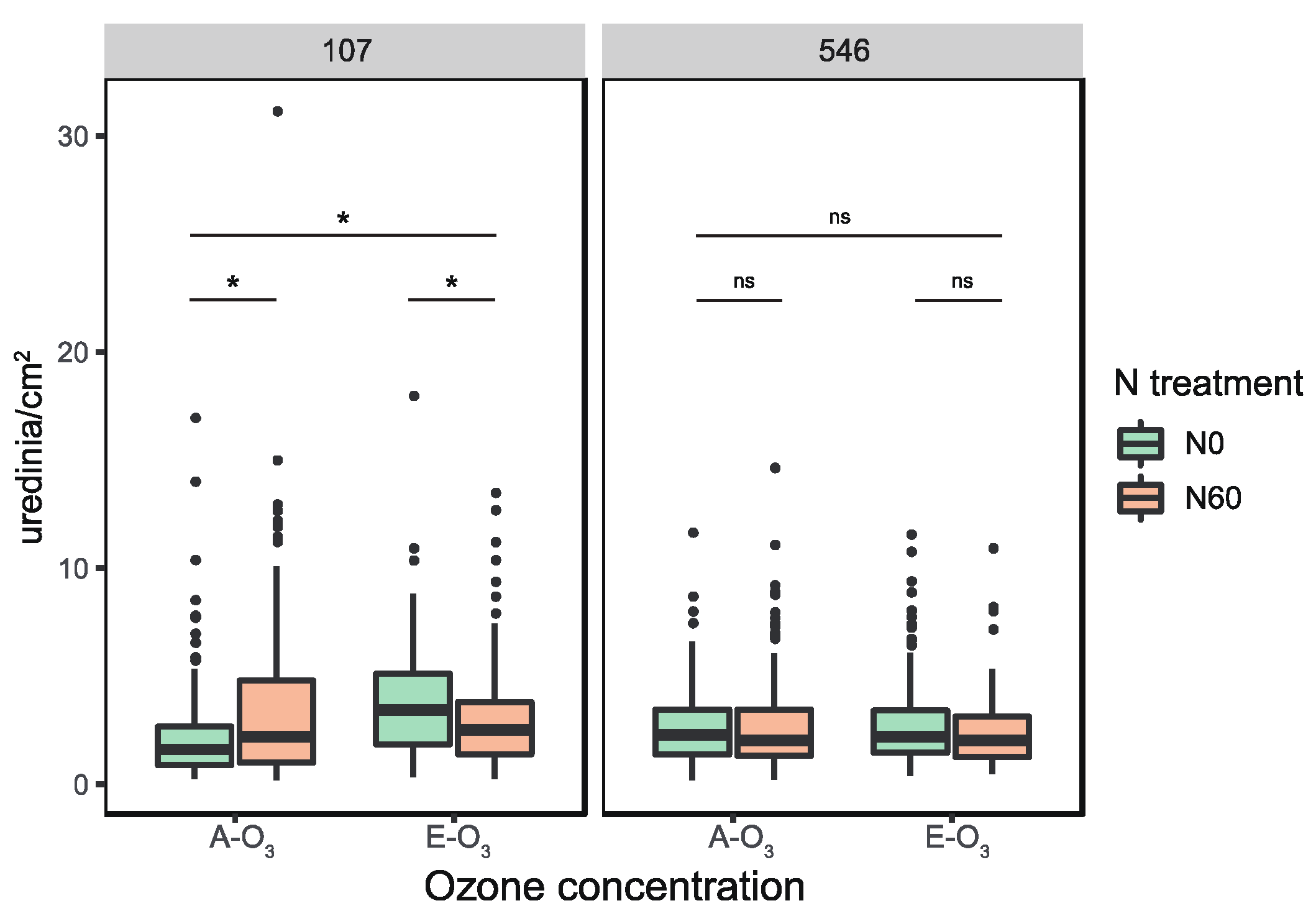
3.1. Combined Effects of Elevated O3 Concentration and N Addition on Foliar Rust Severity of Two Poplar Clones
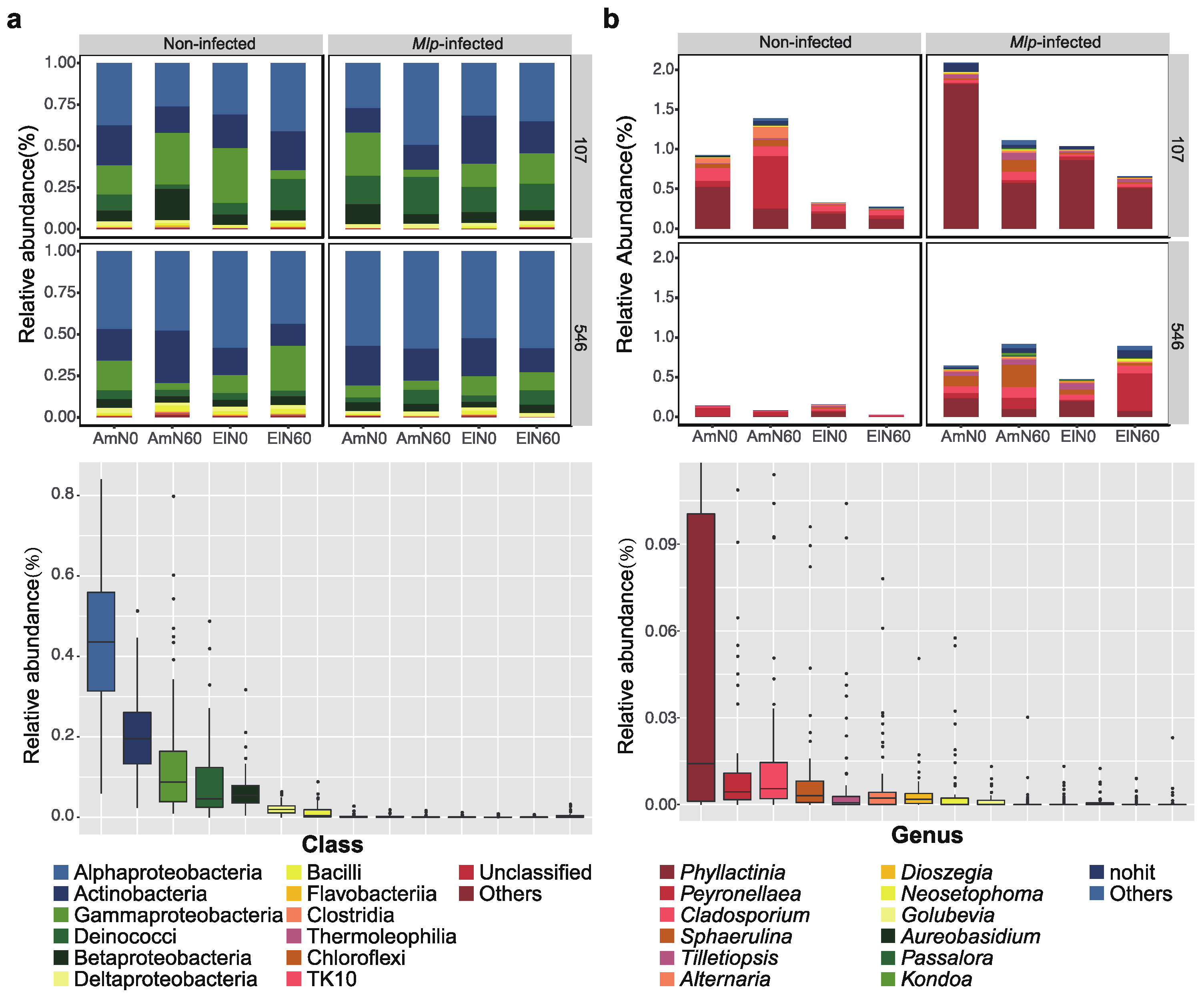
3.2. The Relative Abundance of Phyllosphere Fungal and Bacterial Species under Elevated O3 Concentration and N Addition
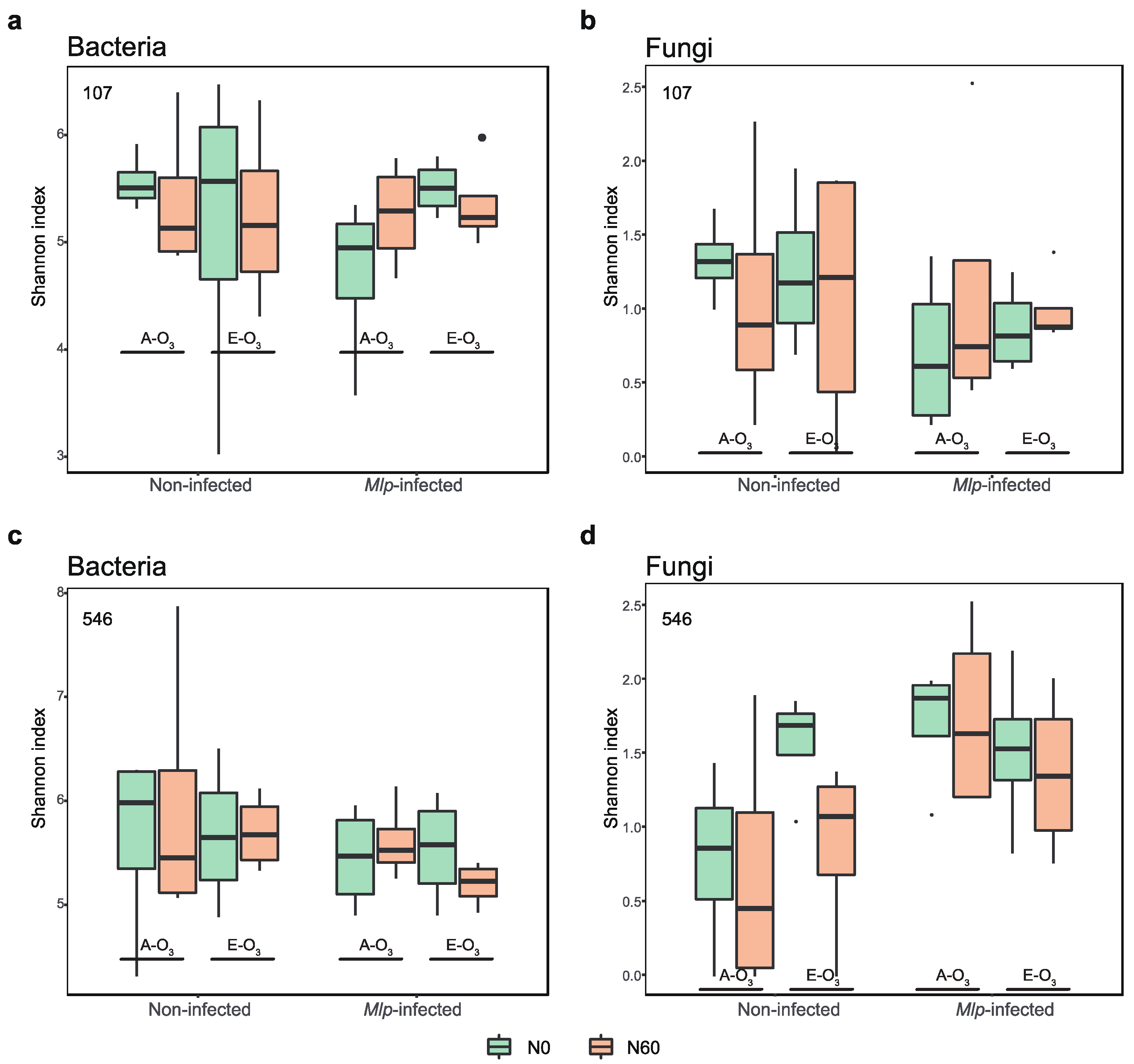
3.3. The Variations of Bacterial and Fungal α-Diversity under Elevated O3 Concentration and N Addition
3.4. The Variation of Bacterial and Fungal β-Diversity under Elevated O3 Concentration and N Addition
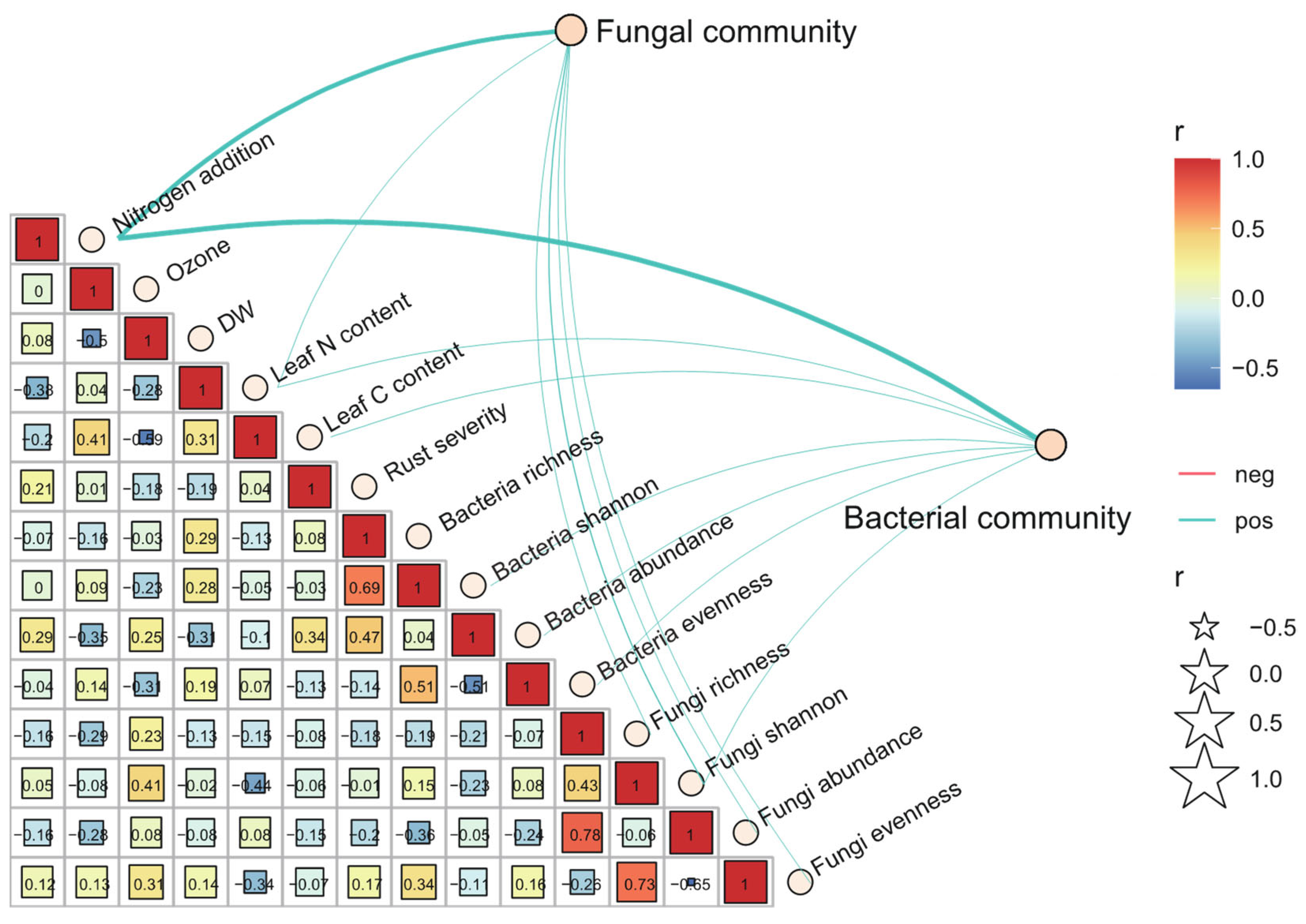
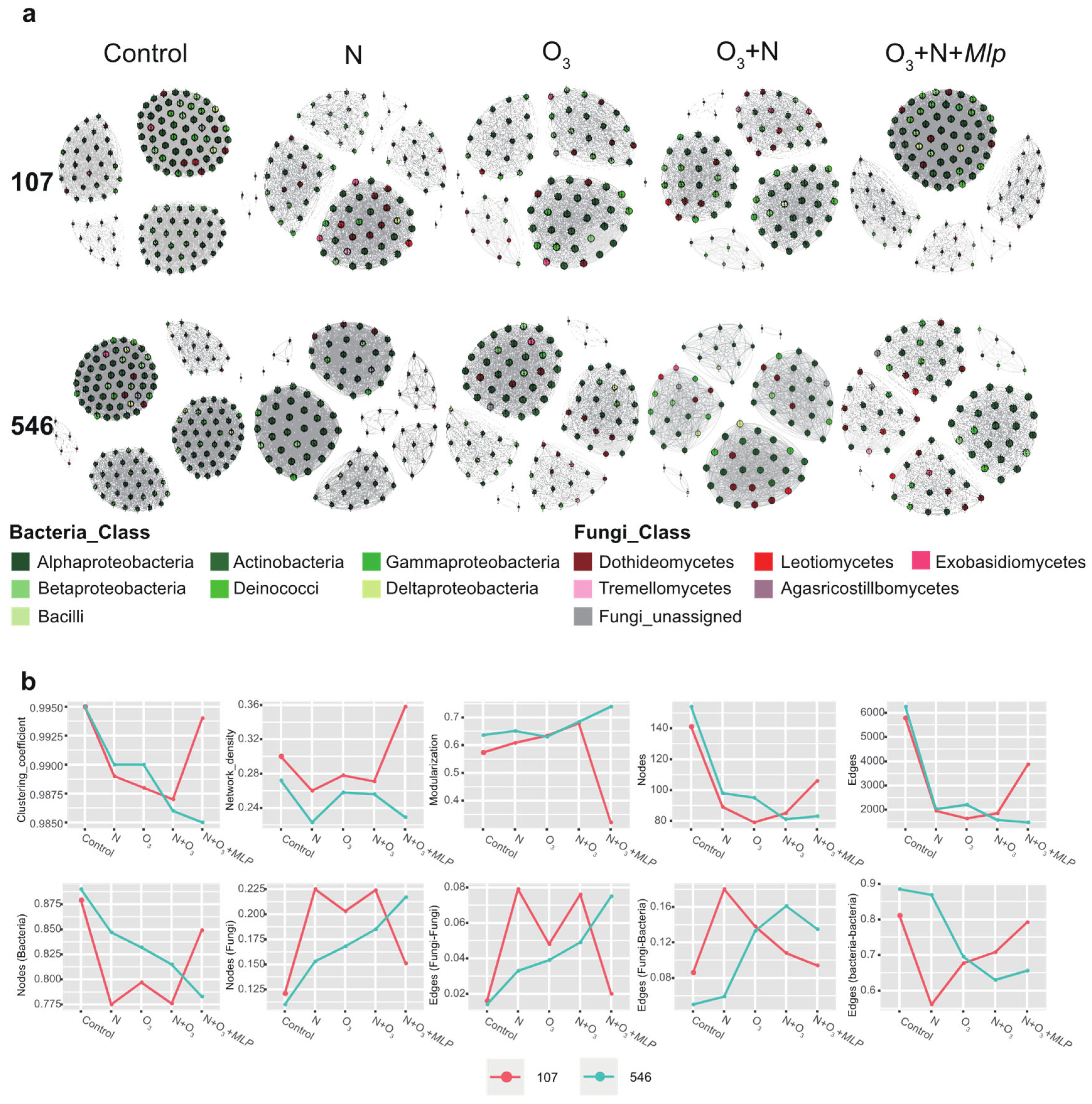
3.5. Co-Occurrence between Poplar Phyllosphere Microbiomes
4. Discussion
4.1. A Trade-Off of Mlp-Susceptibility to Elevated O3 and N Addition for the ‘107’ Poplar
4.2. The Composition of Poplar Phyllosphere Microbiome Shifts under Elevated O3, N Addition and Melampsora larici-populina Infection
4.3. Elevated O3 and N Addition Lead to Distinct Responses in the Phyllosphere Microbial Community Diversity
4.4. Effects of Elevated O3, N Addition and the Combination of Rust Infection on Co-Occurrence of Phyllosphere Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinon, J.; Frey, P. Interactions between Poplar clones and Melampsora populations and their implications for breeding for durable resistance. In Rust Diseases of Willow and Poplar; Pei, M.H., McCracken, A.R., Eds.; CABI Publishing: Wallingford, UK, 2015; pp. 139–155. [Google Scholar]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [PubMed] [Green Version]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.C.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Nat. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacquard, S.; Petre, B.; Frey, P.; Hecker, A.; Rouhier, N.; Duplessis, S. The poplar-poplar rust interaction: Insights from genomics and transcriptomics. J. Pathog. 2011, 2011, e716041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacquard, S.; Joly, D.L.; Lin, Y.C.; Tisserant, E.; Feau, N.; Delaruelle, C.; Legué, V.; Kohler, A.; Tanguat, P.; Petre, B.; et al. A comprehensive analysis of genes encoding small-secreted proteins identifies candidate effectors in Melampsora larici-populina (Poplar leaf rust). Mol. Plant Microbe Interact. 2012, 25, 279–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacquard, S.; Delaruelle, C.; Frey, P.; Tisserant, E.; Kohler, A.; Duplessis, S. Transcriptome analysis of poplar rust telia reveals overwintering adaptation and tightly coordinated karyogamy and meiosis processes. Front. Plant Sci. 2013, 4, 456. [Google Scholar] [CrossRef] [Green Version]
- Persoons, A.; Morin, E.; Delaruelle, C.; Payen, T.; Halkett, F.; Frey, P.; De Mita, S.; Duplessis, S. Patterns of genomic variation in the poplar rust fungus Melampsora larici-populina identify pathogenesis-related factors. Front. Plant Sci. 2014, 5, 450. [Google Scholar] [CrossRef] [Green Version]
- Petre, B.; Saunders, D.G.O.; Sklenar, J.; Lorrain, C.; Win, J.; Duplessis, S.; Kamoun, S. Candidate effector proteins of the rust pathogen Melampsora laricipopulina target diverse plant cell compartments. Mol. Plant Microbe Interact. 2015, 28, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Petre, B.; Lorrain, C.; Saunders, D.G.O.; Win, J.; Sklenar, J.; Duplessis, S.; Kamoun, S. Rust fungal effectors mimic host transit peptides to translocate into chloroplasts. Cell. Microbiol. 2016, 18, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Lorrain, C.; Marchal, C.; Hacquard, S.; Delaruelle, C.; Pétrowski, J.; Petre, B.; Hecker, A.; Frey, P.; Duplessis, S. The rust fungus Melampsora larici-populina expresses a conserved genetic program and distinct sets of secreted protein genes during infection of its two host plants, larch and poplar. Mol. Plant Microbe Interact. 2018, 31, 695–706. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Gill, S.S.; Gill, R. Plant Adaptation to Environmental Change: Significance of Amino Acids and Their Derivatives, 1st ed.; CABI: Wallingford, UK, 2014. [Google Scholar]
- Li, P.; De Marco, A.; Feng, Z.; Anav, A.; Zhou, D.; Paoletti, E. National wide ground-level ozone measurements in China suggest serious risks to forests. Environ. Pollut. 2018, 237, 803–813. [Google Scholar] [CrossRef]
- Feng, Z.Z.; Sun, J.S.; Wan, W.X.; Hu, E.Z.; Calatayud, V. Evidence of widespread ozone-induced visible injury on plants in Beijing, China. Environ. Pollut. 2014, 1932, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.Z.; Shang, B.; Gao, F.; Calatayud, V. Current ambient and elevated ozone effects on poplar: A global meta-analysis and response relationships. Sci. Total Environ. 2019, 654, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Lee, H.I.; Creelman, R.A.; Mullet, J.E.; Davis, K.R. Jasmonate perception desensitises O3-induced salicylic acid biosynthesis and programmed cell death in Arabidopsis. Plant Cell 2000, 12, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Heagle, A.S. Interactions between air pollutants and plant parasites. Ann. Rev. Phytopathol. 1970, 11, 365–388. [Google Scholar] [CrossRef]
- Coleman, J.S.; Jones, C.G.; Smith, W.H. The effect of ozone on cottonwood-leaf rust interactions: Independence of abiotic stress, genotype, and leaf ontogeny. Can. J. Bot 1987, 65, 949–953. [Google Scholar] [CrossRef]
- Beare, J.A.; Archer, S.A.; Bell, J.N.B. Effects of Melampsora leaf rust disease and chronic ozone exposure on poplar. Environ. Pollut. 1999, 105, 419–426. [Google Scholar] [CrossRef]
- Heagle, A.S.; Key, L.W. Effects of ozone on the wheat stem rust fungus. Phytopathology 1973, 63, 397–400. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager Møller, I.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Niu, S.L.; Classen, A.T.; Dukes, J.S.; Kardol, P.; Liu, L.L.; Luo, Y.Q.; Rustad, L.; Sun, J.; Tang, J.W.; Templer, P.H.; et al. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol. Lett. 2016, 19, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Huber, D.; Watson, R. Nitrogen form and plant disease. Ann. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef]
- Lin, Z.Y.; Wang, J.H.; Bao, Y.X.; Guo, Q.; Powell, C.A.; Xu, S.Q.; Chen, B.S.; Zhang, M.Q. Deciphering the transcriptomic response of fusarium verticillioides in relation to nitrogen availability and the development of sugarcane pokkah boeng disease. Sci. Rep. 2016, 6, 29692. [Google Scholar] [CrossRef] [Green Version]
- Ash, G.J.; Brown, J.F. Effect of nitrogen nutrition of the host on the epidemiology of Puccinia striiformis f. sp. tritici and crop yield in wheat. Plant Pathol. 1991, 20, 108–114. [Google Scholar]
- Devadasa, R.; Simpfendorferb, S.; Backhousec, D.; Lamb, D.W. Effect of stripe rust on the yield response of wheat to nitrogen. Crop J. 2014, 2, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.X.; Li, L.; Tang, L.; Zheng, Y.; Yi, Z.; Li, Y.-J.; Zhang, C.-C.; Zhang, F.S. Effect of nitrogen addition on nitrogen nutrition and strip rust occurrence of wheat in wheat/faba bean intercropping system. J. Nucl. Agric. Sci. 2013, 27, 1020–1028. [Google Scholar]
- Zhu, J.H.; Dong, Y.; Xiao, J.X.; Zheng, Y.; Tang, L. Effects of N application on wheat powdery mildew occurrence, nitrogen accumulation and allocation in intercropping system. Chin. J. Appl. Ecol. 2017, 28, 3985–3993. [Google Scholar]
- Shang, B.; Feng, Z.Z.; Gao, F.; Calatayud, V. The ozone sensitivity of five poplar clones is not related to stomatal conductance, constitutive antioxidant levels and morphology of leaves. Sci. Total Environ. 2020, 699, 134402. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Last, F.T. Seasonal incidence of Sporobolomyces on cereal leaves. Trans. Br. Mycol. Soc. 1955, 38, 221–239. [Google Scholar] [CrossRef]
- Last, F.T.; Deighton, F.C. The non-parasitic microflora on the surfaces of living leaves. Trans. Br. Mycol. Soc. 1965, 48, 83–99. [Google Scholar] [CrossRef]
- Leben, C. Epiphytic microorganisms in relation to plant disease. Annu. Rev. Phytopathol. 1965, 3, 209–230. [Google Scholar] [CrossRef]
- Rastogi, G.; Coaker, G.L.; Leveau, J.H.J. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol. Lett. 2013, 348, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Busby, P.E.; Peay, K.G.; Newcombe, G. Common foliar fungi of Populus trichocarpa modify Melampsora rust disease severity. New Phytol. 2016, 209, 1681–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Métraux, J.-P.; L’Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.; Bodenhausen, N.; Gruissem, W.; Vorholt, J.A. The Arabidopsis leaf transcriptome reveals distinct but also overlapping responses to colonization by phyllosphere commensals and pathogen infection with impact on plant health. New Phytol. 2016, 212, 192–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejed, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root- inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Naseri, B.; Ansari Hamadani, S. Characteristic agro-ecological features of soil populations of bean root rot pathogens. Rhizosphere 2017, 3, 203–208. [Google Scholar] [CrossRef]
- Tabande, L.; Naseri, B. How strongly is rhizobial nodulation associated with bean cropping system. J. Plant Protect. Res. 2020, 60, 176–184. [Google Scholar]
- Zhang, J.; Tang, H.; Zhu, J.; Lin, X.; Feng, Y. Divergent responses of methanogenic archaeal communities in two rice cultivars to elevated ground-level O3. Environ. Pollut. 2016, 213, 127–134. [Google Scholar] [CrossRef]
- Feng, Y.; Lin, X.; Yu, Y.; Zhang, H.; Chu, H.; Zhu, J. Elevated ground-level O3 negatively influences paddy methanogenic archaeal community. Sci. Rep. 2013, 3, 3193. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yu, Y.; Tang, H.; Zu, Q.; Zhu, J.; Lin, X. The contrasting responses of soil microorganisms in two rice cultivars to elevated ground-level ozone. Environ. Pollut. 2015, 197, 195–202. [Google Scholar] [CrossRef]
- Turner, N.C.; Waggoner, P.E.; Rich, S. Removal of ozone from the atmosphere by soil and vegetation. Nature 1974, 250, 486–489. [Google Scholar] [CrossRef]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, Y.; Frindte, K.; Knief, C.; Ashrafuzzaman, M.; Frei, M. Effects of Elevated Tropospheric Ozone Concentration on the Bacterial Community in the Phyllosphere and Rhizoplane of Rice. PLoS ONE 2016, 11, e0163178. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, X.X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Xu, Y.S.; Feng, Z.Z.; Shang, B.; Dai, L.L.; Udding, J.; Tarvainen, L. Mesophyll conductance limitation of photosynthesis in poplar under elevated ozone. Sci. Total Environ. 2019, 657, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Kobayashi, K.; Zhu, J.G. Yield and photosynthetic characteristics of flag leaves in Chinese rice (Oryza sativa L.) varieties subjected to free-air release of ozone. Agric. Ecosyst. Environ. 2009, 132, 203–211. [Google Scholar] [CrossRef]
- Shang, B.; Xu, Y.S.; Dai, L.L.; Yuan, X.Y.; Feng, Z.Z. Elevated ozone reduced leaf nitrogen allocation to photosynthesis in poplar. Sci. Total Environ. 2019, 657, 169–178. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, Y.; Yan, P.; Zhang, X.; Guo, X.; et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A.; et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Inter. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Jiao, S.; Xu, Y.Q.; Zhang, J.; Hao, X.; Lu, Y.H. Core Microbiota in Agricultural Soils and Their Potential Associations with Nutrient Cycling. mSystems 2019, 4, e00313-18. [Google Scholar] [CrossRef] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open-source software for exploring and manipulating networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Percy, K.E.; Awmack, C.S.; Lindroth, R.L.; Kubiske, M.E.; Kopper, B.J.; Isebrands, J.G.; Pregitzer, K.S.; Hendrey, G.P.; Dickson, R.E.; Zak, D.R.; et al. Altered performance of forest pests under atmospheres enriched by CO2 and O3. Nature 2002, 420, 403–407. [Google Scholar] [CrossRef]
- Coleman, J.S.; Jones, C.G.; Smith, W.H. Interactions between an acute ozone dose, eastern cottonwood and Marssonina leaf spot: Implications for pathogen community dynamics. Can. J. For. Res. 1988, 66, 863–868. [Google Scholar] [CrossRef]
- Helfer, S. Rust fungi and global change. New Phytol. 2014, 201, 770–780. [Google Scholar] [CrossRef]
- Sandermann, H., Jr.; Ernst, D.; Heller, W.; Langebartels, C. Ozone: An abiotic elicitor of plant defence reactions. Trends Plant Sci. 1988, 3, 47–50. [Google Scholar] [CrossRef]
- Eastburn, D.M.; McElrone, A.J.; Bilgin, D.D. Influence of atmospheric and climatic change on plant–pathogen interactions. Plant Pathol. 2011, 60, 54–69. [Google Scholar] [CrossRef]
- Karnosky, D.F.; Percy, K.E.; Xiang, B.; Callan, B.; Noormets, A.; Mankovska, B.; Hopkin, A.; Sober, J.; Jones, W.; Dickson, R.E.; et al. Interacting elevated CO2 and tropospheric O3 predisposes aspen (Populus tremuloides Michx.) to infection by rust (Melampsora medusae f. sp. tremuloidae). Glob. Change Biol. 2002, 8, 329–338. [Google Scholar] [CrossRef]
- Strengbom, J.; Nordin, A.; Näsholm, T.; Ericson, L. Parasitic fungus mediates change in nitrogen-exposed boreal forest vegetation. J. Ecol. 2002, 90, 61–67. [Google Scholar] [CrossRef]
- Leveau, J.H. A brief from the leaf: Latest research to inform our understanding of the phyllosphere microbiome. Curr. Opin. Microbiol. 2019, 49, 41–49. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Perez-Jaramillo, J.E.; de Hollander, M.; Ramirez, C.A.; Mendes, R.; Raaijmakers, J.M.; Carrion, V.J. Deciphering rhizosphere microbiome assembly of wild and modern common bean (Phaseolus vulgaris) in native and agricultural soils from Colombia. Microbiome 2019, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Brettell, L.E.; Singh, B. Linking the Phyllosphere Microbiome to Plant Health. Trends Plant Sci. 2020, 25, 841–844. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, P.T.; Whiteman, N.K. Insect herbivory reshapes a native leaf microbiome. Nat. Ecol. Evol. 2020, 4, 221–229. [Google Scholar] [CrossRef]
- Busby, P.E.; Aime, M.C.; Newcombe, G. Foliar pathogens of Populus angustifolia are consistent with a hypothesis of Beringian migration into North America. Fungal Biol. 2012, 116, 792–801. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Nat. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridout, M.; Newcombe, G. The frequency of modification of Dothistroma pine needle blight severity by fungi within the native range. For. Ecol. Manag. 2015, 337, 153–160. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, J.; Liang, X.; Zhan, G.; Jiang, S.; Kang, Z. Identification of a novel Alternaria alternata strain able to hyperparasitize Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust. Front. Microbiol. 2017, 8, 7. [Google Scholar]
- Fares, S.; Loreto, F.; Kleist, E.; Wildt, J. Stomatal uptake and stomatal deposition of ozone in isoprene and monoterpene emitting plants. Plant Biol. 2007, 9, e69–e78. [Google Scholar] [CrossRef]
- Junker, R.R.; Tholl, D. Volatile organic compound mediated interactions at the plant-microbe interface. J. Chem. Ecol. 2013, 39, 810–825. [Google Scholar] [CrossRef]
- Mills, G.; Harmens, H.; Wagg, S.; Sharps, K.; Hayes, F.; Fowler, D.; Sutton, M.; Davies, B. Ozone impacts on vegetation in a nitrogen enriched and changing climate. Environ. Pollut. 2016, 208, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Yin, R.B.; Shang, B.; Agathokleous, E.; Zhou, H.M.; Feng, Z.Z. Interactive effects of ozone exposure and nitrogen addition on tree root traits and biomass allocation pattern: An experimental case study and a literature meta-analysis. Sci. Total Environ. 2020, 710, 136379. [Google Scholar] [CrossRef]
- Gopal, M.; Gupta, A. Microbiome selection could spur next-generation plant breeding strategies. Front. Microbiol. 2016, 7, 1971. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, Z.Z.; Li, X.W.; Ping, Q.; Yuan, X.Y.; Agathokleous, E.; Feng, Z.Z. Interactive effects of ozone exposure and nitrogen addition on the rhizosphere bacterial community of poplar saplings. Sci. Total Environ. 2021, 754, 142134. [Google Scholar] [CrossRef]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. Microbiome 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F., Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.; Pieterse, C.M. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, T.; Yuan, J.; He, X.; Lin, Y.; Huang, Q.; Shen, Q. Enrichment of beneficial cucumber rhizosphere microbes mediated by organic acid secretion. Hortic. Res. 2020, 7, 154. [Google Scholar] [CrossRef]
- Miranda, M.; Ralph, S.G.; Mellway, R.; White, R.; Heath, M.C.; Bohlmann, J.; Constabel, P.C. The transcriptional response of hybrid poplar (Populus trichocarpa × P. deltoids) to infection by Melampsora medusae leaf rust involves induction of flavonoid pathway genes leading to the accumulation of proanthocyanidins. Mol. Plant Microbe Interact. 2007, 20, 816–831. [Google Scholar] [CrossRef] [Green Version]
- Ullah, C.; Unsicker, S.B.; Fellenberg, C.; Constabel, P.C.; Schmidt, A.; Gershenzon, J.; Hammerbacher, A. Flavan-3-ols are an effective chemical defense against rust infection. Plant Physiol. 2017, 175, 1560–1578. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. Microbes drive evolution of animals and plants: The hologenome concept. MBio 2016, 7, e01395-15. [Google Scholar] [CrossRef] [Green Version]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling interactions in the microbiome: A network perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef]
- Connor, N.; Barberán, A.; Clauset, A. Using null models to infer microbial co- occurrence networks. PLoS ONE 2017, 12, e0176751. [Google Scholar] [CrossRef] [Green Version]
- Marasco, R.; Rolli, E.; Fusi, M.; Michoud, G.; Daffonchio, D. Grapevine rootstocks shape underground bacterial microbiome and networking but not potential functionality. Microbiome 2018, 6, 3. [Google Scholar] [CrossRef]
- Ahmadi-Rad, S.; Gholamhoseini, M.; Ghalavand, A.; Asgharzadeh, A.; Dolatabadian, A. Foliar application of nitrogen fixing bacteria increases growth and yield of canola grown under different nitrogen regimes. Rhiz 2016, 2, 34–37. [Google Scholar] [CrossRef]
- Trivedi, P.; Duan, Y.; Wang, N. Huanglongbing, a systemic disease, restructures the bacterial community associated with citrus roots. Appl. Environ. Microbiol. 2010, 76, 3427–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial interkingdom interactions in roots promote Arabidopsis survival. Cell 2018, 175, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.M.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | Clone ‘107’ | Clone ‘546’ | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clone | N | O3 | Mlp | N | O3 | Mlp | N × O3 | N × O3 × Mlp | N | O3 | Mlp | N × O3 | N × O3 × Mlp | ||
| Bacterial community | F | 6.661 | 1.517 | 0.288 | 2.702 | 0.377 | 1.295 | 0.883 | 1.374 | 1.364 | 1.065 | 0.998 | 3.016 | 1.799 | 0.717 |
| p | 0.001 | 0.195 | 0.973 | 0.023 | 0.885 | 0.254 | 0.459 | 0.238 | 0.189 | 0.388 | 0.410 | 0.013 | 0.069 | 0.644 | |
| Fungal community | F | 7.656 | 0.776 | 3.684 | 5.693 | 1.081 | 4.494 | 2.756 | 1.410 | 0.534 | 0.846 | 0.337 | 6.223 | 1.013 | 0.435 |
| p | 0.002 | 0.422 | 0.037 | 0.008 | 0.312 | 0.020 | 0.085 | 0.231 | 0.602 | 0.406 | 0.762 | 0.003 | 0.347 | 0.587 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, S.; Zhang, Y.; Tian, C.; Duplessis, S.; Zhang, N. Elevated Ozone Concentration and Nitrogen Addition Increase Poplar Rust Severity by Shifting the Phyllosphere Microbial Community. J. Fungi 2022, 8, 523. https://doi.org/10.3390/jof8050523
Tao S, Zhang Y, Tian C, Duplessis S, Zhang N. Elevated Ozone Concentration and Nitrogen Addition Increase Poplar Rust Severity by Shifting the Phyllosphere Microbial Community. Journal of Fungi. 2022; 8(5):523. https://doi.org/10.3390/jof8050523
Chicago/Turabian StyleTao, Siqi, Yunxia Zhang, Chengming Tian, Sébastien Duplessis, and Naili Zhang. 2022. "Elevated Ozone Concentration and Nitrogen Addition Increase Poplar Rust Severity by Shifting the Phyllosphere Microbial Community" Journal of Fungi 8, no. 5: 523. https://doi.org/10.3390/jof8050523
APA StyleTao, S., Zhang, Y., Tian, C., Duplessis, S., & Zhang, N. (2022). Elevated Ozone Concentration and Nitrogen Addition Increase Poplar Rust Severity by Shifting the Phyllosphere Microbial Community. Journal of Fungi, 8(5), 523. https://doi.org/10.3390/jof8050523