

Essential Oils Reduce Grey Mould Rot of Apples and Modify the Fruit Microbiome during Postharvest Storage

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Essential Oils Preparation
2.2. In Vitro Biofumigation Test
2.3. Efficacy of EOs against Grey Mould Rots on Apples
2.4. Characterization of Volatile Compounds of EOs in the Cabinets during Storage
2.5. Apple Microbiome Sampling
2.6. Genomic DNA Extraction and Sequencing
2.7. Bioinformatics
2.8. Statistical Analysis
3. Results
3.1. EOs Efficacy Tests against Botrytis Cinerea
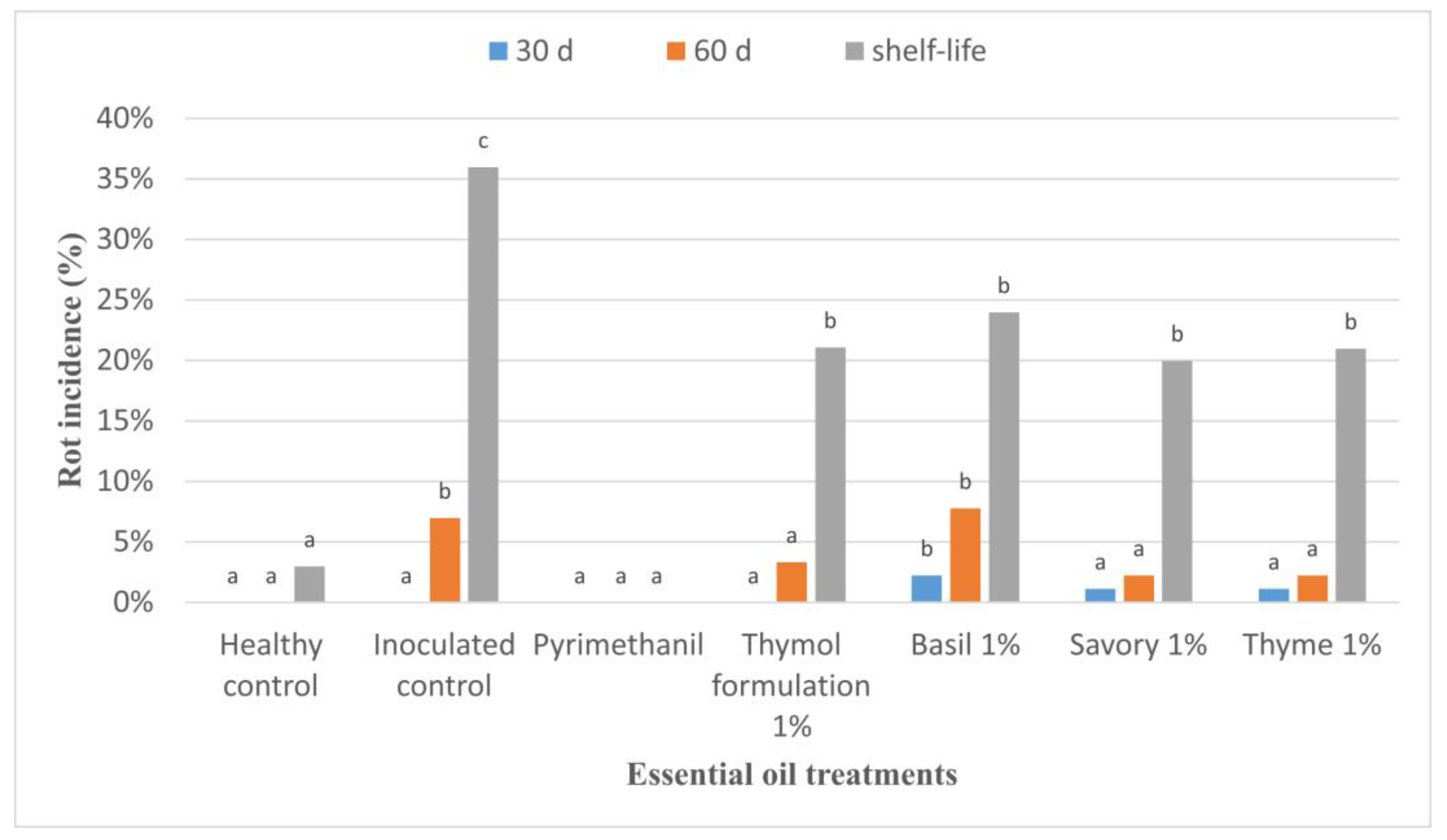
3.2. Efficacy of EOs against Grey Mould Rot on Apples
3.3. Quality Analyses on Fruits
3.4. Chemical Composition of Essential Oils and Characterization of the Storage Atmosphere
3.4.1. Characterization of Thymol Formulation
3.4.2. Characterization of Thyme Essential Oil (1%)
3.4.3. Characterization of Savoury Essential Oil (1%)
3.4.4. Characterization of Basil Essential Oil (1%)
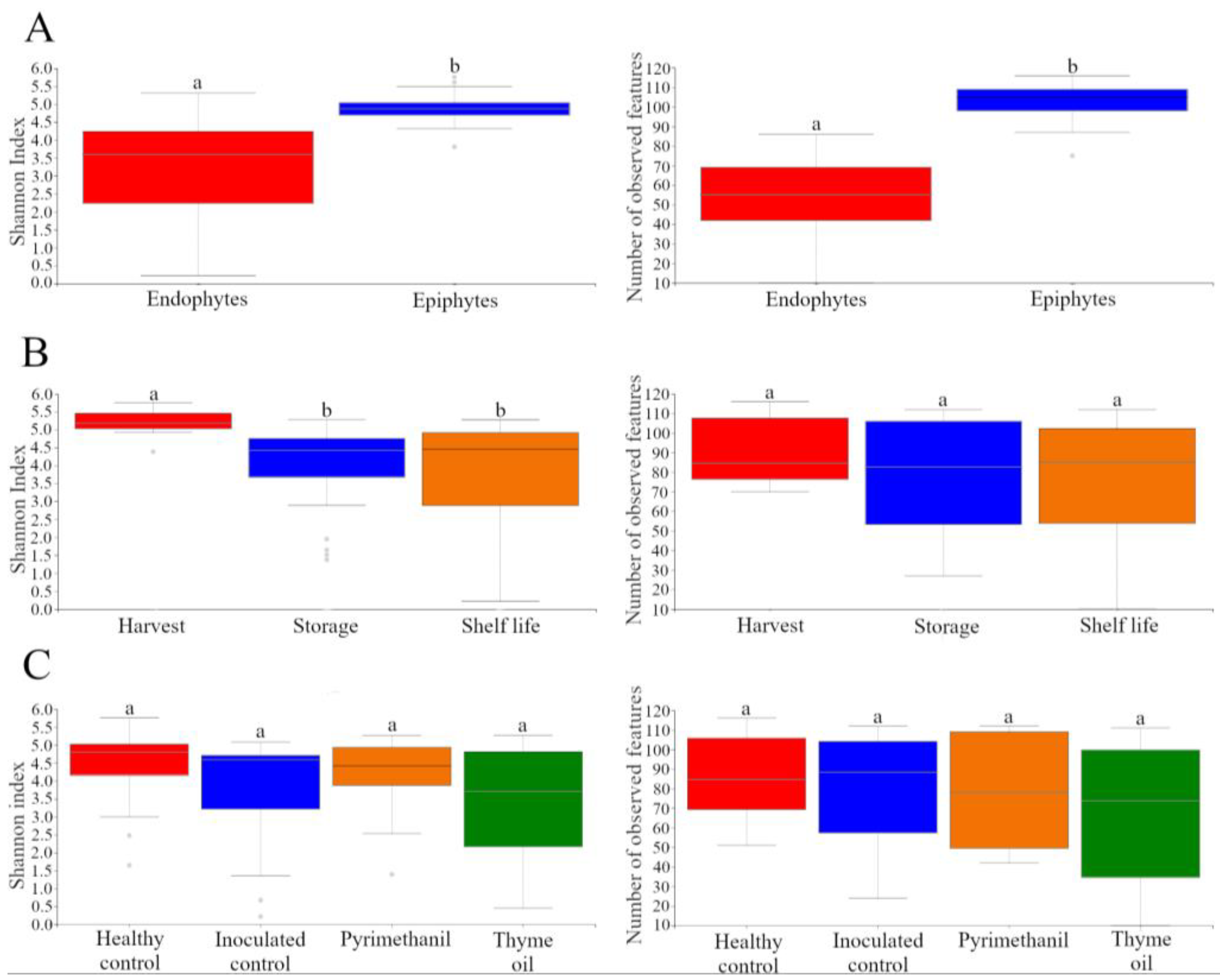
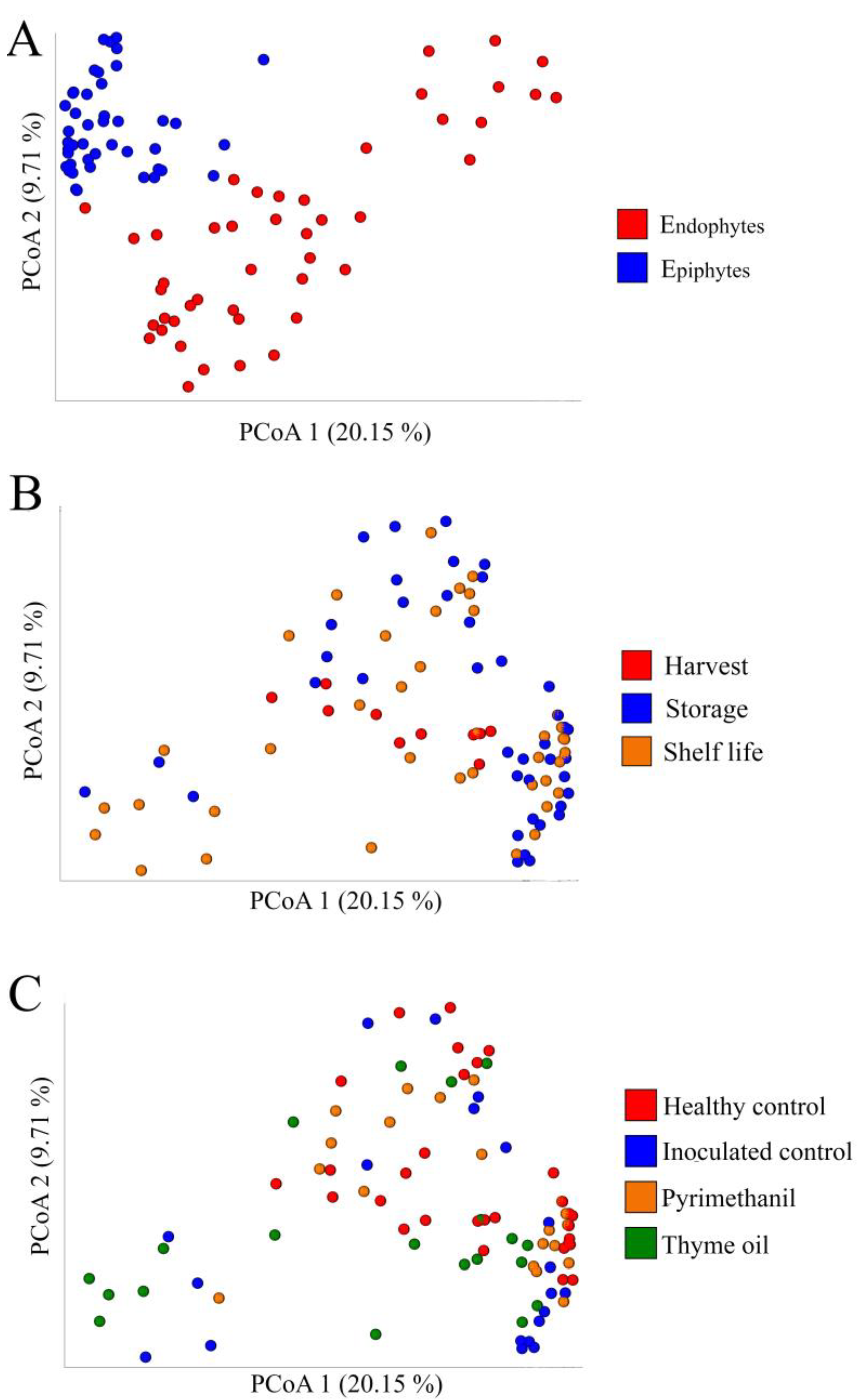
3.5. Microbial Diversity and Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Feliciano, R.P.; Antunes, C.; Ramos, A.; Serra, A.T.; Figueira, M.E.; Duarte, C.M.; de Carvalho, M.R.A.; Bronze, M.R. Characterization of traditional and exotic apple varieties from Portugal. Part 1–Nutritional, phytochemical and sensory evaluation. J. Funct. Foods 2010, 2, 35–45. [Google Scholar] [CrossRef]
- Guo, S.; Guan, L.; Cao, Y.; Li, C.; Chen, J.; Li, J.; Liu, G.; Li, S.; Wu, B. Diversity of Polyphenols in the Peel of Apple (Malus Sp.) Germplasm from Different Countries of Origin. Int. J. Food Sci. Technol. 2016, 51, 222–230. [Google Scholar] [CrossRef]
- Spadaro, D.; Torres, R.; Errampalli, D.; Everett, K.; Ramos, L.; Mari, M. Postharvest Diseases of Pome Fruit. In Postharvest Pathology of Fresh Horticultural Produce; Palou, L., Smilanick, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 55–109. [Google Scholar]
- García-Parra, J.; González-Cebrino, F.; Delgado, J.; Lozano, M.; Hernández, T.; Ramírez, R. Effect of thermal and high-pressure processing on the nutritional value and quality attributes of a nectarine purée with industrial origin during the refrigerated storage. J. Food Sci. 2011, 76, C618–C625. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, G.; Orçonneau, Y.; Moumni, M.; Davillerd, Y.; Marchand, P.A. Basic Substances, a Sustainable Tool to Complement and Eventually Replace Synthetic Pesticides in the Management of Pre and Postharvest Diseases: Reviewed Instructions for Users. Molecules 2022, 27, 3484. [Google Scholar] [CrossRef]
- Casals, C.; Teixidó, N.; Viñas, I.; Silvera, E.; Lamarca, N.; Usall, J. Combination of hot water, Bacillus subtilis CPA-8 and sodium bicarbonate treatments to control postharvest brown rot on peaches and nectarines. Eur. J. Plant Pathol. 2010, 128, 51–63. [Google Scholar] [CrossRef]
- Lahlali, R.; Serrhini, M.N.; Friel, D.; Jijakli, M.H. Predictive modelling of temperature and water activity (solutes) on the in vitro radial growth of Botrytis cinerea Pers. Int. J. Food Microbiol. 2007, 114, 1–9. [Google Scholar] [CrossRef]
- Banani, H.; Garibaldi, A.; Gullino, M.L.; Olivieri, L.; Santoro, K.; Spadaro, D. Thyme and savory essential oil efficacy and induction of resistance against Botrytis cinerea through priming of defense responses in apple. Foods 2018, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of plant essential oils on postharvest control of rots caused by fungi on different stone fruits in vivo. J. Food Prot. 2013, 76, 631–639. [Google Scholar] [CrossRef]
- Servili, A.; Feliziani, E.; Romanazzi, G. Exposure to volatiles of essential oils alone or under hypobaric treatment to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2017, 133, 36–40. [Google Scholar] [CrossRef]
- Santoro, K.; Meghenzani, M.; Chiabrando, V.; Bosio, P.; Gullino, M.L.; Spadaro, D.; Giacalone, G. Thyme and savory essential oil vapor treatments control brown rot and improve the storage quality of peaches and nectarines but could favor gray mold. Foods 2018, 7, 7. [Google Scholar] [CrossRef]
- El Ouadi, Y.; Manssouri, M.; Bouyanzer, A.; Majidi, L.; Bendaif, H.; Elmsellem, H.; Shariati, M.A.; Melhaoui, A.; Hammouti, B. Essential oil composition and antifungal activity of Melissa officinalis originating from north-Est Morocco, against postharvest phytopathogenic fungi in apples. Microb. Pathog. 2017, 107, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.M.F.D.; Steffens, C.A.; Argenta, L.C.; Amarante, C.V.T.D.; Oster, A.H.; Casa, R.T.; Amarante, A.G.M.; Espíndola, B.P. Essential oils for the postharvest control of blue mold and quality of ’Fuji’ apples. Pesqui. Agropecu. Bras. 2018, 53, 547–556. [Google Scholar] [CrossRef]
- Xylia, P.; Chrysargyris, A.; Ahmed, Z.F.R.; Tzortzakis, N. Application of Rosemary and Eucalyptus Essential Oils and Their Main Component on the Preservation of Apple and Pear Fruits. Horticulturae 2021, 7, 479. [Google Scholar] [CrossRef]
- Mbili, N.C.; Opara, U.L.; Lennox, C.L.; Vries, F.A. Citrus and lemongrass essential oils inhibit Botrytis cinerea on ‘Golden Delicious’, ‘Pink Lady’ and ‘Granny Smith’ apples. J. Plant Dis. Prot. 2017, 124, 499–511. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. Application of plant extracts to control postharvest gray mold and susceptibility of apple fruits to B. cinerea from different plant hosts. Foods 2020, 9, 1430. [Google Scholar] [CrossRef]
- Lee, S.O.; Choi, G.J.; Jang, K.S.; Lim, H.K.; Cho, K.Y.; Kim, J.-C. Antifungal activity of five plant essential oils as fumigant against post-harvest and soilborne plant pathogenic fungi. Plant Pathol. J. 2007, 23, 97–102. [Google Scholar] [CrossRef]
- Oli, N.; Singh, U.K.; Jha, S.K. Antifungal Activity of Plant’s Essential Oils against Post Harvest Fungal Disease of Apple Fruit. For. J. Inst. For. Nepal 2019, 16, 86–100. [Google Scholar] [CrossRef]
- Serag, M.S.; Baka, Z.A.; Farg, A.I.; Mohesien, M.T. BIO-CONTROL AND ULTRASTRUCTURE OF POST-HARVEST PATHOGENIC FUNGI OF APPLE FRUITS: Post-harvest pathogenic fungi of fruits. J. Microbiol. Biotechnol. Food Sci. 2022, 12, e4189. [Google Scholar] [CrossRef]
- Soković, M.D.; Vukojević, J.; Marin, P.D.; Brkić, D.D.; Vajs, V.; Van Griensven, L.J. Chemical composition of essential oils of thymus and mentha species and their antifungal activities. Molecules 2009, 14, 238–249. [Google Scholar] [CrossRef]
- Sellamuthu, P.S.; Mafune, M.; Sivakumar, D.; Soundy, P. Thyme oil vapour and modified atmosphere packaging reduce anthracnose incidence and maintain fruit quality in avocado. J. Sci. Food Agric. 2013, 93, 3024–3031. [Google Scholar] [CrossRef]
- Spadaro, D.; Gullino, M.L. Use of essential oils to control postharvest rots on pome and stone fruit. In Post-Harvest Pathology; Springer: Berlin, Germany, 2014; pp. 101–110. [Google Scholar]
- Kusstatscher, P.; Cernava, T.; Abdelfattah, A.; Gokul, J.; Korsten, L.; Berg, G. Microbiome approaches provide the key to biologically control postharvest pathogens and storability of fruits and vegetables. FEMS Microbiol. Ecol. 2020, 96, fiaa119. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Zhimo, V.Y.; Wisniewski, M.; Freilich, S. The pathobiome concept applied to postharvest pathology and its implication on biocontrol strategies. Postharvest Biol. Technol. 2022, 189, 111911. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M. The fruit microbiome: A new frontier for postharvest biocontrol and postharvest biology. Postharvest Biol. Technol. 2018, 140, 107–112. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, W.; Zhou, B.; Du, H.; Xi, L.; Zou, M.; Zou, H.; Xin, L.; Gao, Z.; Chen, Y. Variable characteristics of microbial communities on the surface of sweet cherries under different storage conditions. Postharvest Biol. Technol. 2021, 173, 111408. [Google Scholar] [CrossRef]
- Garello, M.; Piombo, E.; Prencipe, S.; Schiavon, G.; Berra, L.; Wisniewski, M.; Droby, S.; Spadaro, D. Fruit microbiome: A powerful tool to study the epidemiology of dry lenticel rot and white haze-emerging postharvest diseases of apple. Postharvest Biol. Technol. 2023, 196, 112163. [Google Scholar] [CrossRef]
- Rodriguez, A.; Batlle, R.; Nerin, C. The use of natural essential oils as antimicrobial solutions in paper packaging. Prog. Org. Coat. 2007, 60, 33–38. [Google Scholar] [CrossRef]
- Naqib, A.; Poggi, S.; Wang, W.; Hyde, M.; Kunstman, K.; Green, S.J. Making and sequencing heavily multiplexed, high-throughput 16S ribosomal RNA gene amplicon libraries using a flexible, two-stage PCR protocol. In Gene Expression Analysis; Humana Press: New York, NY, USA, 2018; pp. 149–169. [Google Scholar]
- Abdelfattah, A.; Freilich, S.; Bartuv, R.; Zhimo, V.Y.; Kumar, A.; Biasi, A.; Salim, S.; Feygenberg, O.; Burchard, E.; Dardick, C.; et al. Global analysis of the apple fruit microbiome: Are all apples the same? Environ. Microbiol. 2021, 23, 6038–6055. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME Release for Eukaryotes 2. Version 10.05.2021. Available online: https://doi.org/10.15156/BIO/1264861 (accessed on 20 October 2022).
- Nucleotide Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/nucleotide/ (accessed on 28 October 2022).
- Beule, L.; Karlovsky, P. Improved normalization of species count data in ecology by scaling with ranked subsampling (SRS): Application to microbial communities. Peer J. 2020, 8, e9593. [Google Scholar] [CrossRef] [PubMed]
- Halko, N.; Martinsson, P.; Shkolnisky, J.; Tygert, M. An algorithm for the principal component analysis of large data sets. SIAM J. Sci. Comput. 2011, 33, 2580–2594. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Guillaume Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R package version 2.5–3, 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 17 October 2022).
- Marandi, R.J.; Hassani, A.; Ghosta, Y.; Abdollahi, A.; Pirzad, A.; Sefidkon, F. Control of Penicillium expansum and Botrytis cinerea on pear with Thymus kotschyanus, Ocimum basilicum and Rosmarinus officinalis essential oils. J. Med. Plant Res. 2011, 5, 626–634. [Google Scholar]
- Abd-El-Latif, F.M. Postharvest Application of Some Essential Oils for Controlling Gray and Blue Moulds of Apple Fruits. Plant Pathol. J. 2016, 15, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, M.I.; Al-Qurashi, A.D.; Marei Almasaudi, N.; Abo-Elyousr, K.A.M. Efficacy of essential oils against gray mold and effect on fruit quality during cold storage in table grapes. S. Afr. J. Bot. 2022, 146, 481–490. [Google Scholar] [CrossRef]
- Arras, G.; Usai, M. Fungitoxic activity of 12 essential oils against four post-harvest citrus pathogens: Chemical analysis of Thymus capitatus essential oil and its effect in subatmospheric pressure conditions. J. Food Protect. 2001, 64, 1025–1029. [Google Scholar] [CrossRef]
- Russo, M.; Suraci, F.; Postorino, S.; Serra, D.; Roccotelli, A.; Agosteo, G.E. Essential oil chemical composition and antifungal effects on Sclerotium cepivorum of Thymus capitatus wild populations from Calabria, southern Italy. Rev. Bras. Farmacogn. 2013, 23, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Liu, W.T.; Chu, C.L.; Zhou, T. Thymol and acetic acid vapors reduce postharvest brown rot of apricots and plums. HortScience 2002, 37, 151–156. [Google Scholar] [CrossRef]
- Lopez-Reyes, J.G.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Efficacy of plant essential oils on postharvest control of rot caused by fungi on four cultivars of apples in vivo. Flavour. Fragr. J. 2010, 25, 171–177. [Google Scholar] [CrossRef]
- Mahmoud, G.A.; Abbas, M.S.; Soliman, A.S.; Selim, A. Effect of Essential Oils Treatments on Quality Characteristics of Anna Apple (Malus domestica Borkh) Fruits during Cold Storage. American-Eurasian, J. Agric. Environ. Sci. 2019, 19, 448–459. [Google Scholar] [CrossRef]
- Bhaskara Reddy, M.V.; Angers, P.; Gosselin, A.; Arul, J. Characterization and use of essential oil from Thymus vulgaris against Botrytis cinerea and Rhizopus stolonifer in strawberry fruits. Phytochemistry 1998, 47, 1515–1520. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Guillén, F.; Valverde, J.M.; Bailén, G.; Zapata, P.; Serrano, M.; Castillo, S.; Valero, D. Influence of carvacrol on survival of Botrytis cinerea inoculated in table grapes. Int. J. Food Microbiol. 2007, 115, 144–148. [Google Scholar] [CrossRef]
- Sakkas, H.; Papadopoulou, C. Antimicrobial activity of basil, oregano and thyme essential oils. J. Microbiol. Biotechnol. 2017, 27, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Bagamboula, C.F.; Uyttendaele, M.; Debevere, J. Inhibitory effect of thyme and basil essential oils, carvacrol, thymol, estragol, linalool and p-cymene towards Shigella sonnei and S. flexneri. Food Microbiol. 2004, 21, 33–42. [Google Scholar] [CrossRef]
- Mihajilov-Krstev, T.; Radnović, D.; Kitić, D.; Jovanović, V.S.; Mitić, V.; Stojanović-Radić, Z.; Zlatković, B. Chemical composition, antimicrobial, antioxidative and anticholinesterase activity of Satureja montana L. ssp montana essential oil. Open Life Sci. 2014, 9, 668–677. [Google Scholar] [CrossRef]
- Wan, J.; Willcock, A.; Coventry, M.J. The effect of essential oils of basil on the growth of Aeromonas hydrophila and Pseudomonas fluorescens. J. Appl. Microbiol. 1998, 84, 152–158. [Google Scholar] [CrossRef]
- Nguyen, M.P.; Lehosmaa, K.; Martz, F.; Koskimäki, J.J.; Pirttilä, A.M.; Häggman, H. Host species shape the community structure of culturable endophytes in fruits of wild berry species (Vaccinium myrtillus L., Empetrum nigrum L. and Vaccinium vitis-idaea L.). FEMS Microbiol. Ecol. 2021, 97, fiab097. [Google Scholar] [CrossRef] [PubMed]
- Ho, Q.T.; Verboven, P.; Verlinden, B.E.; Lammertyn, J.; Vandewalle, S.; Nicolaï, B.M. A Continuum model for metabolic gas exchange in pear fruit. PLoS Comput. Biol. 2008, 4, e1000023. [Google Scholar] [CrossRef]
- Hecke, K.; Herbinger, K.; Veberic, R.; Trobec, M.; Toplak, H.; Stampar, F.; Keppel, H.; Grill, D. Sugar-, acid- and phenol contents in apple cultivars from organic and integrated fruit cultivation. Eur. J. Clin. Nutr. 2006, 60, 1136–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Nie, J.; Dong, Y.; Kuang, L.; Zhang, J. Compositional shifts in the surface fungal communities of apple fruits during cold storage. Postharvest Biol. Technol. 2018, 144, 55–62. [Google Scholar] [CrossRef]
- Wassermann, B.; Kusstatscher, P.; Berg, G. Microbiome Response to Hot Water Treatment and Potential Synergy With Biological Control on Stored Apples. Front. Microbiol. 2019, 10, 2502. [Google Scholar] [CrossRef] [PubMed]
- Manthou, E.; Coeuret, G.; Chaillou, S.; Nychas, G.E. Evolution of fungal community associated with ready-to-eat pineapple during storage under different temperature conditions. Food Microbiol. 2021, 97, 103736. [Google Scholar] [CrossRef]
- Samaras, A.; Ntasiou, P.; Myresiotis, C.; Karaoglanidis, G. Multidrug resistance of Penicillium expansum to fungicides: Whole transcriptome analysis of MDR strains reveals overexpression of efflux transporter genes. Int. J. Food Microbiol. 2020, 335, 108896. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration | B. cinerea BOT1 | B. cinerea BOT2 |
|---|---|---|---|
| Mean Diameter (mm) ± SD | |||
| Thyme | 1% | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 0.5% | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| 0.1% | 8.8 ± 0.4 bc | 8.1 ± 0.5 b | |
| Basil | 1% | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 0.5% | 2.0 ± 1.1 a | 0.0 ± 0.0 a | |
| 0.1% | 65.5± 1.8 f | 76.3 ± 0.4 e | |
| Savoury | 1% | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 0.5% | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| 0.1% | 16.6 ± 0.2 d | 14.1 ± 0.5 c | |
| Thymol formulation | 1% | 2.4 ± 0.2 ab | 0.0 ± 0.0 a |
| 0.5% | 10.3 ± 1.1 cd | 8.5 ± 0.4 bc | |
| 0.1% | 52.6 ± 1.6 e | 57.3 ± 1.1 d | |
| Control | Not appliable | 77.0 ± 0.0 g | 77.0 ± 0.0 e |
| Time Point | Treatment | Firmness [N/cm2] ± SD * | Total Soluble Solids (TSS) [%] ± SD * | Titratable Acidity [%] ± SD * |
|---|---|---|---|---|
| At harvest | 43.91 ± 4.01 | 14.09 ± 1.19 | 0.60 ± 0.06 | |
| 30 days | Thymol formulation (1%) | 39.33 ± 2.84 ab | 15.03 ± 0.82 a | 0.42 ± 0.02 a |
| Thyme (1%) | 40.83 ± 4.52 b | 15.32 ± 0.85 a | 0.44 ± 0.04 a | |
| Savoury (1%) | 38.06 ± 3.13 ab | 15.24 ± 1.25 a | 0.45 ± 0.09 a | |
| Basil (1%) | 38.38 ± 1.98 ab | 15.30 ± 1.25 a | 0.40 ± 0.12 a | |
| Control | 34.40 ± 0.88 a | 14.54 ± 0.41 a | 0.40 ± 0.03 a | |
| 60 days | Thymol formulation (1%) | 34.30 ± 2.66 a | 15.12 ± 0.83 a | 0.36 ± 0.02 a |
| Thyme (1%) | 38.17 ± 7.66 a | 15.33 ± 0.98 a | 0.42 ± 0.04 b | |
| Savoury (1%) | 34.79 ± 5.10 a | 15.38 ± 0.85 a | 0.43 ± 0.01 c | |
| Basil (1%) | 34.08 ± 4.31 a | 15.59 ± 1.36 a | 0.35 ± 0.03 a | |
| Control | 30.58 ± 4.12 a | 15.09 ± 0.89 a | 0.36 ± 0.03 a |
| VOCs | EO Composition (%) | Days of Storage | ||
|---|---|---|---|---|
| 1 Day | 30 Days | 60 Days | ||
| ppm ± SD * | ||||
| Thymol | 18.81 | 4.02 ± 0.23 | 1.39 ± 0.05 | 0.64 ±0.03 |
| Propylene glycol | 6.87 | |||
| Other compounds | 74.32 | |||
| VOCs | EO Composition (%) | Days of Storage | ||
|---|---|---|---|---|
| 1 Day | 30 Days | 60 Days | ||
| ppm ± SD * | ||||
| Borneol | 1.77 | 0.00 ± 0.00 | 1.08 ± 0.19 | 0.20 ± 0.00 |
| Camphene | 0.72 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| β-Caryophyllene | 1.46 | 4.21 ± 0.52 | 0.31± 0.12 | 0.31 ± 0.11 |
| Carvacrol | 4.77 | 0.57 ± 0.18 | 0.27 ± 0.13 | 0.21 ± 0.01 |
| Eucalyptol | 0.86 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Limonene | 0.56 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Linalool | 6.42 | 2.06 ± 0.12 | 0.80 ± 0.01 | 0.00 ± 0.00 |
| O-Methylthymol | 0.43 | 0.00 ± 0.00 | 0.39 ± 0.10 | 0.41 ± 0.07 |
| β-Myrcene | 1.59 | 0.00 ± 0.00 | 0.50 ± 0.05 | 0.00 ± 0.00 |
| α-Pinene | 1.89 | 1.30 ± 0.01 | 1.05± 0.02 | 0.00 ± 0.00 |
| β-Pinene | 1.77 | 1.77 ± 0.01 | 1.16 ± 0.06 | 0.00 ± 0.00 |
| p-Cymene | 20.39 | 11.40 ± 1.82 | 26.24 ± 4.90 | 2.61 ± 0.18 |
| γ-Terpinene | 6.64 | 6.16 ± 1.03 | 2.59 ± 0.09 | 0.38 ± 0.06 |
| Terpinen-4-olo | 0.64 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Terpineol | 0.50 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Terpinolene | 1.47 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Thymol | 45.95 | 13.38 ± 2.20 | 5.13 ± 0.32 | 3.85 ± 0.57 |
| α-Thujene | 0.56 | 0.00 ± 0.00 | 0.00 ±0.00 | 0.00 ± 0.00 |
| Other compounds | 1.61 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| VOCs | EO Composition (%) | Days of Storage | ||
|---|---|---|---|---|
| 1 Day | 30 Days | 60 Days | ||
| ppm ± SD * | ||||
| Borneol | 0.74 | 0.00 ± 0.00 | 1.60 ± 0.03 | 0.00 ± 0.00 |
| β-Caryophyllene | 1.20 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Carvacrol | 60.33 | 8.64 ± 0.27 | 34.28 ± 3.90 | 3.34 ± 0.30 |
| p-Cymene | 17.07 | 4.69 ± 0.91 | 25.04 ± 2.76 | 3.95 ± 0.56 |
| β-Myrcene | 1.70 | 0.27 ± 0.04 | 0.90 ± 0.08 | 0.16 ± 0.01 |
| α-Pinene | 1.40 | 0.03 ± 0.00 | 0.37 ± 0.07 | 0.00 ± 0.00 |
| β-Pinene | 0.60 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| γ-Terpinene | 6.80 | 5.34 ± 0.42 | 5.61 ± 0.24 | 0.90 ± 0.07 |
| Terpinen-4-ol | 1.26 | 0.00 ± 0.00 | 2.27 ± 0.74 | 0.26 ± 0.01 |
| α-Terpineol | 0.60 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Terpinolene | 0.72 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Thujene | 0.66 | 0.00 ± 0.00 | 0.06 ± 0.00 | 0.00 ± 0.00 |
| Thymol | 5.52 | 0.31 ± 0.02 | 1.98 ± 0.13 | 0.24 ± 0.02 |
| Other compounds | 1.40 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| VOCs | EO Composition (%) | Days of Storage | ||
|---|---|---|---|---|
| 1 Day | 30 Days | 60 Days | ||
| ppm ± SD * | ||||
| p-Anisaldehyde | 2.68 | 0.88 ± 0.02 | 1.01 ± 0.17 | 0.49 ± 0.03 |
| t-α-Bergamotene | 0.47 | 6.60 ± 8.63 | 1.11 ± 0.08 | 0.49 ± 0.03 |
| α-Bisabolene | 1.15 | 0.00 ± 0.00 | 0.59 ± 0.06 | 0.00 ± 0.00 |
| β-Bisabolene | 1.23 | 0.00 ± 0.00 | 0.27 ± 0.20 | 0.00 ± 0.00 |
| Borneol | 0.54 | 0.00 ± 0.00 | 0.00 ±0.00 | 0.00 ± 0.00 |
| β-Cedrene | 0.28 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Citral | 0.16 | 0.21 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| β-Citral | 0.27 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Estragole | 57.79 | 128.60 ± 5.53 | 32.07 ± 1.98 | 31.42 ± 4.04 |
| α-Farnesene | 5.03 | 24.68 ± 0.34 | 111.44 ± 14.78 | 31.15 ± 3.82 |
| Fenchone | 5.21 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Humulene | 0.13 | 0.45 ± 0.06 | 0.17 ± 0.02 | 0.48 ± 0.09 |
| Linalool | 15.34 | 57.76 ± 4.36 | 2.68 ± 3.05 | 0.66 ± 0.05 |
| cis-Linanool oxide | 5.40 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.36 ± 0.05 |
| Menthol | 0.40 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Terpinen-4-ol | 0.66 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| α-Terpineol | 0.88 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Other compounds | 2.38 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Parameter | R2 | Pr (>F) |
|---|---|---|
| Sampling time point | 0.081 | *** |
| Treatment | 0.066 | *** |
| Tissue | 0.135 | *** |
| Botrytis inoculum | 0.029 | *** |
| Sampling time point × Treatment | 0.030 | ** |
| Sampling time point × Botrytis inoculum | 0.009 | n.s. |
| Sampling time point × Tissue | 0.040 | ** |
| Treatment × Tissue | 0.035 | *** |
| Botrytis inoculum × Tissue | 0.009 | n.s. |
| Sampling time point × Treatment × Tissue | 0.016 | n.s. |
| Sampling time point × Botrytis inoculum × Tissue | 0.008 | n.s. |
| Residuals | 0.541 | / |
| Total | 1.000 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavon, G.; Garello, M.; Prencipe, S.; Meloni, G.R.; Buonsenso, F.; Spadaro, D. Essential Oils Reduce Grey Mould Rot of Apples and Modify the Fruit Microbiome during Postharvest Storage. J. Fungi 2023, 9, 22. https://doi.org/10.3390/jof9010022
Schiavon G, Garello M, Prencipe S, Meloni GR, Buonsenso F, Spadaro D. Essential Oils Reduce Grey Mould Rot of Apples and Modify the Fruit Microbiome during Postharvest Storage. Journal of Fungi. 2023; 9(1):22. https://doi.org/10.3390/jof9010022
Chicago/Turabian StyleSchiavon, Giada, Marco Garello, Simona Prencipe, Giovanna Roberta Meloni, Fabio Buonsenso, and Davide Spadaro. 2023. "Essential Oils Reduce Grey Mould Rot of Apples and Modify the Fruit Microbiome during Postharvest Storage" Journal of Fungi 9, no. 1: 22. https://doi.org/10.3390/jof9010022
APA StyleSchiavon, G., Garello, M., Prencipe, S., Meloni, G. R., Buonsenso, F., & Spadaro, D. (2023). Essential Oils Reduce Grey Mould Rot of Apples and Modify the Fruit Microbiome during Postharvest Storage. Journal of Fungi, 9(1), 22. https://doi.org/10.3390/jof9010022