
New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, Fungal Isolation and Morphological Observation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analysis
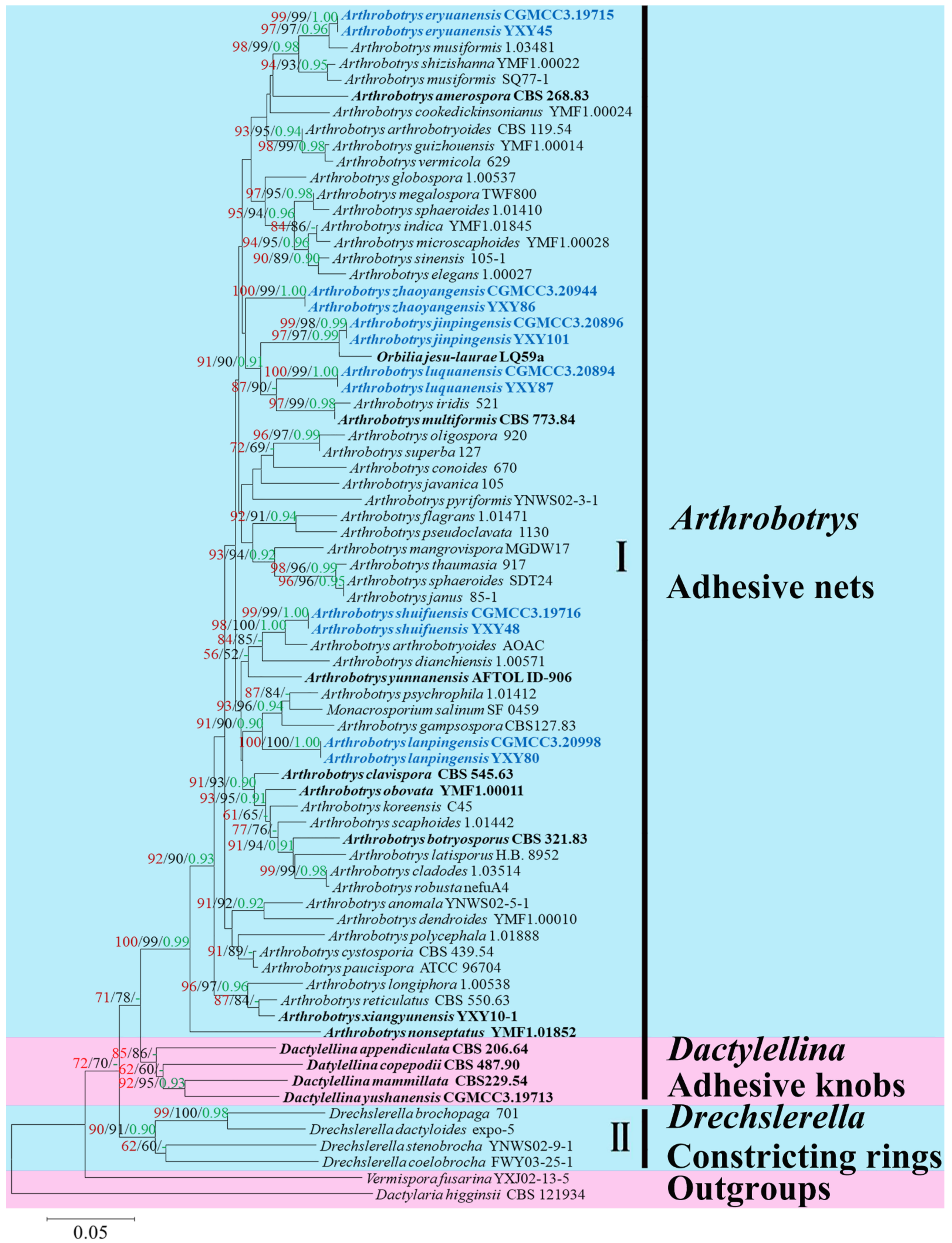
3. Results
3.1. Phylogenetic Analysis
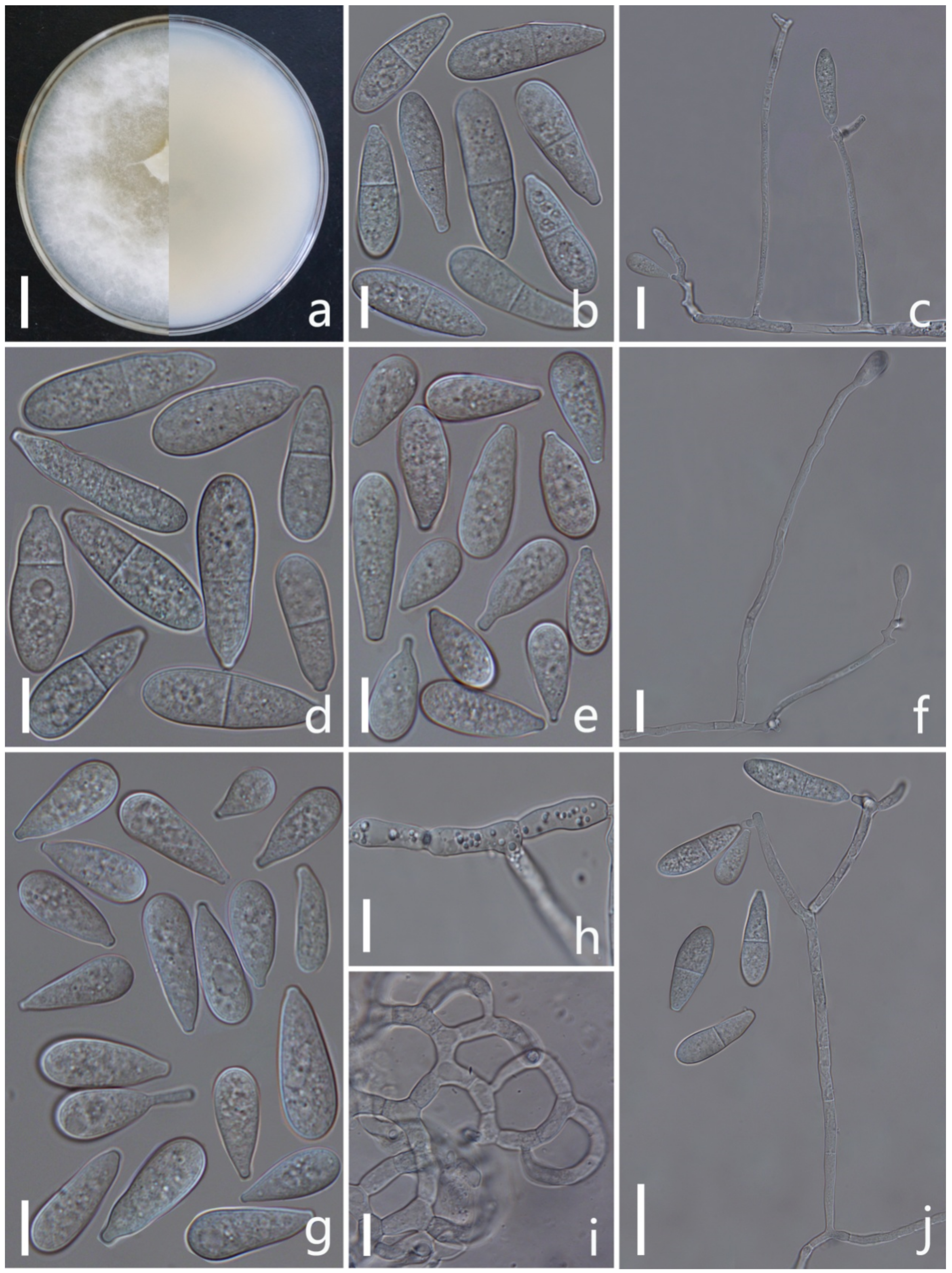
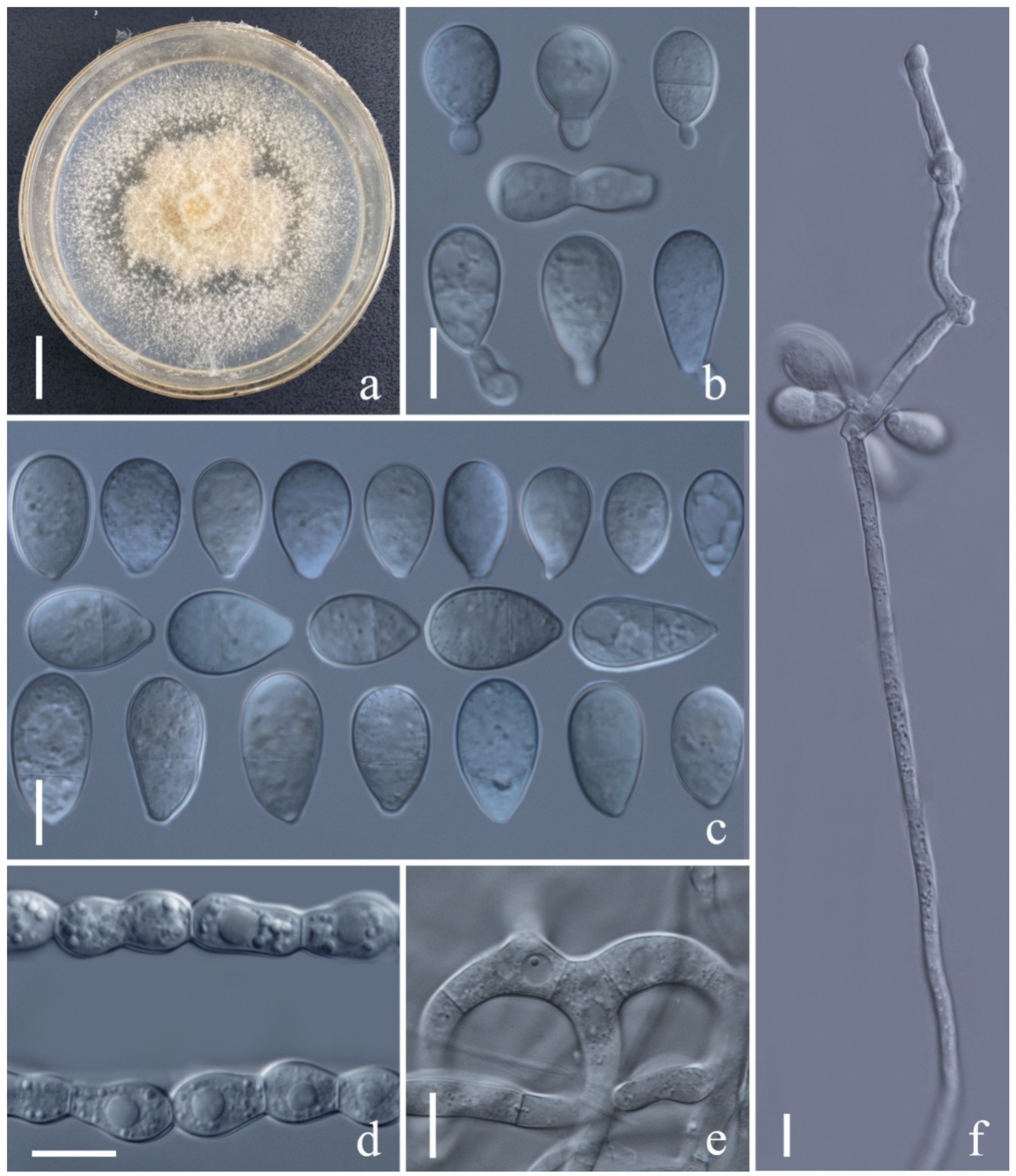
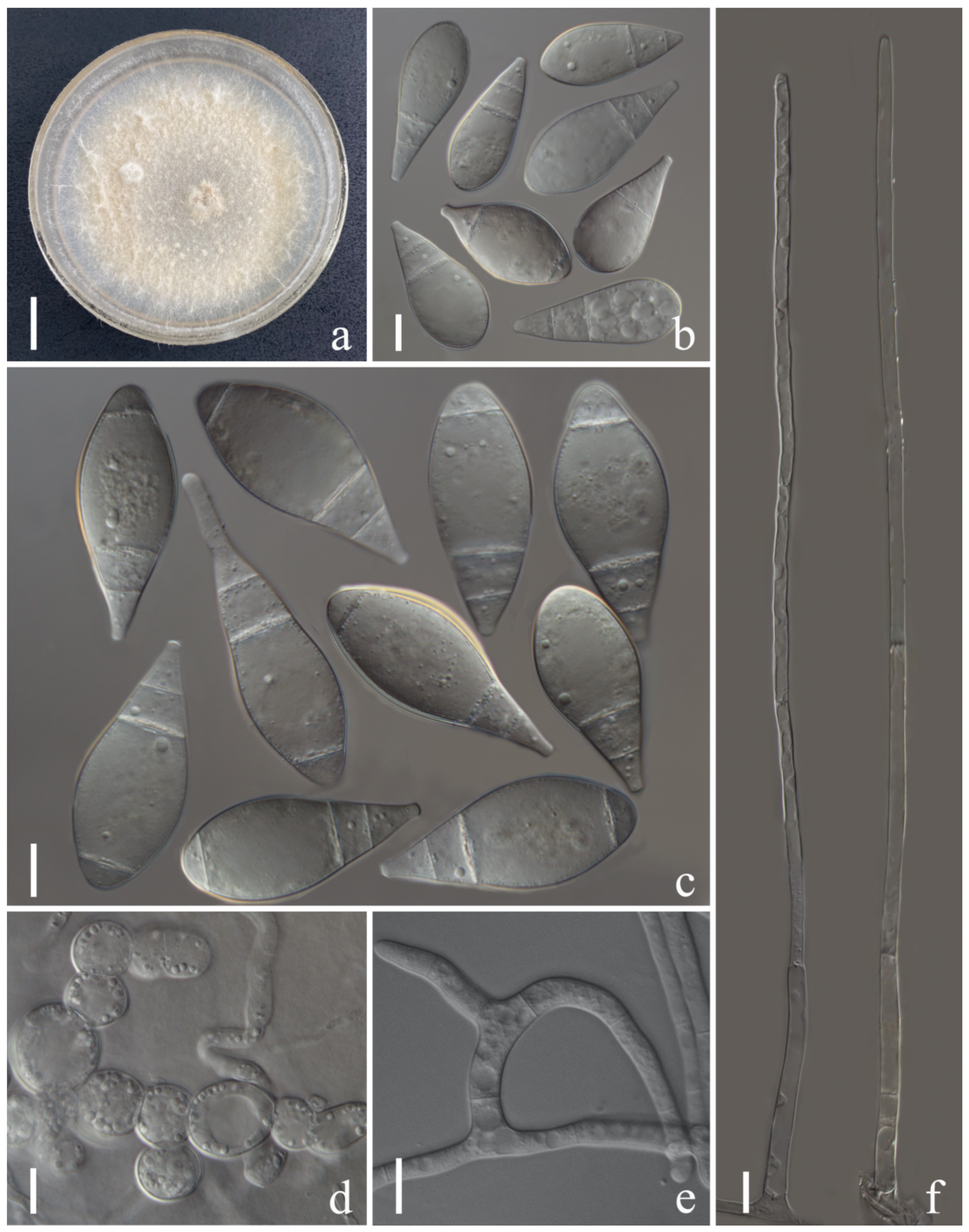
3.2. Taxonomy
3.3. Key to Known Species of Arthrobotrys
- 1.
- Conidia 0–1-septate……………………………………………………………………………2
- 1.
- Conidia multi-septate…………………………………………………………………………30
- 2.
- Conidia mostly aseptate………………………………………………………………………3
- 2.
- Conidia mostly 1-septate………………………………………………………………………6
- 3.
- Conidiophores branched near apex, producing a node at each branch, or producing several separate nodes by repeated elongation; conidia ovate, with a papilliform bulge at the base………………………………………………………………A. botryospora
- 3.
- Conidiophores unbranched……………………………………………………………………4
- 4.
- Conidiophores with a cluster short denticles at apex; conidia obovoid, 15–31 (23.5) × 10–20 (15.9) μm……………………………………………………………………A. amerospora
- 4.
- Conidiophores producing several clusters of short denticles by repeated elongation.…5
- 5.
- Conidia elongated, ellipsoid–cylindrical, 0–1-septate, mostly non-septate, 17.5–32.5 (22.6) × 2.75–7.5 (5.5) μm………………………………………………………A. yunnanensis
- 5.
- Conidia elongated, ellipsoidal, non-septate, constricted at the base, 11–16.8 × 5–6.6 μm………………………………………………………………………………A. nonseptata
- 6.
- Conidia develop on short denticles…………………………………………………………7
- 6.
- Conidia develop on nodes.……………………………………………………………………13
- 7.
- Conidia curved…………………………………………………………………………………8
- 7.
- Conidia straight………………………………………………………………………………10
- 8.
- Conidiophores unbranched, conidia in loose capitate arrangement at apex; conidia ellipsoid, mostly curved, 20–47.5 (30.9) × 7–12.5 (10.3) μm…………………A. musiformis
- 8.
- Conidiophores branched, producing several clusters short denticles by repeated elongation…………………………………………………………………………………………9
- 9.
- Conidiophores simple or occasionally branched; conidia elongate-obovoid or elongate-ellipsoidal, 1-septate, straight or curved, 33.5–57 × 11–15.5 µm………A. shahriari
- 9.
- Conidiophores branched; macroconidia 1-septate, straight or slightly curved, 18–44.5 (28.4) × 5–11.5 (8.7) µm, microconidia aseptate…………………………A. eryuanensis
- 10.
- Conidiophores producing short denticles by repeated elongation; conidia 1-septate near the base, obpyriform, sometimes constricted at the septum, 24–32.5 × 12.5–20 µm…………………………………………………………………………………A. perpasta
- 10.
- Conidiophores with clustered short denticles at apex; conidia in loose capitate arrangement at apex…………………………………………………………………………11
- 11.
- Conidia clavate, 1-septate at median or submedian, slightly constricted at the septum, 20–37.5 (27.9) × 7.5–10 (8.8) µm…………………………………A. javanica
- 11.
- Conidia obovoid or obpyriform…………………………………………………………12
- 12.
- Conidia obovoid, 1-septate near the base, apical cell much larger, smaller at basal cell, 28.5–32 (30) × 18–20.5 (20) µm…………………………………………………A. obovata
- 12.
- Conidia obpyriform, 1-septate at submedian, slightly constricted at the septum, 21.4–26.9 × 11.6–15.6 µm……………………………………………A. koreensis
- 13.
- Conidia develop on short denticles or obscure nodes of conidiophores…………14
- 13.
- Conidia develop in clusters on swollen nodes of conidiophores…………………17
- 14.
- Conidiophores branched, producing short denticles by repeated elongation; conidia obovate, elongate–obovate, 22.5–32 × 11–22.5 µm…………………………A. chazarica
- 14.
- Conidiophores unbranched; conidia clavate or pyriform…………………………15
- 15.
- Conidia develop on apical conidiophores, conidia clavate, 0 or 1-septate, constricted at the base, 30–45 × 8–11 µm……………………………… A. pseudoclavata
- 15.
- Conidia pyriform, 1-septate near the basal, apical cell much larger, smaller at basal cell; conidiophores producing several short denticles by repeated elongation………16
- 16.
- Conidia perceptibly constricted at the septum, 25–33.8 × 12.5–16.3 µm…A. paucispora
- 16.
- Conidia non-constricted, 25–35 × 18–24 µm………………………………A. cystosporia
- 17.
- Conidiophores branched………………………………………………………………18
- 17.
- Conidiophores unbranched………………………………………………………………24
- 18.
- Conidia 1-septate at median………………………………………………………………19
- 18.
- Conidia 1-septate at submedian…………………………………………………………21
- 19.
- Conidia elongate–elliptical or cylindrical, 7.5–27.5 (15.8) × 5–10.5 (6.6) µm…A. superba
- 19.
- Conidia short elliptical to oblong or capsule-shaped……………………………………20
- 20.
- Conidiophores occasionally branched, with distinct continuous swollen apical nodes; conidia ellipsoidal, 20–22 × 9–10 µm…………………………………A. arthrobotryoides
- 20.
- Conidiophores usually branched, bearing conidia on slightly swollen nodes; conidia capsule-shaped, 17–36 (27.2) × 5–12.5 (8.2) µm…………………………A. shuifuensis
- 21.
- Conidiophores bearing conidia on apical nodes; conidia oblong–pyriform, 20–27.5 (24.4) × 7.5–12.5 (10.8) µm…………………………………………………………A. robusta
- 21.
- Conidiophores producing several separate nodes by repeated elongation…………22
- 22.
- Conidia ellipsoid, elongate–obovate, 10–20 (17.5) × 5–8 (6.2) µm……………A. cladodes
- 22.
- Conidia obovoid, obpyriform, or ovoid…………………………………………………23
- 23.
- Conidia subglobose or elliptical, 14.8–21.5 (18.3) × 10.1–16.3 (13.5) µm…… A. latispora
- 23.
- Conidia obovoid or obpyriform, 1 septum at submedian, slightly constricted at the septum, 14–26 × 7.5–13 µm………………………………………………Orbilia jesu-laurae
- 24.
- Condia develop on apical node of conidiophores………………………………………25
- 24.
- Conidiophores producing several separate nodes by repeated elongation…………27
- 25.
- Conidia non-constricted at the septum, obconical or ellipsoidal, 25–50 × 10–15 µm…………………………………………………………………………………A. flagrans
- 25.
- Conidia obconical or pyriform, constricted at the septum……………………………26
- 26.
- Conidia larger size, constricted at septum, 21–42 (30.5) × 8–15 (12.7) µm……………………………………………………………………………A. apscheronica
- 26.
- Conidia small size, perceptibly constricted at the septum, 15–37.5 (28.4) × 7.5–14.5 (11.8) µm…………………………………………………………………………A. conoides
- 27.
- Conidia 1-septate at median……………………………………………………………28
- 27.
- Conidia 1-septate at submedian………………………………………………………….29
- 28.
- Conidiophores producing continuously expanded node or several separate nodes by repeated elongation; conidia cylindric, long ellipsoid, larger size, 13–22 × 3–7 µm…………………………………………………………………………………A. anomala
- 28.
- Conidiophores producing several slightly swollen nodes by repeated elongation; conidia ovate, oblong, cylindric, smaller size, 10–20 (14.6) × 2.5–5 (4) µm…A. dendroides
- 29.
- Conidia obpyriform or drop-shaped, some with a bud-like projection at the base, smaller size, 11.2–26.4(18.6) × 6.6–14.4(10.8) µm………A. jinpingensis
- 29.
- Conidia pyriform or obovoid, slightly constricted at the septum, larger size, 17–35 (23) × 8.5–16 (12) µm…………………………………………………………A. oligospora
- 30.
- Conidia without largest cell, with several septa, uniformly distributed among conidial cells ……………………………………………………………………………………31
- 30.
- Conidia with largest cell…………………………………………………………………37
- 31.
- Y-shaped conidia develop on conidiophores………………………………………A. iridis
- 31.
- Conidia other type, never Y-shaped…………………………………………………32
- 32.
- Conidiophores branched……………………………………………………………….33
- 32.
- Conidiophores unbranched………………………………………………………………34
- 33.
- Macroconidia spindle-shape or clavate, with 1–7-septate, mostly 2–5, 37.5–100 (70) × 10–17.5 (14.3) µm, microconidia spindle-shape, 0 or 1-septate…………A. dianchiensis
- 33.
- Conidia elongate–ovate to elongate–doliform or ellipsoidal, with 1–3-septate, 28.5–56 × 11.5–22.5 µm……………………………………………………….……A. tabrizica
- 34.
- Conidia bearing on apical conidiophores………………………………………………35
- 34.
- Conidiophores producing several cluster conidia by repeated elongation…………36
- 35.
- Several conidia develop on apical conidiophores, macroconidia elongate-fusiform, clavate, 4–12-septate; microconidia clavate, cylindric–clavate, 0 or 1-septate………………………………………………………………………A. multiformis
- 35.
- Conidiophores bearing single conidium; conidia clavate, sometimes slightly curved, 2–9-septate, 22.5–73.8 (50.6) × 5–10 (6.6) µm………………………A. shizishanna
- 36.
- Conidiophores with inconspicuous short denticles; macroconidia fusoid-shaped, curved, 2–4-septate, mostly 3–4, 30–50 (45.1) × 8–16.5 (12.2) µm, microconidia ellispsoid, slightly curved, 1 or 2-septate……………………………………A. polycephala
- 36.
- Conidiophores producing several short denticles by repeated elongation; conidia elongate–pyriform, 1–3-septate, mostly 2 or 3, 17–38 × 6.5–11.5 µm………A. pyriformi
- 37.
- Conidiophores branched…………………………………………………………………38
- 37.
- Conidiophores unbranched……………………………………………………….….……43
- 38.
- Conidiophores bear a single conidium………………………………………………39
- 38.
- Conidiophores bear several conidia…………………………………………………40
- 39.
- Conidia globose or obpyriform, 1–2-septate, 25–37.5 × 15–22.5 µm……A. globospora
- 39.
- Conidia subspherical or obovoid or subfusiform, 1–3-septate, 23.5–30 (27.6) × 17–25 (20) µm……………………………………………………………………………A. sinensis
- 40.
- Conidia in capitate arrangement at apex of conidiophores…………………………41
- 40.
- Conidia in non-capitate arrangement on conidiophores……………………………42
- 41.
- Conidia obovoid or ellipsoidal, 1–4-septate, mostly 1, 18–36 (28.1) × 12–20 (15.3) µm……………………………………………………………………………A. azerbaijanica
- 41.
- Conidia pyriform, 1–2-septate, mostly 1, 7.5–22.5 (15.8) × 5–10 (6.6) µm…A. oviformis
- 42.
- Conidiophores bearing 1 conidium, sometimes 2 conidia; conidia elliptic, top-shaped, 0–2-septate, 17.5–30 (23.2) × 12.5–20 (14.8) µm…………………………………A. indica
- 42.
- Conidiophores bearing several conidia; macroconidia subfusiform, 2–4-septate, 40–65 (52) × 17–23 (20) µm, microconidia obovoid, aseptate ………A. oudemansii
- 43.
- Conidiophores bear several conidia ………………………………………………44
- 43.
- Conidiophores bear a single conidium ………………………………………………52
- 44.
- Conidiophores bear several conidia near apex by repeated elongation…………45
- 44.
- Conidiophores bear several conidia at apex…………………47
- 45.
- Conidia elongate–ellipsoidal to broadly fusiform, 1–3-septate, mostly1 or 2, 25–50 × 17.5–25 µm……………………………………………………………………A. vermicola
- 45.
- Conidiophores producing denticles by repeated elongation, conidia fusiform, elongate–fusoid or clavate ……………………………………………………………………46
- 46.
- Conidia variable in shape, broadly turbinate to elongate–fusoid, ellipsoidal, fusiform, clavate, 1–3-septate, mostly 2, 25–50 (38.9) × 12–24 (17.3) µm…………A. mangrovispora
- 46.
- Conidia fusiform, sometimes slightly curved, 1–6-septate, mostly 2–3, 36.6–79.3 (57) × 11–17.5 (14) µm…………………………………………………………………A. scaphoides
- 47.
- Conidia spindle-shaped, curved,1–4-septate, 25–76 × 7–16 µm………A. gampsospora
- 47.
- Conidia straight……………………………………………………………………………48
- 48.
- Conidia 0–3-septate, mostly 1 or 2………………………………………………………49
- 48.
- Conidia 1–4-septate, mostly 3 or 4………………………………………………………50
- 49.
- Conidia cymbiform or fusiform, mostly 2-septate, 22.5–45 (27.2) × 10–20 (13.9) µm…………………………………………………………………………A. microscaphoides
- 49.
- Conidia pyriform, clavate, mostly 1 or 2 septate, 25–40 (17.5) × 7.5–19 (15.4) µm………………………………………………………………………………A. clavispora
- 50.
- Conidia small, subfusiform, 1–4-septate, mostly 3, 30–60 (36.2) × 15–30(20.2) µm………………………………………………………………………………A. thaumasia
- 50.
- Conidia larger, ellipsoidal, fusoid–ellipsoidal……………………………………51
- 51.
- Conidia ellipsoidal, fusoid–ellipsoidal, 2–4-septate, mostly 4, 46–70 (62.3) × 21–29 (24.7) µm……………………………………………………………………………A. psychrophila
- 51.
- Conidia fusiform, elongate–ellipsoidal or obovoid, 2–4-septate, mostly 3 or 4, 40–75 × 18–35 µm……………………………………………………………………A. megalospora
- 52.
- Conidia spindle-shaped, globose, 1–3-septate, mostly 2 or 3, 37–55 (49) × 17.5–35 (28) µm, microconidia ellipsoid, aseptate……………A. eudermata
- 52.
- Without microconidia……………………………………………………………………53
- 53.
- Conidia turbinate or napiform, 1–2-septate, mostly 1 near the base, the largest cell at the apex of conidia, 15–26 (22.5) × 17.5–37.5 (28.5) µm………………………A. janus
- 53.
- Conidia with more than 2 septa; the largest cell is located in the apex or center of conidia………………………………………………………………………………………54
- 54.
- The septum of the conidia is not more than 3…………………………………………55
- 54.
- Conidia 1–5-septate………………………………………………………………………58
- 55.
- Conidia clavate, obovoid, or subspherical, 0–3-septate……………………………56
- 55.
- Conidia fusiform, 2–3-septate……………………………………………………………57
- 56.
- Conidia clavate or obovoid, 1–3-septate, mostly 2–3, 30–52.5 (42) × 15–22.5 (17.6) µm………………………A. cookedickinson
- 56.
- Conidia subspherical or obovoid, 0–3-septate, mostly 2–3, 20–40 (32) × 17–25 (20.4) µm………………………………………………………………………………A. sphaeroides
- 57.
- Conidia globose or subfusiform, 2–3-septate, 27–47.5 (32.2) × 17.5–27.5 (22) µm………………………………………………………………………………A. rutgeriense
- 57.
- Conidia spindle-shaped, fusiform or ellipsoidal, 2–3-septate, 32.5–47.5 (41) × 12.5–17.5 (15.5) µm………………………………………………………………………A. fusiformis
- 58.
- Conidia 1–5-septate, mostly 3 or 4…………………………………………………….59
- 58.
- Conidia 1–4-septate, mostly 2 or 3………………………………………………………61
- 59.
- Conidia variable in shape, obpyriform, broadly turbinate, subfusiform, elongate-fusoid or clavate, 1–5-septate, 27–72 (55.8) × 14.5–28.5 (21.9) µm…A. xiangyunensis
- 59.
- Conidia ellipsoid, obpyriform or subfusiform, 2–5-septate, mostly 3 or 4……………60
- 60.
- Conidia ellipsoid, fusiform, 50–65 × 20–25 µm…………………………………A. reticulata
- 60.
- Conidia obpyriform or subfusiform, 40–90 (54) × 15–27.5 (18) µm………… A. longiphora
- 61.
- Conidia mostly 2-septate……………………………………………………………………62
- 61.
- Conidia mostly 3-septate……………………………………………………………………63
- 62.
- Conidia obovate, obpyriform or drop-shaped; the distal cell is much smaller, the largest cell usually at the apex, 28–53.5 (40.9) × 17–32.5 (26.3) µm……………… A. luquanensis
- 62.
- Conidia obpyriform or subfusiform, the largest cell usually at the centre, 30.5–71.5 (52.7) × 18.5–28.5 (23.9) µm……………………………………………A. guizhouensis
- 63.
- Conidia, subglobose, obovoid to obpyriform, 25.5–52 (35.4) × 14–32 (22.9) µm……………………………………………………………………………A. zhaoyangensis
- 63.
- Conidia fusiform or ellipsoid…………………………………64
- 64.
- Conidia fusiform to ellipsoid, 32.5–52.5 × 12.5–17.5 µm………………………… A. salina
- 64.
- Conidia mostly subfusiform, 31.1–55.2 (45.4) × 13.5–24.3 (19.7) µm………A. lanpingensis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaffee, B.; Tedford, E.; Muldoon, A. Tests for density-dependent parasitism of nematodes by nematode-trapping and endoparasitic fungi. Biol. Control 1993, 3, 329–336. [Google Scholar] [CrossRef]
- Linford, M.; Yap, F.; Oliveira, J.M. Reduction of soil populations of the root-knot nematode during decomposition of organic matter. Soil Sci. 1938, 45, 127–142. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Li, T.F.; Liu, X.Z. Biology of Nematophagous Fungi; China Science & Technology Press: Beijing, China, 2001. [Google Scholar]
- Zhang, K.Q.; Hyde, K.D. Nematode-Trapping Fungi; Springer Science & Business: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Li, T.F.; Zhang, K.Q.; Liu, X.Z. Taxonomy of Nematophagous Fungi; Chinese Scientific and Technological Publication: Beijing, China, 2000. [Google Scholar]
- Corda, A.K.J. Pracht-Flora Europaeischer Schimmelbildungen; G. Fleischer: Leipzig, Germany, 1839; p. 43. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi; CAB International: Oxon, UK, 2008. [Google Scholar]
- Yang, E.; Xu, L.L.; Yang, Y.; Zhang, X.Y.; Xiang, M.C.; Wang, C.S.; An, Z.Q.; Liu, X.Z. Origin and evolution of carnivorism in the Ascomycota (fungi). Proc. Natl. Acad. Sci. USA 2012, 109, 10960–10965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikawa, M. Ultrastructural studies on zygomycotan fungi in the Zoopagaceae and Cochlonemataceae. Mycoscience 2011, 52, 83–90. [Google Scholar] [CrossRef]
- Poloczek, E.; Webster, J. Conidial traps in Nematoctonus (nematophagous Basidiomycetes). Nova Hedwig. 1994, 59, 201–205. [Google Scholar]
- Durschnerpelz, U.U. Traps of Nematoctonus leiosporus: An unusual feature of an endoparasitic nematophagous fungus. Trans Br. Mycol. Soc. 1987, 88, 129–130. [Google Scholar] [CrossRef]
- Thorn, R.G.; Moncalvo, J.M.; Reddy, C.A.; Vilgalys, R. Phylogenetic analyses and the distribution of nematophagy support a monophyletic Pleurotaceae within the polyphyletic pleurotoid-lentinoid fungi. Mycologia 2000, 92, 241–252. [Google Scholar] [CrossRef]
- Swe, A.; Li, J.; Zhang, K.Q.; Pointing, S.B.; Jeewon, R.; Hyde, K.D. Nematode-trapping fungi. Curr. Res. Environ. Appl. Mycol. 2011, 1, 1–26. [Google Scholar]
- Ahrén, D.; Ursing, B.M.; Tunlid, A. Phylogeny of nematode-trapping fungi based on 18S rDNA sequences. FEMS Microbiol. Lett. 1998, 158, 179–184. [Google Scholar] [CrossRef]
- Li, Y.; Hyde, K.D.; Jeewon, R.; Cai, L.; Vijaykrishna, D.; Zhang, K. Phylogenetics and evolution of nematode-trapping fungi (Orbiliales) estimated from nuclear and protein coding genes. Mycologia 2005, 97, 1034–1046. [Google Scholar] [CrossRef]
- Jaffee, B.; Ferris, H.; Scow, K. Nematode-trapping fungi in organic and conventional cropping systems. Phytopathology 1998, 88, 344–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.R.; Su, H.Y.; Su, X.J.; Zhang, F.; Liao, G.H.; Yang, X.Y. Arthrobotrys xiangyunensis, a novel nematode-trapping taxon from a hot-spring in Yunnan Province, China. Phytotaxa 2014, 174, 89–96. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, X.J.; Monkai, J.; Li, F.T.; Liu, S.R.; Yang, X.Y.; Xiao, W.; Hyde, K.D. Two new species of nematode-trapping fungi (Dactylellina, Orbiliaceae) from burned forest in Yunnan, China. Phytotaxa 2020, 452, 65–74. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Mo, M.H. Flora Fungorum Sinicorum: Arthrobotrys et Gengra Cetera Cognata; Science Press: Beijing, China, 2006; Volume 33. [Google Scholar]
- Fresenius, G. Beitrage zur Mykologie (Heft 1–2); Brönner: Frankfurt, Germany, 1852; pp. 1–80. [Google Scholar]
- Zopf, W.F. Zur Kenntniss der Infections-Krankheiten niederer Thiere und Pflanzen. Acad. Nat. 1888, 52, 314–376. [Google Scholar]
- Drechsler, C. Morphological features of some more fungi that capture and kill nematode. Acad. Sci. 1933, 23, 267–270. [Google Scholar]
- Drechsler, C. Some conidial hyphomycetes destructive to terricolous amoebe. Mycologia 1935, 27, 6–40. [Google Scholar] [CrossRef]
- Drechsler, C. A fusarium-like species of Dactylella capturing and consuming testaceous rhizopods. J. Wash. Acad. Sci. 1936, 26, 397–404. [Google Scholar]
- Drechsler, C. Some hyphomycetes that prey on free-living terricolous nematodes. Mycologia 1937, 32, 448–470. [Google Scholar] [CrossRef]
- Drechsler, C. Several species of Dactylella and Dactylaria that capture free-living nematodes. Mycologia 1950, 42, 1–79. [Google Scholar] [CrossRef]
- Drechsler, C. Some clampless hyphomycetes predaceous on nematodes and rhizopods. Sydowia 1961, 15, 762–782. [Google Scholar]
- Drechsler, C. A slender-spored Dactylella parasitic on zoospores. Phytotholigy 1963, 53, 993–994. [Google Scholar]
- Duddington, C.L. Further records of British predaceous fungi. Trans. Brit. Mycol. Soc. 1950, 33, 3–4. [Google Scholar] [CrossRef]
- Duddington, C.L. Two new predacious hyphomycetes. Trans. Brit. Mycol. Soc. 1951, 34, 598. [Google Scholar] [CrossRef]
- Duddington, C.L. Nematode-destroying fungi in agricultural soils. Nature 1954, 168, 38–39. [Google Scholar] [CrossRef] [PubMed]
- Duddington, C.L. Notes on the technique of handling predacious fungi. Trans. Brit. Mycol. Soc. 1955, 38, 97–103. [Google Scholar] [CrossRef]
- Duddington, C.L. The Friendly Fungi: A3 Problem; Faber and Faber: London, UK, 1957. [Google Scholar] [CrossRef]
- Soprunov, F.F.; Galiulina, Z.A. Predaceous hyphomycetes from Turkmenistan soil. Mikorobiologiya 1951, 20, 489–499. [Google Scholar]
- Sherbakoff, C.D. A new fungus parasitic on nematodes. Mycologia 1933, 25, 258. [Google Scholar] [CrossRef]
- Subramanian, C.V. Revision of hyphomycetes 1. Kawaka 1977, 5, 93–98. [Google Scholar]
- Haard, K. Taxonomic studies on the genus Arthrobotrys Corda. Mycotaxon 1968, 16, 107–113. [Google Scholar] [CrossRef]
- Schenck, S.; Kendrick, W.B.; Pramer, D. A new nematode-trapping hyphomycete and a reevaluation of Dactylaria and Arthrobotrys. Can. J. Bot. 1977, 55, 977–985. [Google Scholar] [CrossRef]
- Castaner, D. Monacrosporium sphaeroides, a new nematode-destroying hyphomycete from Iowa. Am. Midl. Nat. 1968, 80, 280–283. [Google Scholar] [CrossRef]
- Chen, J.; Xu, L.L.; Liu, B.; Liu, X.Z. Taxonomy of Dactylella complex and Vermispora. I. Generic concepts based on morphology and ITS sequences data. Fungal Divers. 2007, 26, 73–83. [Google Scholar]
- Cooke, R.C.; Dickinson, C.H. Nematode-trapping species of Dactylella and Monacrosporium. Trans. Br. Mycol. Soc. 1965, 48, 621–629. [Google Scholar] [CrossRef]
- McCulloch, J.S. New species of nematophagous fungi from Queensland. Trans. Br. Mycol. Soc. 1977, 68, 173–179. [Google Scholar] [CrossRef]
- Kim, D.G.; Bae, S.G.; Shin, Y.S. Distribution of nematophagous fungi under different habitats. Korean J. Mycol. 2001, 29, 123–126. [Google Scholar]
- Mo, M.H.; Chen, W.M.; Yang, H.R.; Zhang, K.Q. Diversity and metal tolerance of nematode-trapping fungi in Pb-polluted soils. J. Microbiol. 2008, 46, 16. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, R.; Singh, K. Occurrence and colonization of nematophagous fungi in different substrates, agricultural soils and root galls. Arch. Phytopathol. Plant. Prot. 2011, 44, 1182–1195. [Google Scholar] [CrossRef]
- Su, H.Y.; Mo, M.H.; Zhang, K.Q. The ecology of nematode-trapping hyphomycetes in cattle dung from three plateau pastures. Vet. Parasitol. 2007, 144, 293–298. [Google Scholar] [CrossRef]
- Swe, A.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Diversity and abundance of nematode-trapping fungi from decaying litter in terrestrial, freshwater and mangrove habitats. Biodivers. Conserv. 2009, 18, 1695–1714. [Google Scholar] [CrossRef]
- Hastuti, L.D.S.; Berliani, K.; Mulya, M.B. Inventarization of Nematode-trapping Fungi in Terrestrial Area of Deli Serdang Regency, North Sumatera. In Proceedings of the International Conference of Science, Technology, Engineering, Environmental and Ramification Researches—ICOSTEERR, Medan, Indonesia, 30–31 August 2018. [Google Scholar] [CrossRef]
- Shepherd, A.M. Some Observations on the Distribution Andbiology of Fungi Predaceouson Nematodes. Ph.D. Thesis, University of London, London, UK, 1955. [Google Scholar]
- Fowler, M. New Zealand predaceous fungi. N. Z. J. Bot. 1970, 8, 283–302. [Google Scholar] [CrossRef] [Green Version]
- Farrell, F.; Jaffee, B.; Strong, D. The nematode-trapping fungi Arthrobotrys oligospora in soil of the Bodega marne reserve: Distribution and dependenceon nematode-parasitized moth larvae. Soil Biol. Biochem. 2006, 38, 1422–1429. [Google Scholar] [CrossRef]
- Drechsler, C. Predacious fungi. Biol. Rev. Camb. Philos. Soc. 1941, 16, 265–290. [Google Scholar] [CrossRef]
- Eren, J.; Pramer, D. The most probable number of nematode-trapping fungi in soil. Soil Sci. 1965, 99, 285. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, E.; An, Z.; Liu, X.Z. Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef] [Green Version]
- Swindell, S.R.; Plasterer, T.N. Seqman. In Sequence Data Analysis Guidebook; Swindell, S.R., Ed.; Springer: Totowa, NJ, USA, 1997; pp. 75–89. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Gronewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Rezakhani, F.; Khodaparast, S.A.; Masigol, H.; Roja-Jimenez, K.; Grossart, H.P.; Bakhshi, M. A preliminary report of aquatic hyphomycetes isolated from Anzali lagoon (Gilan province, North of Iran). Rostaniha 2019, 20, 123–143. [Google Scholar] [CrossRef]
- Liou, G.Y.; Tzean, S.S. Phylogeny of the genus Arthrobotrys and allied nematode-trapping fungi based on rDNA sequences. Mycologia 1997, 89, 876–884. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhu, H.; Zhang, K.Q. Phylogenic analysis of adhesion related genes Mad1 revealed a positive selection for the evolution of trapping devices of nematode-trapping fungi. Sci. Rep. 2016, 6, 22609. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.Y.; Kim, D.G.; Ryu, Y.H.; Zhou, X.B. Arthrobotrys koreensis, a new nematode-trapping species from Korea. Sydowia 2012, 64, 129–136. [Google Scholar]
- Swe, A.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Taxonomy and molecular phylogeny of Arthrobotrys mangrovispora, a new marine nematode-trapping fungal species. Bot. Mar. 2008, 51, 331–338. [Google Scholar] [CrossRef]
- Li, Y.; Jeewon, R.; Hyde, K.D.; Mo, M.H.; Zhang, K.Q. Two new species of nematode-trapping fungi: Relationships inferred from morphology, rDNA and protein gene sequence analyses. Mycol. Res. 2006, 110, 790–800. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O.; Beltrán-Tejera, E.; Pfister, D.H. Orbilia jesu-laurae (Ascomycota, Orbiliomycetes), a new species of neotropical nematode-trapping fungus from Puerto Rico, supported by morphology and molecular phylogenetics. Willdenowia 2020, 50, 241–251. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. Available online: http://mafft.cbrc.jp/alignment/server (accessed on 6 June 2021).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP^*, 4.0 b8; Phylogenetic Analysis Using Parsimony (and Other Methods); Sinauer: Sunderland, MA, USA, 2001. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Phylogenetic Trees Made Easy: A How-to Manual; Massachusetts: Sinauer Associates Sunderland: Sunderland, MA, USA, 2007. [Google Scholar]
- Rambaut, A. FigTree v1. 3.1. 2010. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 5 March 2021).
- Liu, X.; Zhang, K. Dactylella shizishanna sp. nov., from Shizi Mountain, China. Fungal Divers. 2003, 14, 103–107. [Google Scholar]
- Zhang, K.; Liu, X.; Cao, L. Nematophagous species of Monacrosporium from China. Mycol. Res. 1996, 100, 274–276. [Google Scholar] [CrossRef]
- Lindau, G. Die Pilze Deutschlands, Oesterreichs und der Schweiz. VIII. Abteilung: Fungi imperfecti: Hyphomycetes (erste Hälfte). Rabenhorst L. Kryptogamen-Flora. Ed 1904, 2, 1904–1907. [Google Scholar]
- Scholler, M.; Hagedorn, G.; Runner, A. A reevaluation of predatory orbiliaceous fungi. II. A new generic concept. Sydowia-Horn 1999, 51, 89–113. [Google Scholar]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosp 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Chethana, K.W.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspe, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Meier, R.; Blaimer, B.; Buenaventura, E.; Hartop, E.; von Rintelen, T.; Srivathsan, A.; Yeo, D. A re-analysis of the data in Sharkey et al.’s (2021) minimalist revision reveals that BINs do not deserve names, but BOLD Systems needs a stronger commitment to open science. Cladistics 2022, 38, 264–275. [Google Scholar] [CrossRef]
- She, R.; Zhou, X.J.; Wang, H.Q.; Zhang, F.; Yang, X.Y.; Xiao, W. Succession of soil nematode-trapping fungi following fire disturbance in forest. J. For. Res. 2020, 25, 433–438. [Google Scholar] [CrossRef]
- Serra, E.F.; Silva, A.L.; Ripoll, M.K.; Tavares, N.C.; Waller, S.B.; Osório, L.D.G.; Meireles, M.C.A. Nematode-trapping fungi in soil samples from Alegrete-RS. Sci. Anim. Health 2017, 5, 21–34. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
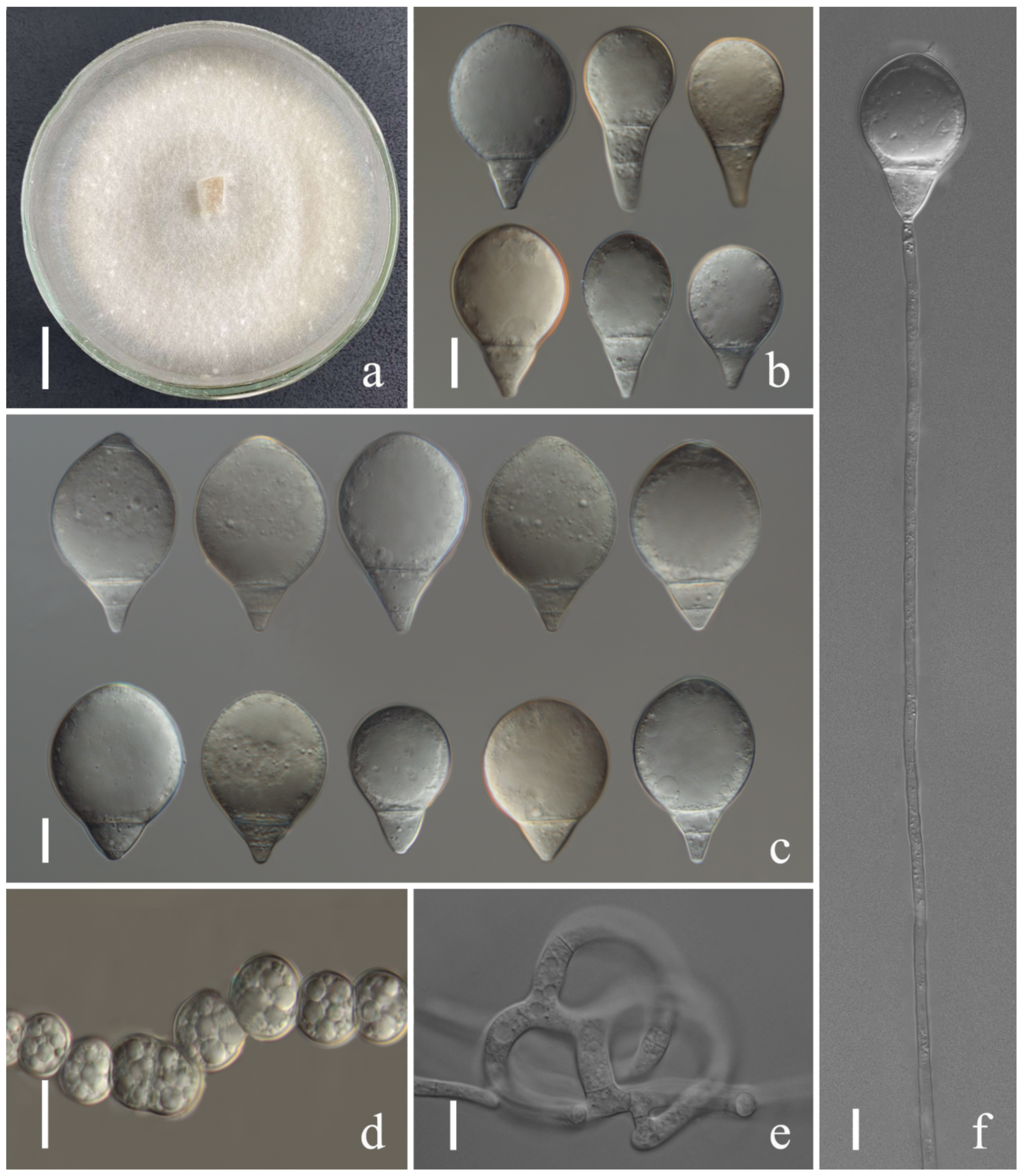{kind=link}
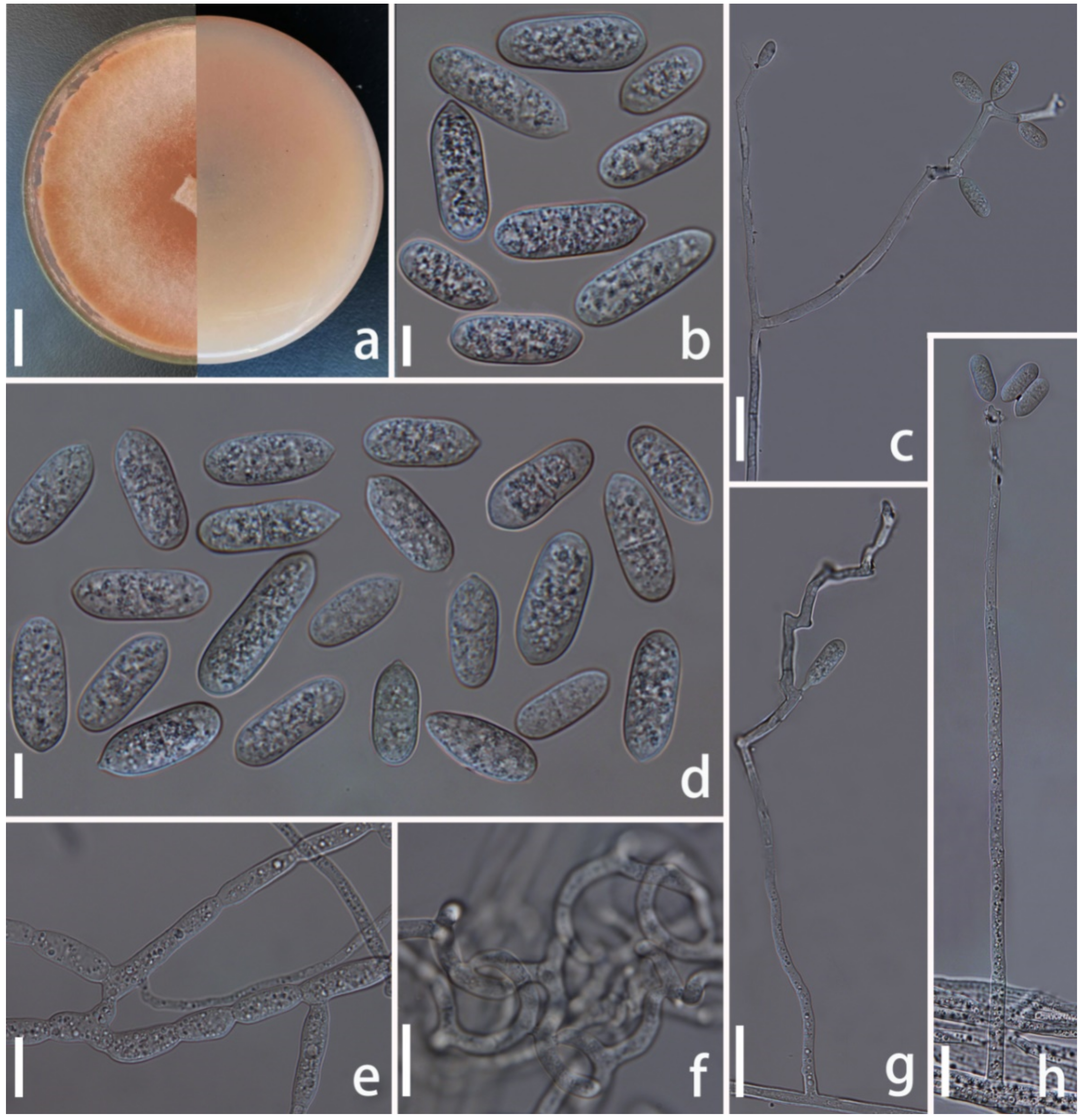{kind=link}
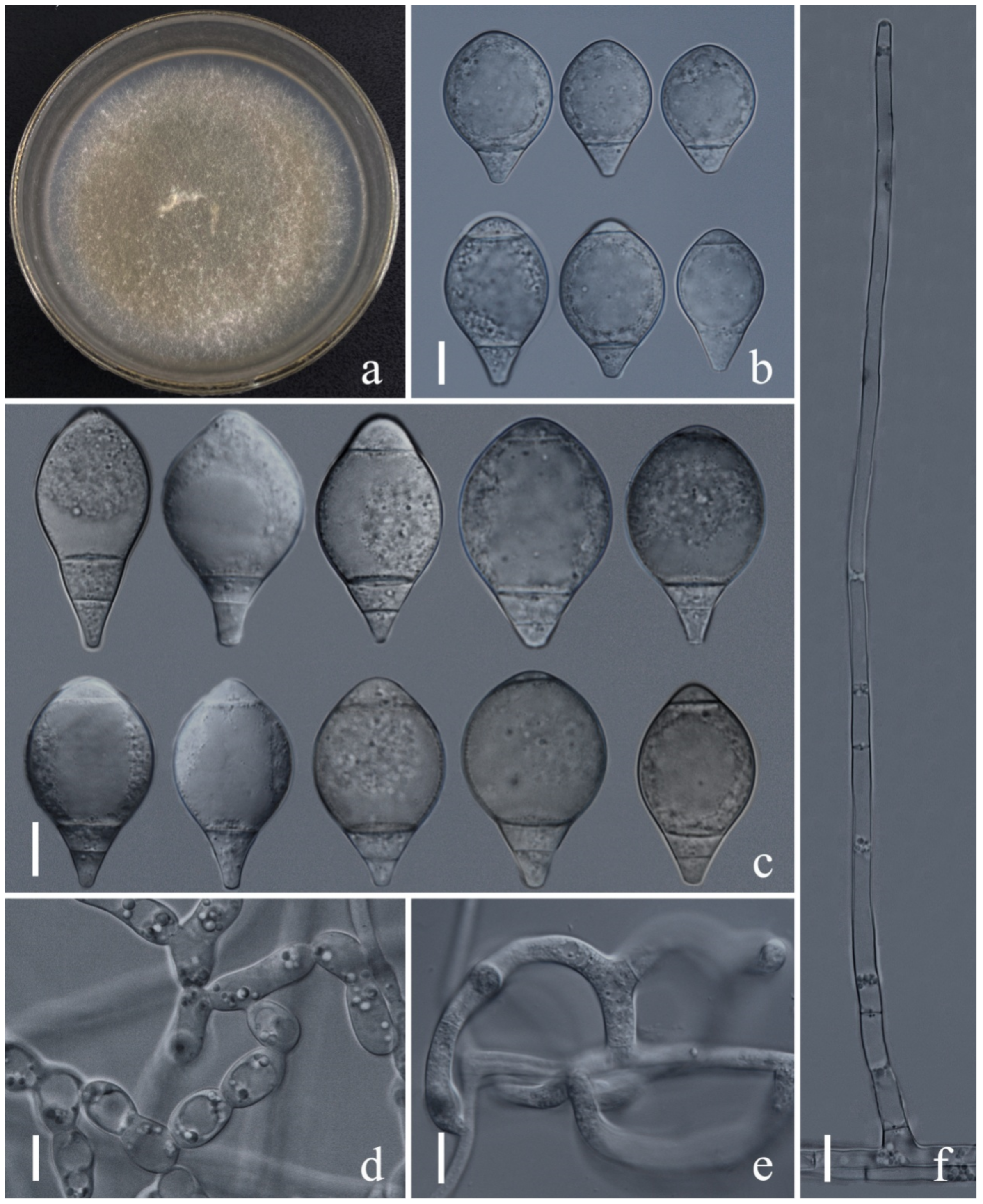{kind=link}
| Taxon | Strain Number | GenBank Accession Number | Reference | ||
|---|---|---|---|---|---|
| ITS | TEF | RPB2 | |||
| Arthrobotrys amerospora | CBS 268.83 | NR 159625 | — | — | [59] |
| Arthrobotrys anomala | YNWS02-5-1 | AY773451 | AY773393 | AY773422 | [57] |
| Arthrobotrys arthrobotryoides | CBS 119.54 | MH857262 | — | — | [59] |
| Arthrobotrys arthrobotryoides | AOAC | MF926580 | — | — | Unpublished |
| Arthrobotrys botryospora | CBS 321.83 | NR 159626 | — | — | [59] |
| Arthrobotrys cladodes | 1.03514 | MH179793 | MH179616 | MH179893 | Unpublished |
| Arthrobotrys clavispora | CBS 545.63 | MH858353 | — | — | [59] |
| Arthrobotrys conoides | 670 | AY773455 | AY773397 | AY773426 | [57] |
| Arthrobotrys cookedickinson | YMF1.00024 | MF948393 | MF948550 | MF948474 | [4] |
| Arthrobotrys cystosporia | CBS 439.54 | MH857384 | — | — | [59] |
| Arthrobotrys dendroides | YMF1.00010 | MF948388 | MF948545 | MF948469 | [4] |
| Arthrobotrys dianchiensis | 1.00571 | MH179720 | — | MH179826 | [60] |
| Arthrobotrys elegans | 1.00027 | MH179688 | — | MH179797 | Unpublished |
| Arthrobotrys eryuanensis | CGMCC3.19715 | MT612105 | OM850307 | OM850301 | This study |
| Arthrobotrys eryuanensis | YXY45 | ON808616 | ON809547 | ON809553 | This study |
| Arthrobotrys eudermata | SDT24 | AY773465 | AY773407 | AY773436 | [57] |
| Arthrobotrys flagrans | 1.01471 | MH179741 | MH179583 | MH179845 | Unpublished |
| Arthrobotrys gampsospora | CBS 127.83 | U51960 | — | — | [61] |
| Arthrobotrys globospora | 1.00537 | MH179706 | MH179562 | MH179814 | Unpublished |
| Arthrobotrys guizhouensis | YMF1.00014 | MF948390 | MF948547 | MF948471 | [4] |
| Arthrobotrys indica | YMF1.01845 | KT932086 | — | — | [62] |
| Arthrobotrys iridis | 521 | AY773452 | AY773394 | AY773423 | [57] |
| Arthrobotrys janus | Jan-85 | AY773459 | AY773401 | AY773430 | [57] |
| Arthrobotrys javanica | 105 | EU977514 | — | — | Unpublished |
| Arthrobotrys jinpingensis | CGMCC3.20896 | OM855569 | OM850311 | OM850305 | This study |
| Arthrobotrys jinpingensis | YXY101 | ON808621 | ON809552 | ON809558 | This study |
| Arthrobotrys koreensis | C45 | JF304780 | — | — | [63] |
| Arthrobotrys lanpingensis | CGMCC3.20998 | OM855566 | OM850308 | OM850302 | This study |
| Arthrobotrys lanpingensis | YXY80 | ON808618 | ON809549 | ON809555 | This study |
| Arthrobotrys latispora | H.B. 8952 | MK493125 | — | — | Unpublished |
| Arthrobotrys longiphora | 1.00538 | MH179707 | — | MH179815 | Unpublished |
| Arthrobotrys luquanensis | CGMCC3.20894 | OM855567 | OM850309 | OM850303 | This study |
| Arthrobotrys luquanensis | YXY87 | ON808619 | ON809550 | ON809556 | This study |
| Arthrobotrys mangrovispora | MGDW17 | EU573354 | — | — | [64] |
| Arthrobotrys megalospora | TWF800 | MN013995 | — | — | Unpublished |
| Arthrobotrys microscaphoides | YMF1.00028 | MF948395 | MF948552 | MF948476 | [4] |
| Arthrobotrys multiformis | CBS 773.84 | MH861834 | — | — | [59] |
| Arthrobotrys musiformis | SQ77-1 | AY773469 | AY773411 | AY773440 | [57] |
| Arthrobotrys musiformis | 1.03481 | MH179783 | MH179607 | MH179883 | Unpublished |
| Arthrobotrys nonseptata | YMF1.01852 | FJ185261 | — | — | [62] |
| Arthrobotrys obovata | YMF1.00011 | MF948389 | MF948546 | MF948470 | [4] |
| Arthrobotrys oligospora | 920 | AY773462 | AY773404 | AY773433 | [57] |
| Arthrobotrys paucispora | ATCC 96704 | EF445991 | — | — | [57] |
| Arthrobotrys polycephala | 1.01888 | MH179760 | MH179592 | MH179862 | Unpublished |
| Arthrobotrys pseudoclavata | 1130 | AY773446 | AY773388 | AY773417 | [57] |
| Arthrobotrys psychrophila | 1.01412 | MH179727 | MH179578 | MH179832 | Unpublished |
| Arthrobotrys pyriformis | YNWS02-3-1 | AY773450 | AY773392 | AY773421 | [57] |
| Arthrobotrys reticulata | CBS 550.63 | MH858355 | — | — | [59] |
| Arthrobotrys robusta | nefuA4 | MZ326655 | — | — | Unpublished |
| Arthrobotrys salina | SF 0459 | KP036623 | — | — | Unpublished |
| Arthrobotrys scaphoides | 1.01442 | MH179732 | MH179580 | MH179836 | Unpublished |
| Arthrobotrys shizishanna | YMF1.00022 | MF948392 | MF948549 | MF948473 | [4] |
| Arthrobotrys shuifuensis | CGMCC3.19716 | MT612334 | OM850306 | OM850300 | This study |
| Arthrobotrys shuifuensis | YXY48 | ON808617 | ON809548 | ON809554 | This study |
| Arthrobotrys sinensis | 105-1 | AY773445 | AY773387 | AY773416 | [57] |
| Arthrobotrys sphaeroides | 1.0141 | MH179726 | MH179577 | MH179831 | Unpublished |
| Arthrobotrys superba | 127 | EU977558 | — | — | Unpublished |
| Arthrobotrys thaumasia | 917 | AY773461 | AY773403 | AY773432 | [57] |
| Arthrobotrys vermicola | 629 | AY773454 | AY773396 | AY773425 | [57] |
| Arthrobotrys xiangyunensis | YXY10-1 | MK537299 | — | — | [17] |
| Arthrobotrys yunnanensis | AFTOL-ID 906 | DQ491512 | — | — | Unpublished |
| Arthrobotrys zhaoyangensis | CGMCC3.20944 | OM855568 | OM850310 | OM850304 | This study |
| Arthrobotrys zhaoyangensis | YXY86 | ON808620 | ON809551 | ON809557 | This study |
| Dactylaria higginsii | CBS 121934 | KM009164 | — | — | Unpublished |
| Dactylellina appendiculata | CBS 206.64 | AF106531 | DQ358227 | DQ358229 | [59] |
| Dactylellina copepodii | CBS 487.90 | U51964 | DQ999835 | DQ999816 | [61] |
| Dactylellina mammillata | CBS229.54 | AY902794 | DQ999843 | DQ999817 | [65] |
| Datylellina yushanensis | CGMCC3.19713 | MK372061 | MN915113 | MN915112 | [18] |
| Drchslerella coelobrocha | FWY03-25-1 | AY773464 | AY773406 | AY773435 | [57] |
| Drchslerella dactyloides | expo-5 | AY773463 | AY773405 | AY773434 | [57] |
| Drchslerella stenobrocha | YNWS02-9-1 | AY773460 | AY773402 | AY773431 | [57] |
| Drechslerella brochopaga | 701 | AY773456 | AY773398 | AY773427 | [57] |
| Orbilia jesu-laurae | LQ59a | MN816816 | — | — | [66] |
| Vermispora fusarina | YXJ02-13-5 | AY773447 | AY773389 | AY773418 | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Boonmee, S.; Bhat, J.D.; Xiao, W.; Yang, X.-Y. New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China. J. Fungi 2022, 8, 671. https://doi.org/10.3390/jof8070671
Zhang F, Boonmee S, Bhat JD, Xiao W, Yang X-Y. New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China. Journal of Fungi. 2022; 8(7):671. https://doi.org/10.3390/jof8070671
Chicago/Turabian StyleZhang, Fa, Saranyaphat Boonmee, Jayarama D. Bhat, Wen Xiao, and Xiao-Yan Yang. 2022. "New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China" Journal of Fungi 8, no. 7: 671. https://doi.org/10.3390/jof8070671
APA StyleZhang, F., Boonmee, S., Bhat, J. D., Xiao, W., & Yang, X. -Y. (2022). New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China. Journal of Fungi, 8(7), 671. https://doi.org/10.3390/jof8070671