
Metabolic Diversity of Xylariaceous Fungi Associated with Leaf Litter Decomposition

Abstract
:1. Introduction
2. Materials and Methods
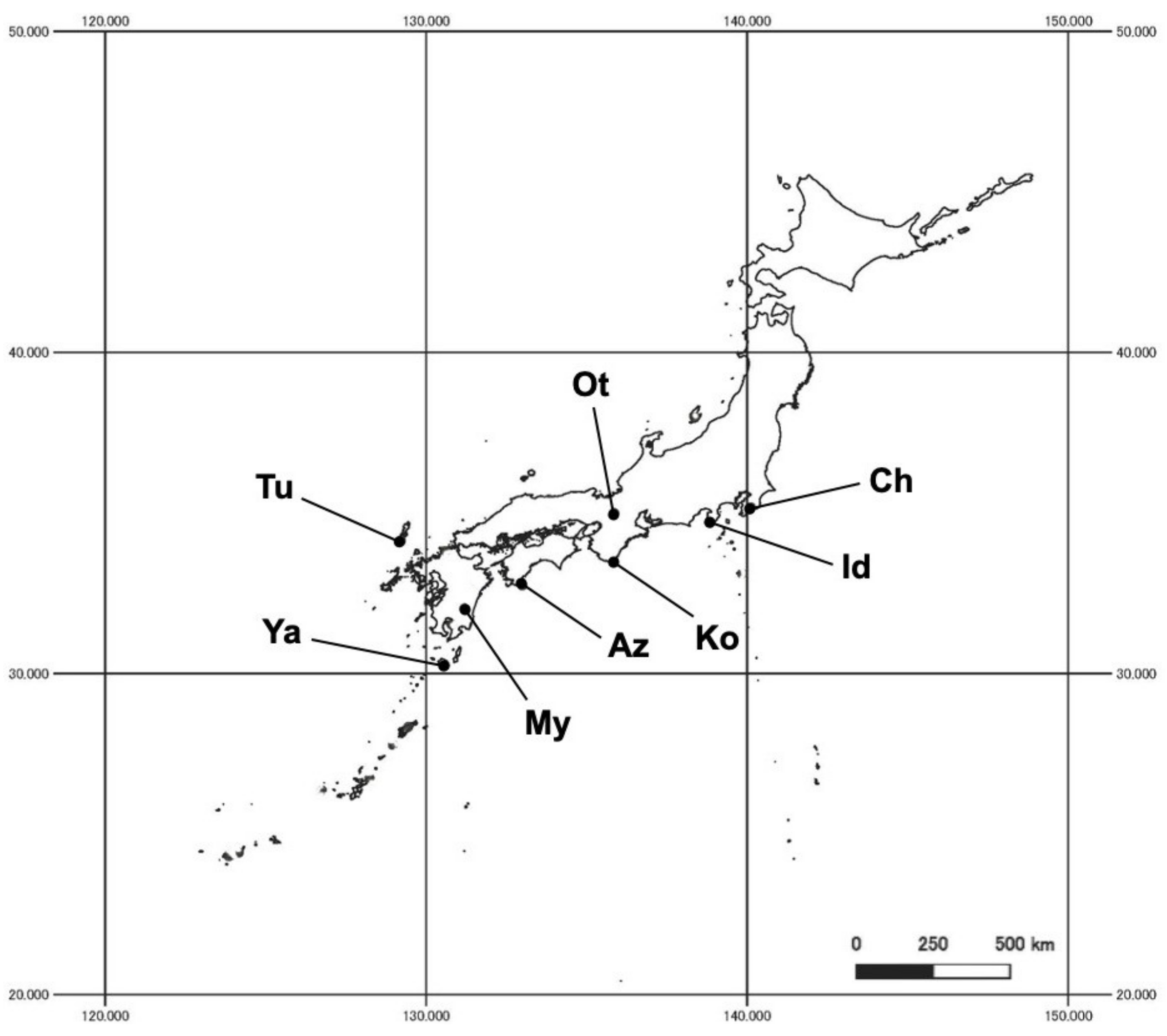
2.1. Study Site and Fungal Isolation
2.2. Molecular Analysis
2.3. Phylogenetic Analysis
2.4. Carbon Source Utilization
2.5. Statistical Analyses
3. Results
3.1. Taxonomic Assignment
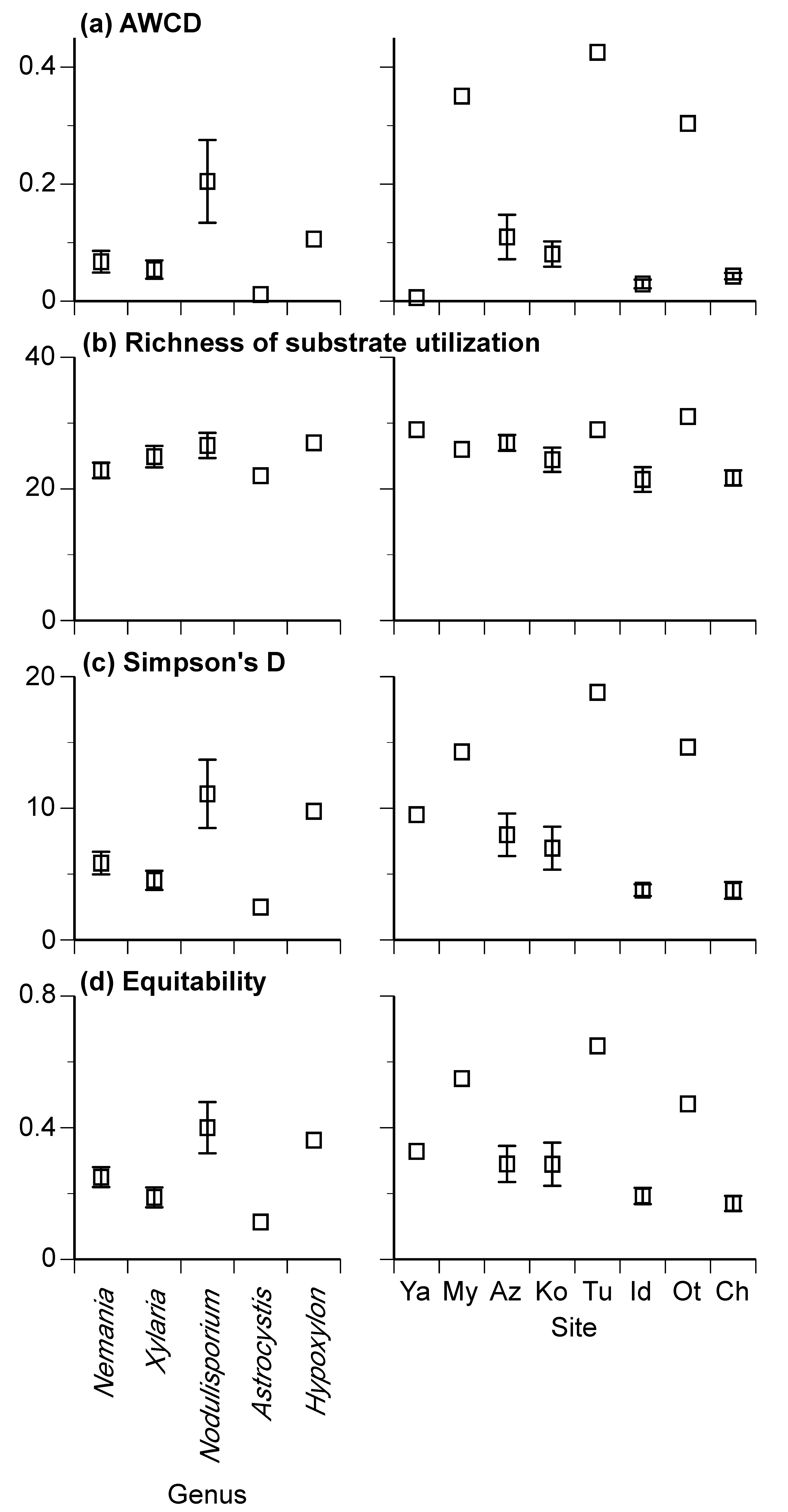
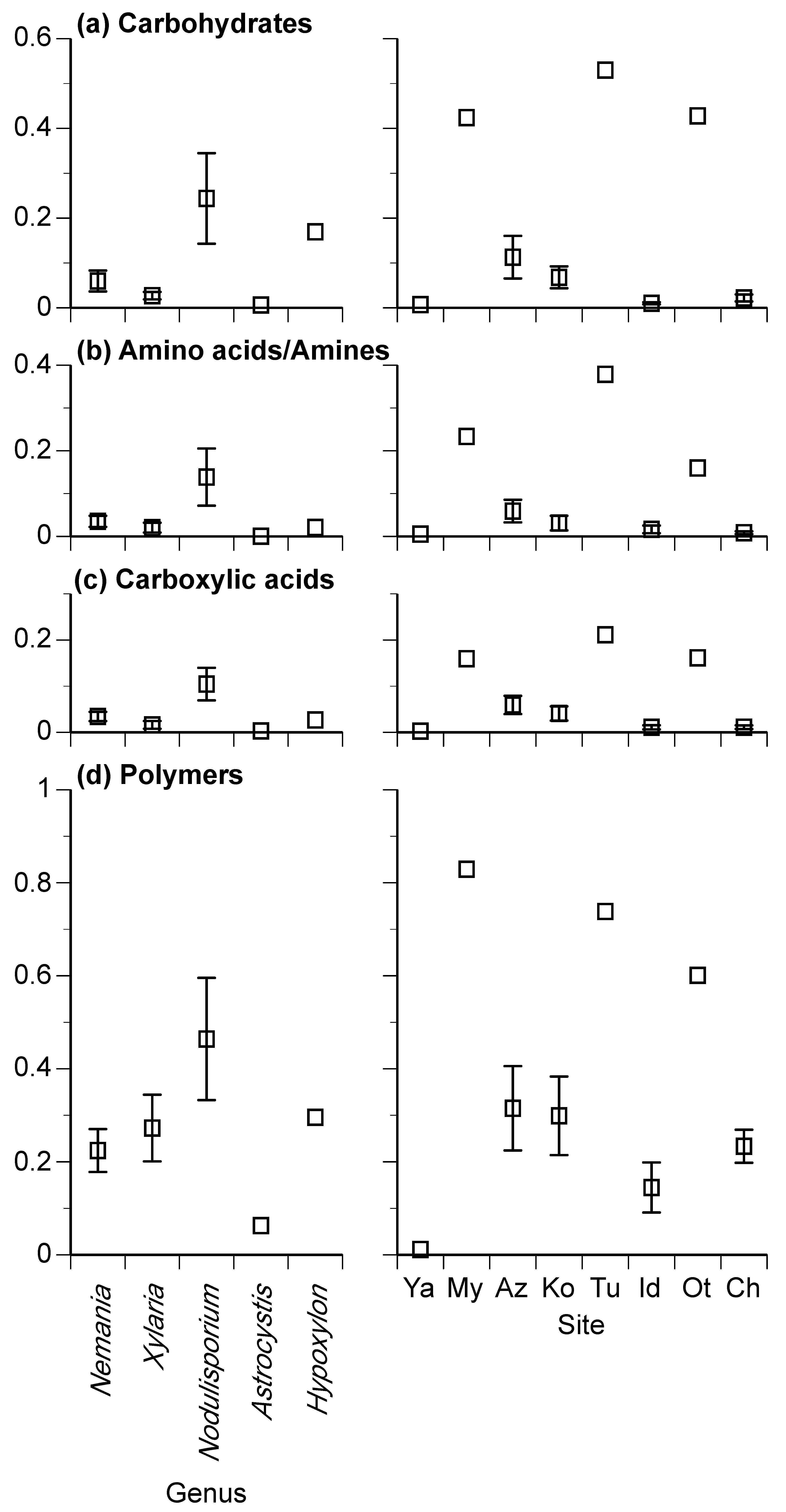
3.2. Carbon Source Utilization Patterns
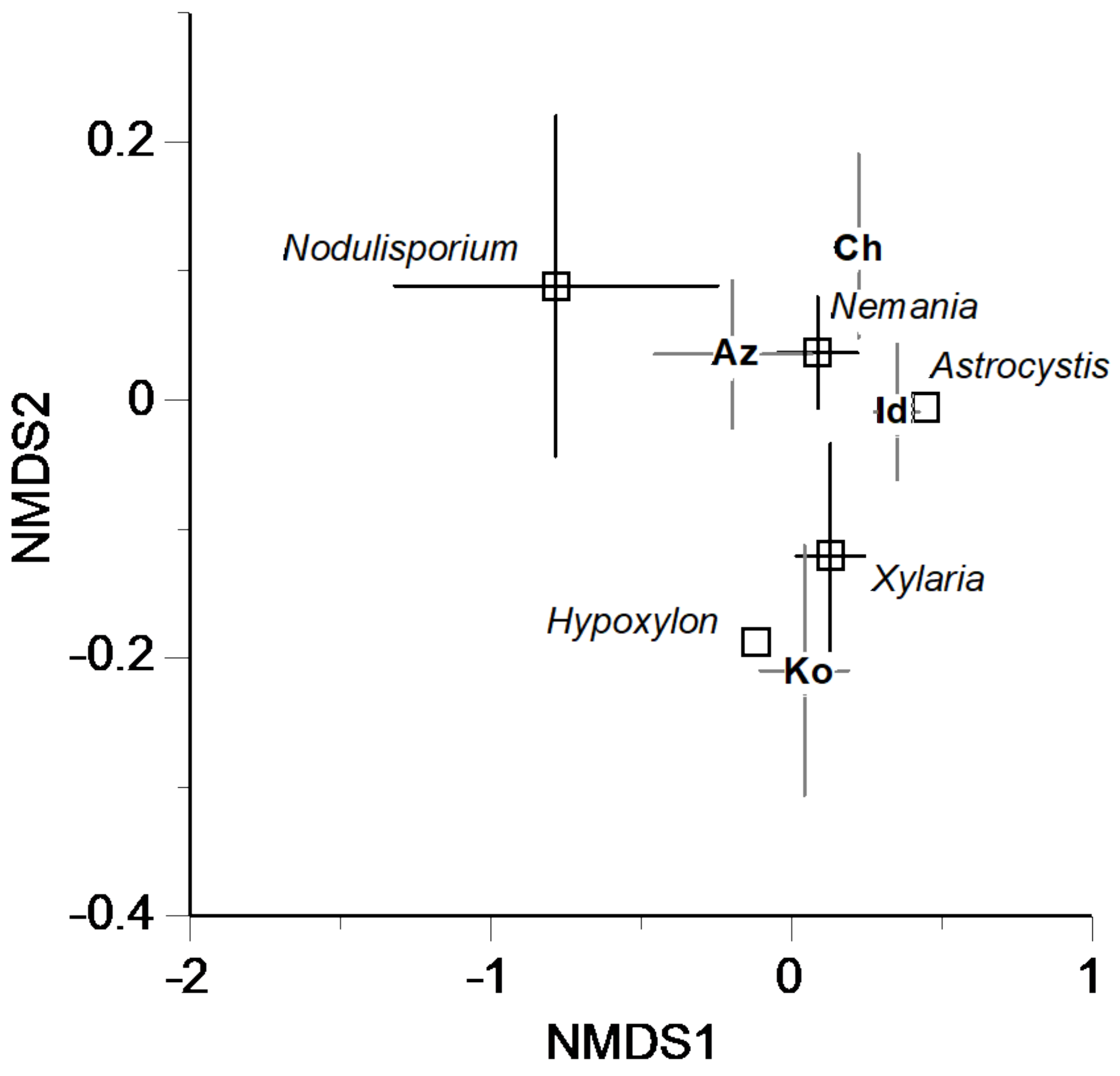
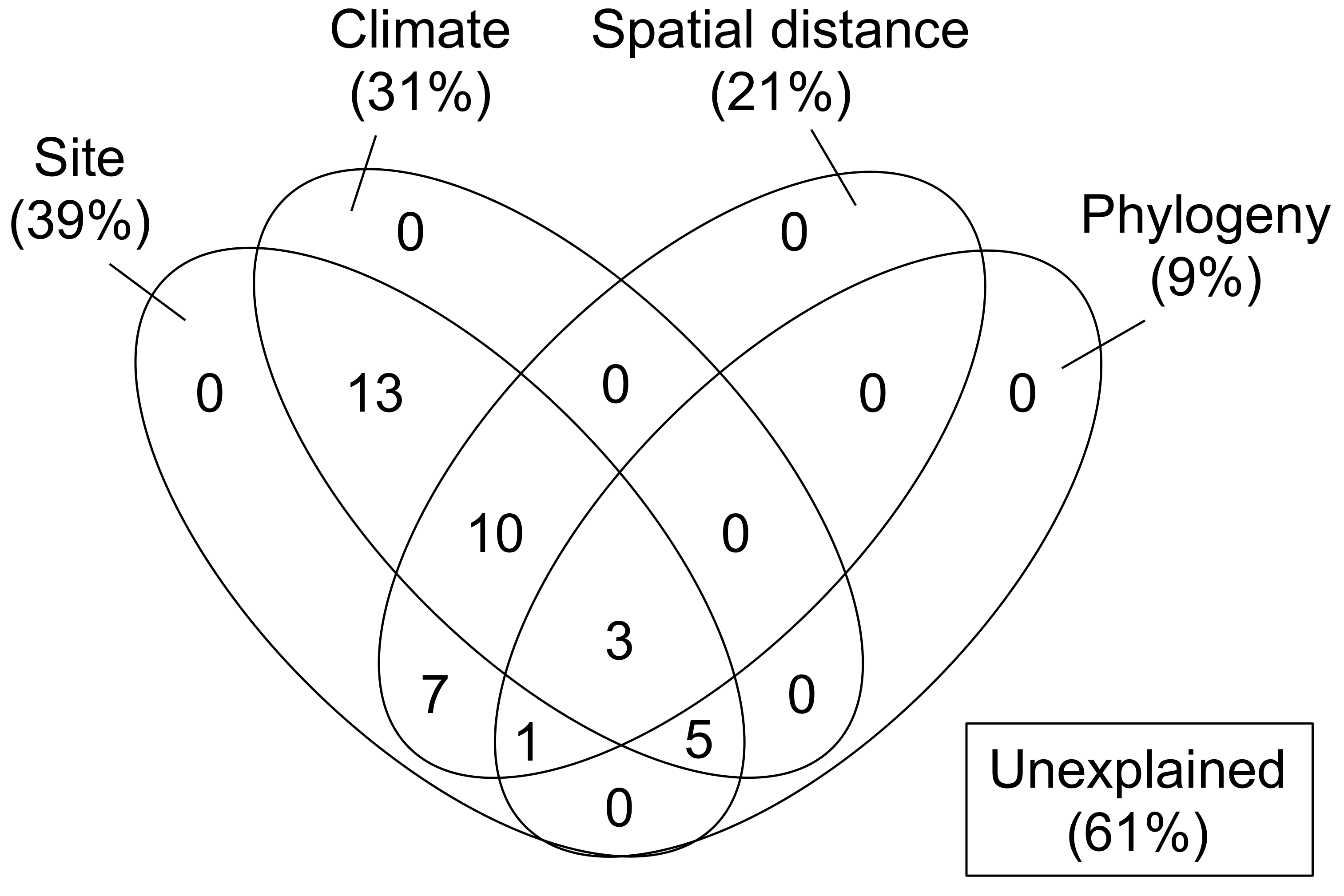
3.3. Partitioning the Dissimilarity of Carbon Source Utilization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Wal, A.; Geydan, T.D.; Kuype, T.W.; de Boer, W. A thready affair: Linking fungal diversity and community dynamics to terrestrial decomposition processes. FEMS Microbiol. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E. An enzymic approach to the analysis of microbial activity during plant litter decomposition. Agric. Ecosyst. Environ. 1991, 34, 43–54. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L.; Linkins, A.E. The enzymic basis of plant litter decomposition: Emergence of an ecological process. Appl. Soil Ecol. 1994, 1, 97–111. [Google Scholar] [CrossRef]
- Osono, T. Functional diversity of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2020, 35, 30–43. [Google Scholar] [CrossRef]
- Whalley, A.J.S. The xylariaceous way of life. Mycol. Res. 1996, 100, 897–922. [Google Scholar] [CrossRef]
- Rogers, J.D. Thoughts and musings on tropical Xylariaceae. Mycol. Res. 2000, 104, 1412–1420. [Google Scholar] [CrossRef]
- Osono, T.; To-Anun, C.; Hagiwara, Y.; Hirose, D. Decomposition of wood, petiole, and leaf litter by Xylaria species from northern Thailand. Fungal Ecol. 2011, 4, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.C.; Vandegrift, A.; Ludden, A.; Carroll, G.C.; Roy, B.A. Spatial ecology of the fungal genus Xylaria in a tropical cloud forest. Biotropica 2016, 48, 381–393. [Google Scholar] [CrossRef]
- Thomas, D.C.; Vandegrift, R.; Roy, B.A. An agent-based model of the foraging ascomycete hypothesis. Fungal Ecol. 2020, 47, 100963. [Google Scholar] [CrossRef]
- Nelson, A.; Vandegrift, R.; Carroll, G.C.; Roy, B.A. Double lives: Transfer of fungal endophytes from leaves to woody substrates. Peer J. 2020, 8, e9341. [Google Scholar] [CrossRef] [PubMed]
- Okane, I.; Srikitikulchai, P.; Toyama, K.; Læssøe, T.; Sivichai, S.; Hywel-Jones, N.; Nakagiri, A.; Potacharoen, W.; Suzuki, K. Study of endophytic Xylariaceae in Thailand: Diversity and taxonomy inferred from rDNA sequence analyses with saprobes forming fruit bodies in the field. Mycoscience 2008, 49, 359–372. [Google Scholar] [CrossRef]
- Osono, T.; Tateno, O.; Masuya, H. Diversity and ubiquity of xylariaceous endophytes in live and dead leaves of temperate forest trees. Mycoscience 2013, 54, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, A.; Matsuoka, S.; Masuya, H.; Mori, A.S.; Hirose, D.; Osono, T. Comparison of the diversity, composition, and host recurrence of xylariaceous endophytes in subtropical, cool temperate, and subboreal regions in Japan. Popul. Ecol. 2014, 56, 289–300. [Google Scholar] [CrossRef]
- Ko, W.H.; Kunimoto, R.K. Quick decline of macadamia trees: Association with Xylaria arbuscula. Plant Pathol. 1991, 40, 643–644. [Google Scholar] [CrossRef]
- Edwards, R.L.; Jonglaekha, N.; Kshirsagar, A.; Maitland, D.J.; Mekkamol, S.; Nugent, L.K.; Phosri, C.; Rodtong, S.; Ruchichachorn, N.; Sangvichien, E.; et al. The Xylariaceae as phytopathogens. Recent Res. Dev. Plant Sci. 2003, 1, 1–19. [Google Scholar]
- Okane, I.; Nakagiri, A. Taxonomy of an anamorphic xylariaceous fungus from a termite nest found together with Xylaria angulosa. Mycoscience 2007, 48, 240–249. [Google Scholar] [CrossRef]
- Guedegbe, H.J.; Miambi, E.; Pando, A.; Houngnandan, P.; Rouland-Lefevre, C. Molecular diversity and host specificity of termite-associated Xylaria. Mycologia 2009, 101, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, T.; Daniel, G. Chemistry and microscopy of wood decay by some higher ascomycetes. Holzforschung 1989, 43, 11–18. [Google Scholar] [CrossRef]
- Osono, T. Decomposition of organic chemical components in wood by tropical Xylaria species. J. Fungi 2020, 6, 186. [Google Scholar] [CrossRef]
- Wei, D.L.; Chang, S.Y.; Wei, Y.H.; Lin, Y.W.; Chuang, C.L.; Jong, S.C. Production of cellulolytic enzymes from the Xylaria and Hypoxylon species of Xylariaceae. World J. Microbiol. Biotechnol. 1992, 8, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S. Qualitative evaluation of ligninolytic enzymes in xylariaceous fungi. J. Microbiol. Biotechnol. 2000, 10, 462–469. [Google Scholar]
- Pointing, S.B.; Parungao, M.M.; Hyde, K.D. Production of wood-decay enzymes, mass loss and lignin solubilization in woody by tropical Xylariaceae. Mycol. Res. 2003, 107, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef]
- Kamilar, J.M.; Cooper, N. Phylogenetic signal in primate behaviour, ecology and life history. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120341. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, S.; Iwasaki, T.; Sugiyama, Y.; Kawaguchi, E.; Doi, H.; Osono, T. Biogeographic patterns of ectomycorrhizal fungal communities associated with Castanopsis sieboldii across the Japanese archipelago. Front. Microbiol. 2019, 10, 2656. [Google Scholar] [CrossRef] [Green Version]
- Osono, T.; Matsuoka, S.; Hirose, D. Diversity and geographic distribution of ligninolytic fungi associated with Castanopsis sieboldii leaf litter in Japan. Front. Microbiol. 2020, 11, 595427. [Google Scholar] [CrossRef]
- Osono, T.; Bhatta, B.K.; Takeda, H. Phyllosphere fungi on living and decomposing leaves of giant dogwood. Mycoscience 2004, 45, 35–41. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, A.S.; Toju, H. Two new computational methods for universal DNA barcoding: A benchmark using barcode sequences of bacteria, archaea, animals, fungi, and land plants. PLoS ONE 2013, 8, e76910. [Google Scholar]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Poyraz, N.; Mutlu, M.B. Assessment of changes in microbial communities in different operational units from a wastewater treatment plant. Pol. J. Environ. Stud. 2017, 26, 1615–1625. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Package ‘Vegan’ Version 2.6-2. Available online: http://cran.r-project.org/web/packages/vegan/index.html (accessed on 13 May 2022).
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Matsuoka, S.; Kawaguchi, E.; Osono, T. Temporal distance decay of similarity of ectomycorrhizal fungal community composition in a subtropical evergreen forest in Japan. FEMS Microbiol. Ecol. 2016, 92, fiw061. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Osono, T. Diversity and functioning of fungi associated with leaf litter decomposition in Asian forests of different climatic regions. Fungal Ecol. 2011, 4, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Osono, T.; Matsuoka, S.; Hirose, D. Variability of decomposing ability among fungi associated with the bleaching of subtropical leaf litter. Mycologia 2021, 113, 703–714. [Google Scholar] [CrossRef]
- Tornberg, K.; Olsson, S. Detection of hydroxyl radicals produced by wood-decomposing fungi. FEMS Microb. Ecol. 2002, 40, 13–20. [Google Scholar] [CrossRef]
- Ernakovich, J.G.; Wallenstein, M.D. Permafrost microbial community traits and functional diversity indicate low activity at in situ thaw temperatures. Soil Biol. Biochem. 2015, 87, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Luláková, P.; Perez-Mon, C.; Šantrůčková, H.; Ruethi, J.; Frey, B. High-alpine permafrost and active-layer soil microbiomes differ in their response to elevated temperatures. Front. Microbiol. 2019, 10, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Koda, K.; Kawaguchi, A.; Uraki, Y. Ligninolytic activity at 0 °C of fungi on oak leaves under snow cover in a mixed forest in Japan. Microb. Ecol. 2017, 74, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Legendre, P. Environmental control and spatial structure in ecological communities: An example using oribatid mites (Acari, Oribatei). Environ. Ecol. Stat. 1994, 1, 37–61. [Google Scholar] [CrossRef]
- Peay, K.G.; Garbelotto, M.; Bruns, T.D. Evidence of dispersal limitation in soil microorganisms: Isolation reduces species richness on mycorrhizal tree islands. Ecology 2010, 91, 3631–3640. [Google Scholar] [CrossRef]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of biodiversity in the Earth mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef]
- Muñiz, S.; Lacarta, J.; Pata, M.P.; Jiménez, J.J.; Navarro, E. Analysis of the diversity of substrate utilisation of soil bacteria exposed to Cd and earthworm activity using generalised additive models. PLoS ONE 2014, 9, e85057. [Google Scholar] [CrossRef]
- Talbot, J.M.; Martin, F.; Kohler, A.; Henrissat, B.; Peay, K.G. Functional guild classification predicts the enzymatic role of fungi in litter and soil biogeochemistry. Soil Biol. Biochem. 2015, 88, 441–456. [Google Scholar] [CrossRef] [Green Version]
- Russel, J.; Røder, H.L.; Madsen, J.S.; Burmølle, M.; Sørensen, S.J. Antagonism correlates with metabolic similarity in diverse bacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 10684–10688. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | Site Name | Lat (°N) | Long (°E) | Elev (m) | MAT (°C) | MAP (mm) | MSD (cm) | Sampling Month and Year |
|---|---|---|---|---|---|---|---|---|
| Ya | Yakushima Is., Kagoshima | 30.257 | 130.585 | 195 | 19.1 | 3589.8 | 0 | Aug 09 |
| My | Aya, Miyazaki | 32.027 | 131.194 | 241 | 15.4 | 2838.8 | 0 | Sept 09 |
| Az | Cape Ashizuri, Kochi | 32.740 | 132.999 | 271 | 16.9 | 2458.8 | 0 | Jul 10 |
| Ko | Kii-Oshima, Kyoto Univ., Wakayama | 33.466 | 135.833 | 94 | 17.0 | 2547.1 | 0 | Aug 11 |
| Tu | Mt. Taira, Tsushima Is., Nagasaki | 34.142 | 129.218 | 347 | 14.0 | 2122.7 | 0 | Sept 09 |
| Id | Jugei, Univ. Tokyo, Izu Pen., Shizuoka | 34.691 | 138.839 | 135 | 14.9 | 2042.9 | 2 | Aug 11 |
| Ot | Mt. Nagara, Otsu, Shiga | 35.004 | 135.857 | 130 | 14.3 | 1496.2 | 0 | Jul 10 |
| Ch | Kiyosumi, Univ. Tokyo, Chiba | 35.176 | 140.120 | 195 | 13.5 | 2075.7 | 5 | Aug 11 |
| Genus | Ya | My | Az | Ko | Tu | Id | Ot | Ch | Total |
|---|---|---|---|---|---|---|---|---|---|
| Nemania | 1 | 1 | 6 | 2 | 0 | 10 | 0 | 6 | 26 |
| Xylaria | 0 | 0 | 1 | 3 | 0 | 3 | 0 | 3 | 10 |
| Nodulisporium | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 5 |
| Astrocystis | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Hypoxylon | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Total | 1 | 1 | 9 | 7 | 1 | 14 | 1 | 9 | 43 |
| Responsible Variable | Genus | Site | Genus × Site | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Deviance | p | Deviance | p | Deviance | p | ||||
| Indices | |||||||||
| AWCD | 0.09 | 0.000 | *** | 0.22 | 0.000 | *** | 0.01 | 0.855 | ns |
| Richness of substrates utilization | 90.4 | 0.372 | ns | 333.2 | 0.028 | * | 335.6 | 0.007 | ** |
| Simpson’s D | 177.7 | 0.001 | *** | 347.2 | 0.000 | *** | 74.2 | 0.155 | ns |
| Equitability | 0.18 | 0.021 | * | 0.31 | 0.006 | ** | 0.08 | 0.420 | ns |
| Chemical groups | |||||||||
| Carbohydrates | 0.19 | 0.000 | *** | 0.40 | 0.000 | *** | 0.01 | 0.764 | ns |
| Amino acids/Amines | 0.05 | 0.000 | *** | 0.14 | 0.000 | *** | 0.02 | 0.052 | ns |
| Carboxylic acids | 0.029 | 0.000 | *** | 0.059 | 0.000 | *** | 0.002 | 0.904 | ns |
| Polymers | 0.28 | 0.181 | ns | 0.79 | 0.015 | * | 0.22 | 0.436 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabuchi, K.; Hirose, D.; Hasegawa, M.; Osono, T. Metabolic Diversity of Xylariaceous Fungi Associated with Leaf Litter Decomposition. J. Fungi 2022, 8, 701. https://doi.org/10.3390/jof8070701
Tabuchi K, Hirose D, Hasegawa M, Osono T. Metabolic Diversity of Xylariaceous Fungi Associated with Leaf Litter Decomposition. Journal of Fungi. 2022; 8(7):701. https://doi.org/10.3390/jof8070701
Chicago/Turabian StyleTabuchi, Kohei, Dai Hirose, Motohiro Hasegawa, and Takashi Osono. 2022. "Metabolic Diversity of Xylariaceous Fungi Associated with Leaf Litter Decomposition" Journal of Fungi 8, no. 7: 701. https://doi.org/10.3390/jof8070701
APA StyleTabuchi, K., Hirose, D., Hasegawa, M., & Osono, T. (2022). Metabolic Diversity of Xylariaceous Fungi Associated with Leaf Litter Decomposition. Journal of Fungi, 8(7), 701. https://doi.org/10.3390/jof8070701